千屈菜无菌苗对重金属铜和铬的生理生化响应
2021-09-08张诗雯刘畅子母丹丹吴志刚
张诗雯 刘畅子 母丹丹 吴志刚 李 涛
(1. 中国科学院水生生物研究所, 武汉 430072; 2. 中国科学院大学, 北京 100049;3. 水生藻安生物科技(武汉)有限公司, 武汉 430075)
水体重金属污染是全球主要的环境问题之一[1]。随着工业和农业的不断发展以及城市化进程的持续推进, 工农业废水、固体废弃物和生活废水等污染物的排放量日趋增加, 导致水体重金属污染越来越严重。水体重金属污染会影响水生植物和动物的生存, 还会通过直接或间接的作用危害人类生命健康。铜是植物生长必需的微量元素, 但水体中高浓度的铜离子(Cu2+)会严重影响水生生物的生存,对植物具有高毒性[2], 影响植物的光合作用、膜系统、酶系统等。因此铜也被认为是生态系统中毒性最大的重金属之一[3]。铬是现代工业如电镀、橡胶、印染、油漆和机械制造等行业的重要金属元素。但其也是地壳中发现的毒性最大的重金属之一[4]。当其随工业废渣排入环境中后, 会严重污染土壤和水体, 危害植物的各项生理活动[4]。在水体中, 铬主要以三价铬离子(Cr3+)和六价铬离子(Cr6+)的形式存在。其中六价铬离子易溶于水, 其毒性约为三价铬离子的100倍。
千屈菜(Lythrum salicariaLinnaeus)属于桃金娘目千屈菜科千屈菜属, 多年生草本, 分布于全国各地。千屈菜能够有效去除水体中的总氮和总磷[5],具有较强的重金属抗性, 对土壤中的金属镉[6]、镍[1]、铜、铬、锌、锰和砷等[7,8]具有良好的富集能力。在含有铜和铬等多种重金属的废水中, 千屈菜长势良好, 具有较强的重金属抗性[9]。此外, 千屈菜为花卉植物, 外形优美, 花朵繁茂且花期长。因此, 千屈菜十分适用于修复污染水体, 尤其是重金属污染水体。但目前关于千屈菜的研究主要集中在其富集效果和净化效率方面, 对其生理响应机制的研究还十分缺乏。本研究以千屈菜无菌苗为实验材料, 可以尽量排除植株个体差异和水体其他生物(尤其是微生物)的影响。实验从光合色素、膜脂系统和抗氧化系统等方面研究千屈菜对铜(Cu2+)和铬(Cr6+)的耐性机理, 为其在重金属污染水体修复中的应用提供理论依据, 也为其他植物的抗重金属研究提供参考。
1 材料与方法
1.1 实验材料
本实验以千屈菜无菌苗为植物材料。千屈菜无菌苗培养方法: 选取生长健壮的千屈菜, 用自来水冲洗干净后再于流水下冲洗0.5h。随后在紫外超净台中, 将千屈菜先用无菌水反复冲洗3次, 再用0.1%的氯化汞灭菌8—10min, 最后用无菌水反复冲洗5次。将灭菌后的千屈菜切成5—10 mm长的茎段, 辨认形态学上端后, 置于MS基础培养基(青岛海博HB8469)中。其中, 培养基含6-苄氨基嘌呤(6-BA)1 mg/L和萘乙酸(NAA)0.1 mg/L, pH 5.8—6.0。待千屈菜茎段萌芽后, 将其转入新的MS培养基中(激素和酸碱度同上), 待其继续生长和生根, 最终成长为完整植株。
选择长势良好的千屈菜无菌苗, 取约10 cm左右长度的植株, 置于1/10 无菌Hoagland培养液(pH 5.8—6.0)中进行预培养。每隔2天更换1次培养液,预培养2周后进行实验。整个培养过程在全封闭培养箱中进行, 培养条件为平均光照40 μE/(m2·s), 光照周期为12h﹕12h光暗循环, 温度25℃。
1.2 实验设计
选取长势一致的千屈菜无菌苗,分别置于添加了0(对照)、5、10、20和40 mg/L Cu2+(CuSO4·5H2O)与添加了0(对照)、5、10、20和40 mg/L Cr6+(K2Cr2O7)的1/10无菌 Hoagland 培养液中。整个实验均在全封闭培养箱中完成, 培养条件与预培养时一致, 全程处理4d。待处理结束后, 在相同的时间段内取出千屈菜无菌苗, 用去离子水洗净、揩干, 随后测定各项生理生化指标, 每组设置3个重复。
1.3 实验方法
叶绿素含量的测定采用丙酮乙醇(1﹕1)浸提法[10]。取植株叶片剪碎后于暗光条件下浸提, 分别在645和663 nm处测定吸光度, 用Arnon公式计算叶绿素a、叶绿素b和总叶绿素的含量。
丙二醛含量的测定: 取植株叶片置于玻璃匀浆器中, 加入适量0.1 mol/L的磷酸盐缓冲液(pH 7.2),于冰水浴条件下研磨成10%的植物匀浆。离心后取上清液, 应用硫代巴比妥酸法原理[11], 采用南京建成生物工程研究所试剂盒(A003-1)进行测定。
可溶性糖含量的测定: 取植株叶片置于玻璃研磨器中, 加入适量蒸馏水研磨成10%的植物匀浆,沸水浴后离心并取上清, 应用蒽酮比色法原理[11],采用南京建成生物工程研究所试剂盒(A145-1-1)进行测定。
可溶性蛋白含量的测定: 取植株叶片置于玻璃匀浆器中, 加入适量生理盐水在冰水浴条件下研磨成10%的植物匀浆, 离心后取上清, 应用考马斯亮蓝G-250法原理[11], 采用南京建成生物工程研究所试剂盒(A045-2)进行测定。
抗氧化酶(SOD、POD、CAT)活性的测定: 取植株叶片置于玻璃匀浆器中, 加入适量0.1 mol/L的磷酸盐缓冲液(pH 7.2), 于冰水浴条件下研磨成10%的植物匀浆。3500 r/min, 离心10min, 取上清液进行测定。其中SOD、POD和CAT酶活分别应用黄嘌呤氧化酶-羟胺法[12]、愈创木酚法[11]和钼酸铵法[13]原理, 采用南京建成生物工程研究所试剂盒(A001-1、A084-3-1和A007-1 -1)进行测定。
1.4 统计分析
实验数据通过Microsoft Excel 2017进行整理并作图分析, 采用SPSS 23.0统计软件对所有指标与Cu2+和Cr6+浓度进行单因素方差分析和邓肯检验,P<0.05表示差异显著。图中不同字母表示差异水平为P<0.05, 相同字母表示差异水平为P>0.05。
2 结果
2.1 铜离子和铬离子对千屈菜叶绿素含量的影响
随着Cu2+浓度的升高, 叶绿素a、叶绿素b和叶绿素总量均呈“升-降-升”趋势(图 1a)。在Cu2+浓度为5 mg/L时, 叶绿素a、叶绿素b和叶绿素总量均显著升高(P<0.05), 达到最高值。随后, 叶绿素a逐渐下降, 并在Cu2+浓度为20 mg/L时相对对照组显著减少, 而在最高浓度(40 mg/L)时又有所上升。叶绿素b整体变化趋势与叶绿素a相同, 且在Cu2+浓度为40 mg/L时显著上升。叶绿素总量变化趋势与叶绿素a相同, 在Cu2+浓度为5 mg/L时达到最高含量为1.26 mg/g,后逐渐下降至0.93 mg/g。当Cu2+浓度升高至40 mg/L时, 叶绿素总量又显著回升。
随着Cr6+浓度的升高, 叶绿素a、叶绿素b和叶绿素总量也呈现“升-降-升”趋势(图 1b), 在Cr6+浓度为10 mg/L时三者含量均达到最高。在0—10 mg/L Cr6+浓度条件下, 千屈菜叶绿素a、叶绿素b和叶绿素总量不断上升, 其中叶绿素总量达到1.34 mg/g。当Cr6+浓度继续升高至20 mg/L时, 三者均有所下降, 其中叶绿素总量下降至1.18 mg/g。在此过程中, 叶绿素a和叶绿素总量变化显著。在最高Cr6+浓度(40 mg/L)条件下, 千屈菜叶绿素含量有所回升,其中叶绿素总量上升至1.30 mg/g, 但未超过10 mg/L时的叶绿素总量。
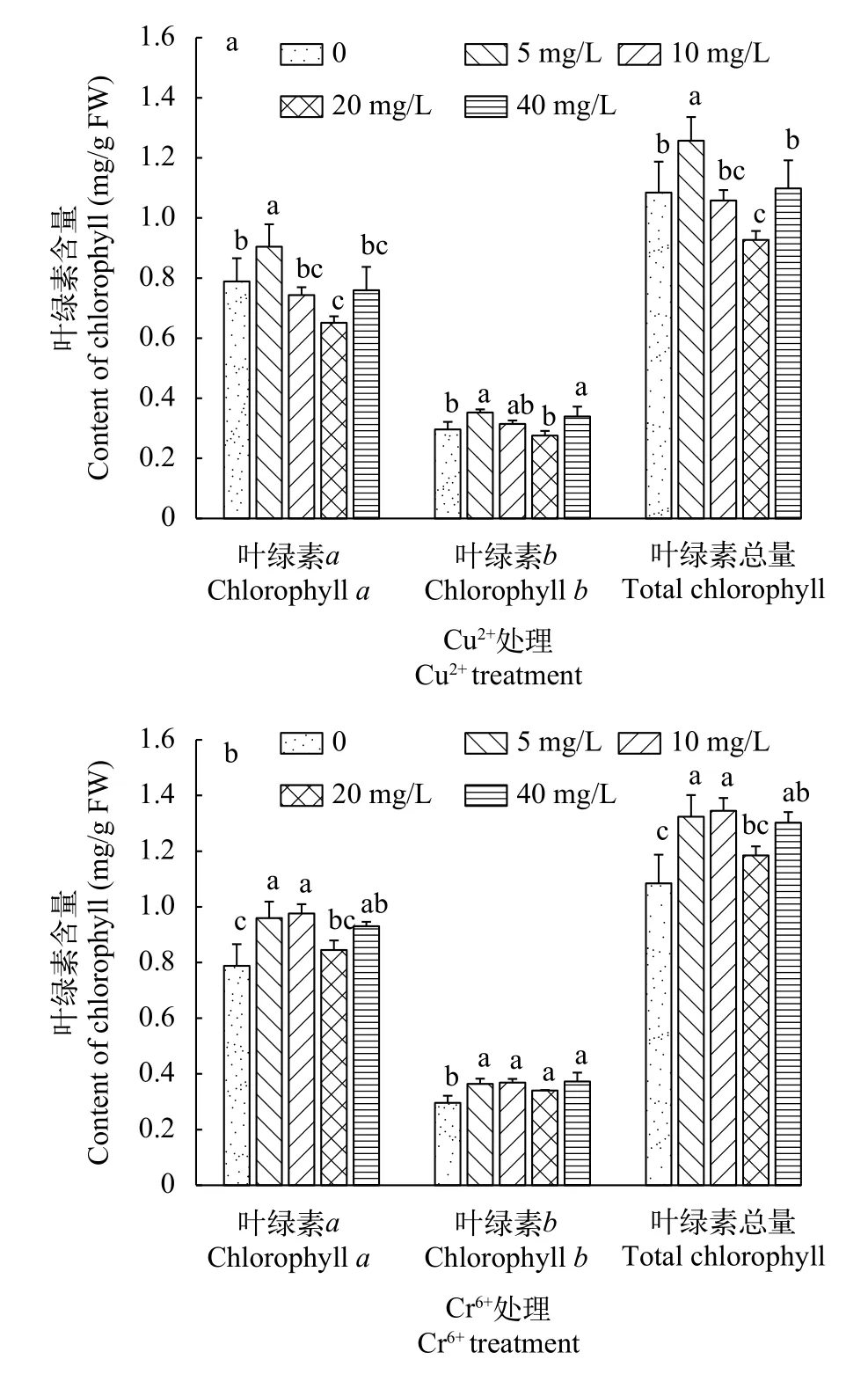
图1 不同浓度Cu2+和Cr6+对千屈菜无菌苗叶绿素含量的影响Fig. 1 The effects of different concentrations of Cu2+ and Cr6+on chlorophyll contents of Lythrum salicaria
2.2 铜离子和铬离子对千屈菜丙二醛含量的影响
在不同浓度铜离子培养条件下, 千屈菜无菌苗的丙二醛(MDA)含量均高于对照组(图 2), 整体呈上升趋势。其中Cu2+浓度为10和40 mg/L时, 千屈菜MDA含量显著高于对照组(P<0.05)。
在不同浓度铬离子培养条件下, 千屈菜无菌苗的MDA含量先降后升(图 2)。在Cr6+浓度为0—20 mg/L时, 千屈菜无菌苗MDA含量变化不显著。当Cr6+浓度增加至40 mg/L时, 千屈菜无菌苗MDA含量达到最大值0.95 nmol/mg prot, 显著高于其余所有处理组。
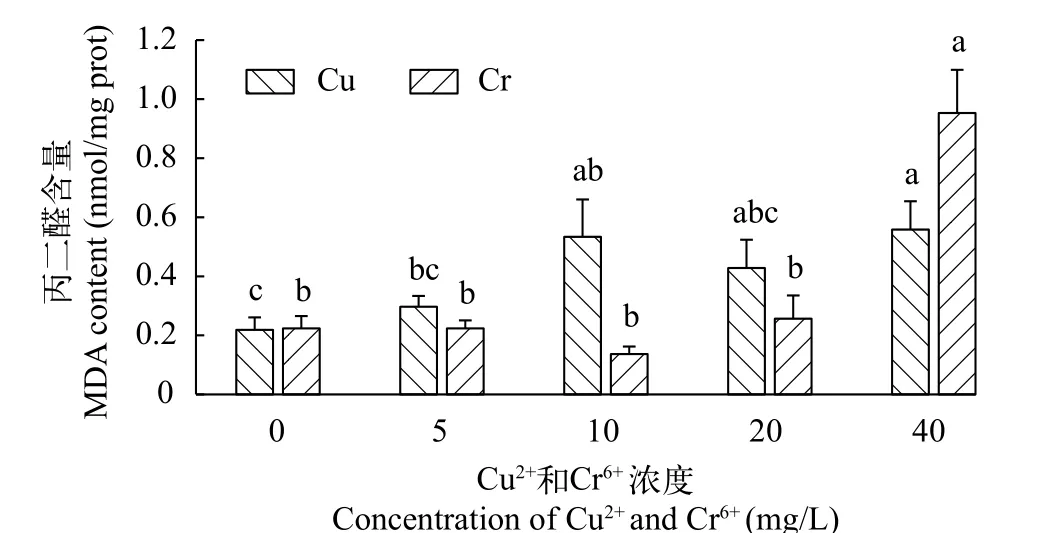
图2 不同浓度Cu2+和Cr6+对千屈菜无菌苗丙二醛含量的影响Fig. 2 The effects of different concentrations of Cu2+ and Cr6+ on malonaldehyde contents of Lythrum salicaria
2.3 铜离子和铬离子对千屈菜可溶性糖含量的影响
在不同浓度铜离子培养条件下, 千屈菜无菌苗的可溶性糖含量整体呈先升后降趋势(图 3)。在Cu2+浓度为10 mg/L时, 千屈菜无菌苗可溶性糖含量达到最高值5.02 mg/g, 显著高于对照组(P<0.05)。当Cu2+浓度从10增加至20 mg/L时, 可溶性糖含量降至2.18 mg/g, 降幅达56.61%。当Cu2+浓度继续增加至40 mg/L后, 可溶性糖含量有所上升, 但仍低于对照组。
在不同浓度铬离子培养条件下, 千屈菜无菌苗的可溶性糖含量变化趋势与铜处理下相反(图 3)。在Cr6+浓度为0—10 mg/L, 千屈菜无菌苗的可溶性糖含量逐渐下降。在Cr6+浓度为10 mg/L时可溶性糖含量降至最低为2.41 mg/g, 显著低于对照组。当Cr6+浓度增加至20—40 mg/L, 可溶性糖含量又显著上升至对照组水平。

图3 不同浓度Cu2+和Cr6+对千屈菜无菌苗可溶性糖含量的影响Fig. 3 The effects of different concentrations of Cu2+ and Cr6+ on soluble sugar contents of Lythrum salicaria
2.4 铜离子和铬离子对千屈菜可溶性蛋白含量的影响
在不同浓度铜离子培养条件下, 千屈菜无菌苗的可溶性蛋白含量整体呈先升后降趋势(图 4)。千屈菜无菌苗可溶性蛋白含量在Cu2+浓度为5 mg/L时达到最大值为4.02 g prot/L, 显著高于对照组(P<0.05)。在Cu2+浓度为10 mg/L时, 可溶性蛋白含量显著下降了39.28%。在Cu2+浓度为40 mg/L时, 可溶性蛋白浓度已降为对照组的60.86%。
在不同浓度铬离子培养条件下, 千屈菜无菌苗可溶性蛋白含量表现为“降-升-降”趋势(图 4)。当Cr6+浓度为5 mg/L时, 千屈菜无菌苗可溶性蛋白含量显著下降。随后在Cr6+浓度为10 mg/L时, 可溶性蛋白浓度上升。在Cr6+浓度为10—40 mg/L, 可溶性蛋白含量持续下降。在Cr6+浓度为40 mg/L时, 可溶性蛋白含量下降至1.10 g prot/L, 显著低于其余所有处理组。
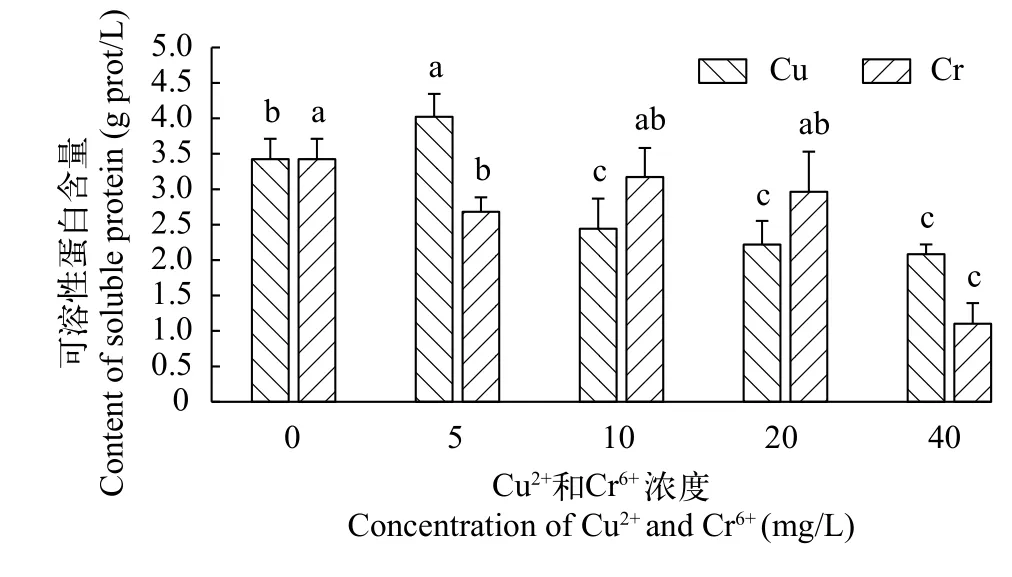
图4 不同浓度Cu2+ 和Cr6+ 对千屈菜无菌苗可溶性蛋白含量的影响Fig. 4 The effects of different concentrations of Cu2+ and Cr6+ on soluble protein contents of Lythrum salicaria
2.5 铜离子和铬离子对千屈菜抗氧化酶活性的影响
铜离子和铬离子对千屈菜SOD活性的影响 随着铜离子浓度的上升, 千屈菜无菌苗的超氧化物歧化酶(SOD)活性呈持续上升趋势(图 5)。在Cu2+浓度为5、10、20和40 mg/L条件下, SOD分别比对照组上升了8.37%、41.13%、74.17%和100.04%。在Cu2+浓度为40 mg/L时, SOD活性达到最大值为14.96 U/mg prot。
随着铬离子浓度的上升, 千屈菜无菌苗的SOD活性也呈持续上升趋势, 且均显著高于对照组(P<0.05;图 5)。在Cr6+浓度为5和20 mg/L时, 千屈菜无菌苗SOD活性分别上升41.77%和44.7%。当Cr6+浓度增加至40 mg/L时, SOD活性则上升了138.18%。
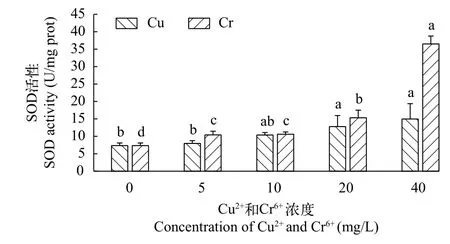
图5 不同浓度Cu2+和Cr6+对千屈菜无菌苗SOD活性的影响Fig. 5 The effects of different concentrations of Cu2+ and Cr6+ on SOD activity of Lythrum salicaria
铜离子和铬离子对千屈菜POD活性的影响 随着铜离子浓度的上升, 千屈菜无菌苗的过氧化物酶(POD)活性整体呈先升后降趋势(图 6)。当Cu2+浓度为20 mg/L时, 千屈菜无菌苗POD活性达到最大值11.76 U/mg prot, 显著高于对照组(P<0.05)。当Cu2+浓度继续增至40 mg/L时, POD活性则显著下降为6.87 U/mg prot, 低于对照组。
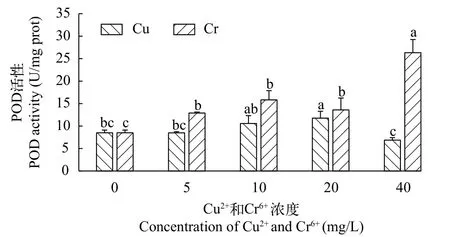
图6 不同浓度Cu2+和Cr6+对千屈菜无菌苗POD活性的影响Fig. 6 The effects of different concentrations of Cu2+ and Cr6+ on POD activity of Lythrum salicaria
千屈菜无菌苗在铬离子培养条件下的POD活性整体高于在铜离子培养条件下的(图 6)。在Cr6+浓度为0—10 mg/L, 千屈菜无菌苗的POD活性随Cr6+浓度的增加而升高。当Cr6+浓度为20 mg/L时,POD活性略有下降, 但仍高于对照组。当Cr6+浓度增加至最高40 mg/L时, POD活性显著上升至最大值26.33 U/mg prot, 显著高于其余任何处理组。
铜离子和铬离子对千屈菜CAT活性的影响 千屈菜无菌苗的过氧化氢酶(CAT)活性随着铜离子浓度的增加呈持续上升趋势(图 7), 且其整体活性远高于铬离子培养条件下的活性。CAT活性在各Cu2+浓度处理组间变化显著(P<0.05)。当Cu2+浓度在10 mg/L时, CAT活性已显著高于对照组。当Cu2+浓度为40 mg/L时, CAT活性达到最大值12.10 U/mg prot,是对照组的2.7倍。
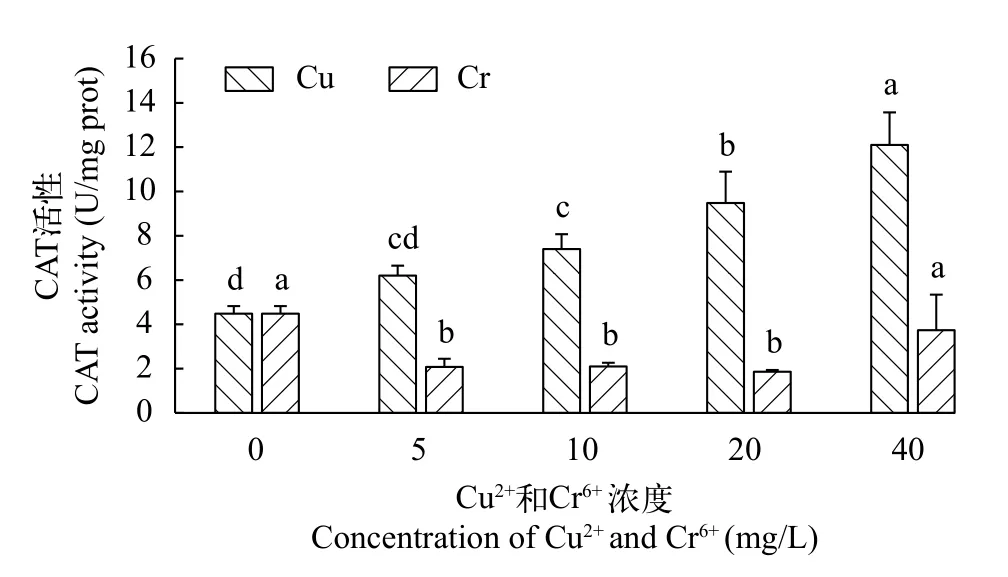
图7 不同浓度Cu2+和Cr6+对千屈菜无菌苗CAT活性的影响Fig. 7 The effects of different concentrations of Cu2+ and Cr6+ on CAT activity of Lythrum salicaria
在铬离子培养条件下, 千屈菜无菌苗CAT活性整体水平均低于对照组(图 7)。在最低Cr6+浓度5 mg/L条件下, CAT活性即显著低于对照组。在Cr6+浓度为5—20 mg/L, CAT活性变化不大。当Cr6+浓度增加至40 mg/L时, CAT活性显著上升至3.73 U/mg prot, 但仍低于对照组。
3 讨论
3.1 铜和铬离子对千屈菜叶绿素含量的影响
叶绿素广泛存在于植物的绿色组织中, 是植物进行光合作用的物质基础。叶绿素的含量与植物的光合作用和营养状况密切相关, 能够反映植物的生长状况。重金属对植物叶绿素的影响主要是破坏叶绿体膜和类囊体膜, 抑制叶绿素的合成[14]。
在本研究中, 千屈菜无菌苗在铬离子培养条件下的叶绿素含量显著高于铜离子。在2种离子培养条件下, 千屈菜无菌苗叶绿素含量均呈“升-降-升”的趋势。在低浓度铜处理中千屈菜叶绿素含量呈上升趋势, 其原因可能是Cu2+是叶绿体质体蓝素的组成成分, 还是叶绿素合成过程中酶促反应的辅因子[15]。因此, 低浓度Cu2+能促进叶绿素的合成。但随着Cu2+浓度的升高, 大量Cu2+进入植物体内, 破坏了叶绿素的结构或使得叶绿素加速分解。
在低浓度的Cr6+培养条件下, 千屈菜叶绿素含量显著上升, 这可能是千屈菜受到低浓度Cr6+刺激,促进了叶绿素的合成。但随着Cr6+浓度在千屈菜细胞内的大量积累, 取代了叶绿体所需要的铁和镁元素, Cr6+与蛋白质上的SH基结合[16,17], 损害了叶绿体的结构和功能。此外, 千屈菜在铜和铬离子培养条件下叶绿素a/b值逐渐降低, 说明叶绿素a受到的影响更大, 植株对光能的捕获能力降低。苦草在铜处理下的响应也是如此[18]。
重金属常通过直接或间接损伤叶绿体结构或与相关酶作用干扰叶绿素合成途径来降低植物体内的叶绿素浓度。植物在重金属条件下叶绿素含量常表现为逐渐降低或低促高抑现象。此外也有研究发现一些植物在高浓度重金属处理下, 其叶绿素含量会增加。例如美人蕉在高浓度(40 mg/L)铬胁迫下, 叶绿素含量有所升高[19]。再力花 (Thalia dealbata)[20]、江蓠 (Gracilaria verrucosa)[21]和线叶眼子菜 (Potamogeton pusillus)[22]等在重金属处理下, 叶绿素含量呈“升-降-升”趋势。但出现该现象的机理尚不清楚, 在本实验中, 千屈菜在40 mg/L铜和铬处理下, 叶绿素含量也有所上升, 可能与特定浓度造成的抗性机制或应答有关[20,23]。
3.2 铜和铬离子对千屈菜丙二醛含量的影响
细胞膜具有调节和控制物质进出细胞的重要生理功能。植物在胁迫条件下, 酶系统和非酶系统会产生氧自由基, 导致膜脂过氧化[2], 形成脂质过氧化物, 损害细胞膜稳定性, 影响植物的正常代谢。其中MDA是膜脂过氧化最重要的产物之一, 其含量可反映植物脂质过氧化程度和细胞损伤程度。在本实验中, 千屈菜无菌苗在不同浓度Cu2+培养条件下MDA含量整体呈上升趋势, 各Cu2+浓度处理组MDA含量均高于对照组。在Cu2+浓度为20 mg/L时膜脂化程度略有缓解, 说明千屈菜细胞保护机制起了一定作用, 但其含量依然高于对照组。这说明千屈菜细胞膜在各浓度铜处理下均受到损害。菹草无菌苗在铜处理下也表现出类似的机制[24]。
在本实验中, 千屈菜无菌苗MDA含量整体随Cr6+浓度的增加而升高。黑藻和绿萝[25]在铬胁迫下其MDA含量也随Cr6+浓度的升高而升高, 但上升幅度明显。而千屈菜在Cr6+浓度为0—20 mg/L时,MDA含量上升幅度较小, 与对照组的差异均不显著, 表明其受到的影响较小。在Cr6+浓度为10 mg/L时, 千屈菜MDA含量甚至略有下降, 说明其保护机制产生了一定的作用。但当Cr6+浓度增加至40 mg/L时, 千屈菜MDA含量骤增, 显著高于其余所有处理组。这表明此时其保护机制可能已经无法及时清除过多的氧自由基, 导致细胞膜脂化程度快速加重。综上, 千屈菜细胞膜脂化程度受铜离子损害较大, 而低浓度Cr6+(≤20 mg/L)对千屈菜细胞膜脂化程度影响很小。
3.3 铜和铬离子对千屈菜渗透调节物质的影响
可溶性糖和可溶性蛋白是植物体内重要的渗透调节物质。在重金属胁迫下, 植物可以通过积累可溶性糖和可溶性蛋白来维持细胞渗透势, 保证水分供应和生理代谢。可溶性糖在植物体内分布广泛, 可在胁迫条件下提高植物保水力和吸水力, 提高原生质的弹性[26]。在本实验中, 千屈菜无菌苗可溶性糖含量在铜处理下呈先升后降趋势, 与千屈菜在Cd和Pb胁迫下的变化一致[27], 表现为低促高抑现象。当Cu2+浓度为5和10 mg/L时, 千屈菜可溶性糖含量高于对照组, 说明其可通过提高可溶性糖含量来维持细胞渗透势。而高浓度的Cu2+对千屈菜可溶性糖有抑制作用, 但当Cu2+浓度为40 mg/L时可溶性糖含量又有所回升, 且与对照组差异不显著。在铬处理下, 千屈菜可溶性糖含量变化趋势则与Cu2+相反, 表现为先降后升趋势, 与菹草在铬胁迫下的响应一致[28]。这可能是由于低浓度Cr6+不足以引起千屈菜渗透系统的防御反应, 反而刺激了千屈菜的某些生理活动如光合产物合成, 使得可溶性糖的利用效率增加。但可溶性糖含量除在Cr6+浓度为10 mg/L时显著下降外, 其余处理组变化均不显著。这表明千屈菜可溶性糖含量在铬处理下所受影响可能不是很大, 具有较好的铬抵抗能力。
可溶性蛋白也是植物体内分布广泛的营养物质和渗透调节物质[26], 可维持细胞渗透势, 调节代谢, 提高植物抗胁迫的能力。在本研究中, 千屈菜无菌苗可溶性蛋白在铜处理下呈先升后降趋势。这与菹草(Potamogeton crispus)在铜胁迫下可溶性蛋白的变化趋势一致[24]。三叶草(Trifolium pretense)的铜污染研究也表明低浓度Cu2+可诱导可溶性蛋白的含量, 而高浓度Cu2+则有抑制作用[29]。这说明低浓度Cu2+对千屈菜可溶性蛋白含量具有激活作用。此外, 可溶性蛋白含量在10、20和40 mg/L之间下降幅度不显著, 说明千屈菜可能对Cu2+具有一定抗性。在铬处理下, 千屈菜可溶性蛋白含量则整体呈下降趋势, 这与苦草在铬处理下的响应一致[30]。但较低浓度的Cr6+对其影响不大, 在Cr6+浓度20 mg/L时, 千屈菜可溶性蛋白含量依然与对照组无显著差异。而当Cr6+浓度进一步升高为40 mg/L时, 可溶性蛋白含量才发生显著下降。综上, 千屈菜在铜和铬处理下均具有一定的渗透调节抗性, 其中铬处理对千屈菜渗透调节物质的影响相对较小。
3.4 铜和铬离子对千屈菜抗氧化酶活性的影响
在重金属胁迫下, 植物会产生大量活性氧(ROS), 这些活性氧在细胞中大量积累后会损伤细胞膜、大分子蛋白质、DNA和RNA等。为抵御活性氧带来的毒性作用, 植物会激活抗氧化酶机制来清除过多的活性氧。SOD、POD和CAT是植物体内重要的清除活性氧的酶类。SOD可以将O2-转化为毒性相对较弱的H2O2, 而CAT和POD则可以把H2O2进一步转化为H2O和O2[31]。在铜胁迫下, 轮叶黑藻 (Hydrilla verticillata) 的SOD和POD活性持续升高[32], 鱼草(Cabomba caroliniana)的POD和CAT活性也逐渐增强[33], 以缓解铜离子的胁迫作用。在铬胁迫下, 2种苋属植物的SOD、POD和CAT活性不断增强[16], 超富集植物李氏禾(Leersia hexandra)在Cr6+浓度20 mg/L以内抗氧化酶活性也逐渐升高[31],以抵御氧化胁迫。
在本实验中, 千屈菜SOD活性在铜和铬处理下均呈持续上升趋势, 且在铬处理下千屈菜的SOD活性整体显著高于铜。这表明千屈菜在铜和铬处理下能良好地激发SOD活性, 清除过多的自由基, 降低氧化损伤。并且相对于铜处理而言, 千屈菜SOD对铬处理反应更加敏感。千屈菜POD活性在铜处理下表现为先升后降, 表明低浓度Cu2+能激发POD活性, 而高浓度Cu2+会抑制POD活性。在铬处理下, 千屈菜POD活性则整体呈上升趋势, POD活性随Cr6+浓度的增加不断上升, 以清除机体内的过氧化氢, 且其POD响应机制相对于铜处理下更为积极。千屈菜在铜处理下, CAT活性随Cu2+浓度增加而不断升高, 表明CAT在铜处理下发挥了重要作用,与伊乐藻(Elodea canadensis)的响应一致[34]。而铬处理则对千屈菜CAT活性表现为在所有浓度范围内的持续抑制效应, 这与Halimione portulacoides根系对铬的响应相似[35]。综上, 千屈菜抗氧化酶系统在铜和铬处理下响应积极。在铜处理下千屈菜体内活性氧的清除由SOD、POD和CAT三种酶共同参与, 而在铬处理下参与活性氧清除的酶主要是SOD和POD。
4 结论
本实验分别测定了铜(Cu2+)和铬(Cr6+)培养条件下千屈菜无菌苗的叶绿素、丙二醛、可溶性糖、可溶性蛋白含量和SOD、POD、CAT活性。结果表明千屈菜对Cu2+和Cr6+具有一定的抗性, 在2种离子处理下千屈菜叶绿素含量基本未受影响,抗氧化酶活性上升, 且具有一定的渗透调节能力。千屈菜在Cu2+浓度低于5 mg/L时生理生化指标所受影响相对较小, 表明其具有修复水体低浓度铜污染的潜力; 而千屈菜在Cr6+浓度低于20 mg/L时其生理生化指标所受影响相对较小, 与一些铬超富集植物的耐受能力相近, 表明其在铬污染水体修复方面具有较高的应用潜力。但千屈菜无菌苗在水体修复中的实际应用还需要更深入的研究。
