围封年限对内蒙古灌丛化草原小叶锦鸡儿灌丛结构及群落种间关联的影响
2021-09-01丁新峰王金龙赵念席高玉葆
董 轲,丁新峰,郝 广,王金龙,赵念席,*,高玉葆
1 南开大学生命科学学院, 天津 300071 2 天津农学院农学与资源环境学院, 天津 300191
近半个世纪以来,气候变化及人类活动的加剧导致全球范围内的干旱半干旱草原发生了不同程度的退化,具体表现在草地生产力下降,物种多样性丧失,水土流失加剧,草原生态系统的社会服务功能降低[1- 2]。草原灌丛化作为退化草原的一种类型在全球范围内广泛发生,在我国内蒙古干旱半干旱典型草原区,主要表现为小叶锦鸡儿(Caraganamicrophylla)对典型草原的入侵,并形成了连片的灌丛化草地[3- 4]。虽然已经有大量的研究证明围封能够有效促进退化草地恢复到退化前状态[5- 6],但这些研究主要关注了未发生灌丛化的退化草地。对灌丛化草地的长期观测发现,仅通过围封很难使灌丛化草地恢复到退化前状态,如关于美国亚利桑那州东南部灌丛化草地的研究中指出,50 a的围封禁牧后其仍处于灌丛化草地状态[7];同时,有研究者推测灌丛化草地也是干旱半干旱草原群落的一种稳态[8]。因此,合理评价灌丛化草地群落状态及演替规律,对该类草原的利用和保护具有重要意义,且已经成为当前草地生态学及恢复生态学中重点关注的内容之一[9]。
群落关键种种群结构及群落内主要物种的种间关联,均能从不同角度反映群落的发展规律,并有助于对群落稳定性及演替规律进行合理的预测[10-13]。就种群结构来说,关键种或者建群种种群内小个体级占优时,为上升型种群;大个体级占优势时,为下降型种群;中个体级占优时,种群相对稳定[14],种群的结构特征对其所在群落也具有指示作用。而群落内种间关联的变化直接影响群落特征,是理解群落稳定程度及群落演替方向的重要基础[15- 16]。群落内主要物种的种间正关联通常有利于维持植物群落的生产力和稳定性、提高生态系统的服务功能;而负关联体现了物种间的排斥性,不利于生态系统的稳定;无关联常常表现为中性作用调控的种间关系,物种彼此几乎不会对邻居物种产生影响[17]。另外,通过种间关联分析有助于理解群落发育过程中种群之间的相互作用,表征优势种群的地位[18]。奇凯等对赤峰市沙地油松(Pinustabulaeformis)林草本植物种间关联的研究,有效地反映了群落内的物种发育动态以及物种组成在不同发育时期此消彼长的动态变化[17]。然而,自然恢复或者围封条件如何影响入侵灌木种群结构及灌丛间主要物种种间关联的研究并不多见,严重限制了人们对灌丛化草地群落动态的解释及合理预测,相关工作急需展开。
本文在锡林郭勒草原区选择不同围封年限的小叶锦鸡儿灌丛化草地,按不同灌丛化程度设立样区并进行调查,在对关键种小叶锦鸡儿种群结构及灌丛间主要物种种间关联调查的基础上,深入理解小叶锦鸡儿灌丛化草地在围封条件下的演替规律,为该类草原的合理利用及恢复政策的制定提供数据支持及理论指导。
1 材料与方法
1.1 研究区域概况
2018年8月,选取围封年限不同的小叶锦鸡儿灌丛化草地,分别为1979年围封样地(43°32′N,116°40′E,海拔1200 m)、1983年围封样地(43°37′N,116°40′E,海拔1186 m)和2003年围封样地(43°38′ N,116°41′ E,海拔1185 m)。三处样地均位于内蒙古锡林郭勒盟白音锡勒牧场,两两之间的距离大约5 km,具有相似的气候特征(半干旱温带大陆性气候)和土壤特征(栗钙土),地带性植被为大针茅(Stipagrandis)-羊草(Leymuschinensis)典型草原。由于不合理利用,该区域退化严重,大针茅和羊草群落被小叶锦鸡儿灌丛化草地替代现象已经非常普遍。据统计,小叶锦鸡儿已入侵了超过5×106hm2内蒙古草原[9],部分区域即使围封40 a,小叶锦鸡儿灌丛在景观上仍具有显著作用。
1.2 实验设置
受退化程度大小的影响,灌丛化程度会表现出由高至低的趋势。在选择的三处样地内,依据小叶锦鸡儿灌丛盖度划分为低盖度(小叶锦鸡儿盖度3%—5%)、中盖度(小叶锦鸡儿盖度12%—18%)和高盖度(小叶锦鸡儿盖度35%—45%)3种灌丛化水平样区。于每个灌丛化水平样区内,在灌丛间随机设置10个0.5 m×0.5 m草本样方,保证样方边界距离小叶锦鸡儿灌丛的距离大于1.0 m,调查样方内植物种类并按照多年生禾草、多年生杂类草、一二年生植物及半灌木进行功能群分类,统计每种植物的密度和最大株高,并以种为单位收获地上部带回实验室于80℃烘干至恒重,测定地上部生物量;同时,随机设置4个20 m × 20 m灌木样方来调查样方内每株小叶锦鸡儿的冠幅大小,用于种群大小结构分析。其中小叶锦鸡儿种群大小结构以样区为单位进行数据分析,灌丛间群落以样地为单位进行数据分析。
1.3 数据分析
1.3.1种群大小结构
种群的大小能够反映种群增长的动态变化[19],对于构件灌木,通常用冠幅大小来代表。在本研究中,将小叶锦鸡儿冠幅长度每隔0.5 m划分为一个等级,共划分成7个等级:[0, 0.5],(0.5, 1],(1, 1.5],(1.5, 2],(2, 2.5],(2.5, 3],(3, 3.5],分别统计不同围封年限不同灌丛盖度水平下每个等级小叶锦鸡儿的数量,并评估小叶锦鸡儿种群大小结构的偏度,确定分布模式。
1.3.2物种重要值(IV)[20]及生态位宽度(B)[21]
重要值(IV)=(相对密度+相对高度+相对生物量)/3
相对密度(高度、生物量)=某一物种在一样方内的平均密度(高度、生物量)/该样方内所有物种的密度(高度、生物量)之和
式中,Bi为物种i的生态位宽度,Pij为物种i在资源梯度级j上的数量特征,nij为物种i在资源梯度级j上的优势度(在本实验中指物种i在对应样方中的重要值),r为资源等级数,上述公式具有值域[0,logr][22]。
1.3.3群落整体关联性分析-方差比率法(VR)[23]
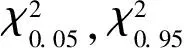
1.3.4统计分析
对小叶锦鸡儿冠幅大小数据进行对数转化使其符合正态分布及方差齐性,利用双因素方差分析(Two-way ANOVA)检验围封年限、灌丛化水平及其交互作用对小叶锦鸡儿冠幅大小的影响。如交互作用显著,则进一步通过简单效应分析来评估各个单一因子的影响。以上数据分析均在SPSS 22.0中完成。
2 结果与分析
2.1 小叶锦鸡儿种群大小结构
对小叶锦鸡儿冠幅大小结构分析发现,仅1983年围封低盖度样区内,小叶锦鸡儿冠幅分布格局为正态分布;其他8个样区内分布格局偏度系数均大于0,表现为显著正偏。在所有样区内,均为0.5—2 m冠幅区间的小叶锦鸡儿灌丛占优势,0—0.5 m冠幅区间内的小叶锦鸡儿比例低,且在1979年围封低盖度样区内,未统计到0—0.5 m冠幅区间内的小叶锦鸡儿(表1,图1)。
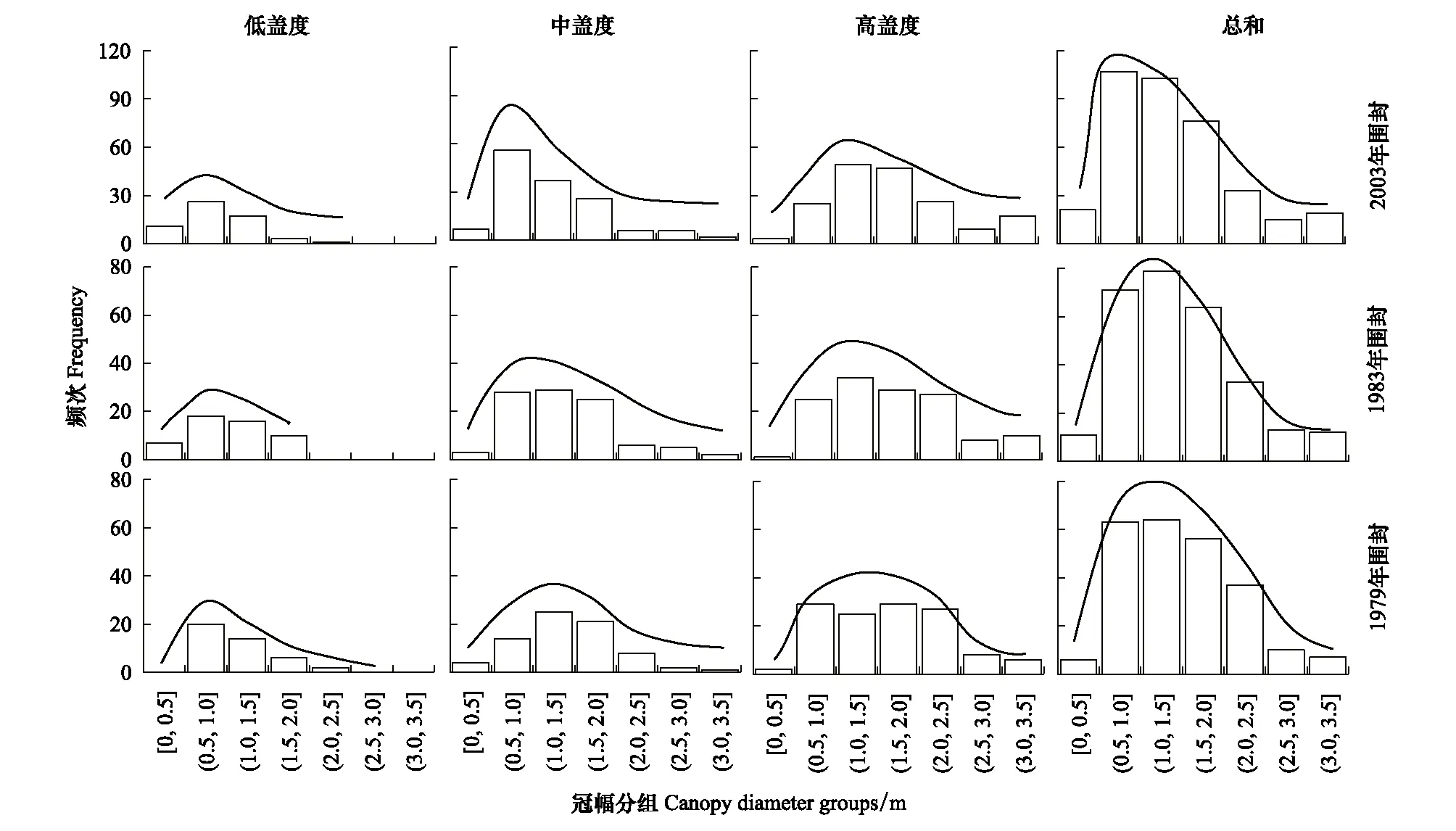
图1 小叶锦鸡儿冠幅频度分布Fig.1 Frequency distribution of canopy diameter of C. microphylla
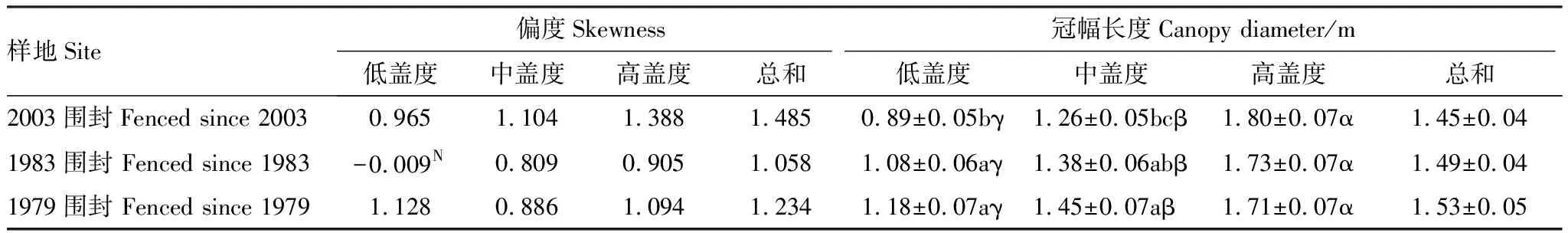
表1 小叶锦鸡儿灌丛大小及分布结构
偏度值后面N,表示该组数据满足正态分布,未标注表示数据不满足正态分布;同一列平均值间,含有相同的英文字母表示围封年限间差异不显著;而同一行,含有相同希腊字母表示不同小叶锦鸡儿盖度间差异不显著方差分析结果显示围封年限、灌丛化盖度及其交互作用均对小叶锦鸡儿冠幅大小具有显著影响(P<0.05)。简单效应结果显示,同一围封年限下,随着灌丛化盖度水平增加,小叶锦鸡儿冠幅显著增大;在低盖度样区内,1979年和1983年围封样区内的小叶锦鸡儿冠幅显著高于2003年围封样区;在中盖度区,1979年围封样区内的小叶锦鸡儿冠幅显著高于2003年围封样区,两者均与1983年围封样区所得结果无显著差异;在高盖度区,不同围封年限间无显著差异(表1)。
2.2 物种重要值及生态位宽度
在1979年围封样地中,多年生禾草羽茅(Achnatherumibiricum)的重要值最高,为0.253;多年生禾草大针茅次之,为0.144。在1983和2003年围封样地中,多年生杂类草黄囊苔草(Carexkorshinskyi)的重要值最高,分别为0.298和0.379;多年生杂类草二裂委陵菜(Potentillabifurca)次之,分别为0.176和0.151(表2)。
在1979围封样地中,生态位宽度高于1的物种多出现在多年生禾草功能群内,包括冰草(Agropyroncristatum)、大针茅、羊草和羽茅,还有多年生杂类草中的二裂委陵菜和黄囊苔草;在1983围封样地中,生态位宽度高于1的物种分别为多年生禾草大针茅和羊草,多年生杂类草二裂委陵菜、黄囊苔草、一年生植物轴藜(Axyrisamaranthoides)和半灌木木地肤(Kochiaprostrata);在2003围封样地中,生态位宽度高于1的物种分别为多年生禾草大针茅,多年生杂类草二裂委陵菜、黄囊苔草和一年生植物轴藜(表2)。
2.3 群落整体关联性分析
基于方差比率VR及其检验结果,可以得出2003年围封样地中,VR值为0.764,群落中主要物种间的整体关联为轻微负关联;而1983和1979年围封样地中所得VR值均高于1,分别为1.347和1.039,主要物种间的整体关联为轻微正关联(表3)。
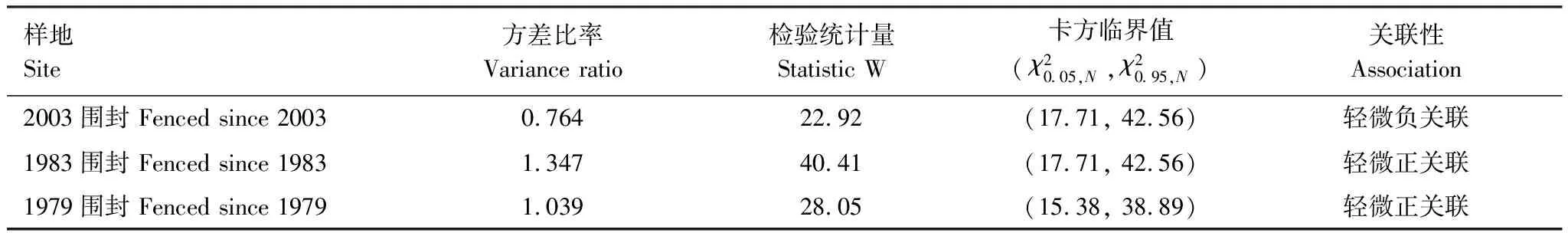
表3 小叶锦鸡儿灌丛间群落主要物种种间整体关联性分析
3 讨论
在干旱半干旱草原区,对于小叶锦鸡儿灌丛化草地的增加,很早就引起了生态学研究者的关注,然而关于小叶锦鸡儿灌丛化草地是否为该区域的另一种稳态的观点并没有一致的结论[9, 24-25]。本课题组在2014年报道了基于围封5—8 a的实验结果,发现随着围封时间的延长,小叶锦鸡儿种群盖度有所降低,且得出小叶锦鸡儿种群盖度与多年生禾草的重要值呈显著负相关关系,并基于此结果推测小叶锦鸡儿种群开始衰退[25]。本实验在围封15—39 a的样地内开展的研究发现,围封仅使得小于0.5 m冠幅的小叶锦鸡儿的数量降低或者消失,进而使得种群的冠幅随围封年限的延长呈现出增大的趋势,这种趋势在小叶锦鸡儿低、中盖度样区内尤为明显,支持了前一研究中小叶锦鸡儿种群盖度降低的结论。另外,本研究还发现多年生禾草重要值随着围封年限的增加而增加,从生态位配置方面为小叶锦鸡儿种群盖度与多年生禾草的重要值显著负相关提供解释,同时还可以为小于0.5 m小叶锦鸡儿的降低或消失与灌丛间草本植物对入侵灌木的竞争有关提供解释。
在围封恢复早期,放牧压力的去除使得灌丛间草本植物补偿性生长,迅速占据空余生态位,并抑制了具有较小冠幅的小叶锦鸡儿的生长[25-26];随着围封年限增加,植物凋落物的积累和分解促进了土壤理化性质的改善(如土壤氮、磷含量的增加),多年生禾草因其自身较高的氮素利用率及快速生长能力,从而在围封草地植物群落中逐渐占据优势地位[27]。Pierce等在美国新墨西哥州南部的乔纳达实验区对灌丛化草地的研究发现,草本植物对冠幅小于0.5 m的灌木生长表现出明显的抑制作用[28]。另外,在美国亚利桑那州东南部一项长期观测实验发现,当灌木达到一定冠幅后,灌木相对于草本有竞争优势,且不会被竞争排除[7]。在我们的实验结果中,8/9的样区内小叶锦鸡儿种群分布表现为正偏态分布,表明种群结构为上升型,但同时表现为小于0.5 m冠幅的缺失,即缺乏小个体级的补充,这说明不同围封年限下小叶锦鸡儿种群实际处于一种相对稳定的状态。综合相关实验,我们得出当小叶锦鸡儿冠幅较小(<0.5 m)时,围封促进草本植物的快速生长并作为强竞争者竞争排除小叶锦鸡儿,可以通过围封促使灌丛化草地转变为典型草原;而当小叶锦鸡儿冠幅较大(>0.5 m)时,仅自然条件下的围封而无外源能量的输入,草本植物对灌木的竞争作用将不足以促使灌丛化草地向多年生禾草典型草原的转变[8,29],即小叶锦鸡儿灌丛化草地为该区域的另一种稳态。
基于种间关联理论,种间负关联通常表现为物种间对于相似环境资源的需求所产生的竞争作用;而正关联通常表现为物种分属不同生活型或具有不同资源需求从而产生的生态补偿效应[30-31]。本研究发现,随着围封时间的延长,小叶锦鸡儿灌丛间群落整体关联关系由负关联向正关联转变,表明该群落内植物对相同资源的竞争利用得到缓解,群落内多数物种种间竞争作用弱化或表现为促进作用[30]。龚容对围封条件下温带半干旱典型草原群落种间关联的研究发现,围封能够显著增加群落内的正关联物种对,有利于群落的稳定[30];单贵莲等的研究发现随围封年限的延长,群落物种多样性及均匀度增加[31]。在本研究中,2003年围封样地内黄囊苔草的重要值最大(0.379),显著高于群落内其他物种;而随着围封年限的延长,单种优势作用逐渐弱化。这些研究均表明当群落内物种达到一定数量后,对资源的吸收和利用达到平衡,种间资源竞争会趋于平稳,群落更趋于稳定[32]。另外,资源竞争的减弱也与群落内物种生态位宽度的变化有关,随着围封年限的延长,群落由多年生杂类草占主导的生态位配置向以多年生禾草和多年生杂类草共同主导的生态位配置转变,而群落生态位配置的改善,有利于种间关系和群落结构的稳定[19]。
通过对围封15—39 a的小叶锦鸡儿灌丛化草地的研究观测,一方面从关键种小叶锦鸡儿种群大小结构方面给出了小叶锦鸡儿种群的发展动态,另一方面从群落主要物种的生态位配置及种间关联方面阐述了围封对灌丛间群落演替的影响。所得结果为合理评价和定位小叶锦鸡儿灌丛化草地的状态和演替规律提供了数据支持,也为制定合理的灌丛化退化草地恢复措施提供了理论支持。
