川西亚高山不同林龄粗枝云杉人工林土壤微生物生物量及酶活性
2021-09-01罗明霞胡宗达刘兴良李亚非欧定华吴德勇
罗明霞, 胡宗达,*, 刘兴良, 李亚非, 胡 璟, 欧定华, 吴德勇
1 四川农业大学资源学院, 成都 611130 2 四川省林业科学研究院, 成都 610081 3 湖南农业大学资源环境学院, 长沙 410128
土壤微生物在土壤养分供给和转换过程中扮演着十分重要的角色,其生物量(土壤中体积小于5×103μm3的生物总量)属于土壤中最活跃和最易变化的有机质部分[1],是植物生长所需物质和能量的重要来源,也是驱动土壤养分循环的主要动力,能很好地反映微生物在土壤中的实际含量和作用潜力[2- 3]。土壤酶作为有机质分解、周转和矿化的主要生物催化剂,在土壤物质循环过程中发挥着重要的驱动作用,其活性能代表土壤中碳氮代谢的旺盛程度,亦能表征微生物对土壤养分的吸收利用状况[4-5]。土壤微生物生物量和酶活性对土壤底物有效性变化较为敏感[6],可综合表征土壤微生物活性[7],以反映土壤质量在时间序列或不同环境条件下的变化,是监测土壤底物有效性变化和评价土壤肥力的重要指标[8]。
土壤酶主要来源于土壤微生物、根际分泌物和动植物残体分解,参与土壤生物化学循环的主要酶类为氧化还原酶和水解酶[9],水解酶(如β-葡萄糖苷酶、纤维素酶和β-N-乙酰氨基葡萄糖苷酶等)和氧化酶(如过氧化物酶和多酚氧化酶等)分别与土壤有机物质分解和腐殖化过程紧密相关[10],其酶活性差异可反映微生物养分的需求变化[11]。研究显示,以针叶树种为主的人工林地土壤多酚氧化酶活性高于以阔叶为主的天然次生林,而过氧化物酶无显著差异,β-葡萄糖苷酶和纤维素酶活性则相反[12]。在人工林生态系统中,林龄是影响土壤养分分配的重要因素,可通过改变林分结构引起凋落物、根系分泌物、理化性质(水分、有机质含量、pH等)和微气候等的变化,造成微生物种类和组成不同,进而影响土壤酶活性[13-14]。目前的大量研究表明,人工林土壤酶活性随着林龄的增长表现出增加[15]、先增后降[16]、先增后降再增[17]的变化趋势均有,随林龄增加土壤酶活性变化差异显著,主要源于研究区域、物种组成、植物学特性和森林管护方式不同所致[18-19]。
川西亚高山森林是我国重要的水源涵养和水土保持林,是维护长江上游生态安全的绿色屏障[20]。20世纪50年代以来,川西亚高山原始暗针叶林被大规模采伐,至1998年停采封存,已形成天然和人工更新共同作用下的原始暗针叶林-天然次生林-人工林等不同林型镶嵌分布的森林格局[21]。云杉(Piceaasperata)作为中国西部和西南亚高山地区人工林主要树种之一,在保障西北地区和青藏高原东南部生态安全方面有着重要作用。近年来,对该区域云杉人工林的研究主要集中在土壤碳动态[22]、土壤呼吸响应机制[23]和土壤微生物群落结构[24]等方面,对土壤酶活性多侧重于冻融循环[25]、氮沉降[26]等因素对酶活性的影响,而对不同林龄人工林林下土壤微生物生物量和酶活性及其影响因素鲜见报道。本研究采用空间代替时间的方法,选择川西亚高山米亚罗林区不同林龄粗枝云杉(Piceaasperata)人工林,探讨林下表层土壤微生物生物量及土壤酶活性对不同林龄的响应,分析土壤酶活性与土壤理化性质和有效养分的关系,旨在揭示川西亚高山粗枝云杉人工林随造林时间延长其土壤质量的变化特征及其影响因素。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于四川省阿坝州理县米亚罗林区(31°24′—31°55′ N,102°35′—103°4′ E),海拔梯度为2850—4200 m。地貌属于典型的高山峡谷地貌,气候为季风山地气候,夏季湿润多雨,冬季寒冷干燥。1月份平均气温—8℃,7月份平均气温12.6℃,年平均气温 6—12℃,≥10℃的年积温1200—1400℃,年降水量600—1100 mm,年蒸发量1000—1900 mm。森林土壤类型主要为山地棕壤,成土母质主要为千枚岩、板岩和白云岩等残积风化物,土层薄,石砾含量高[27-28]。研究样地选择在米亚罗林区不同年代采伐迹地、经过人工种植更新及实施封育后,不同恢复阶段的粗枝云杉人工林集中分布地段。主要物种为粗枝云杉(Piceaasperata),林下主要物种有疏花槭(Acerlaxiflorum)、三棵针(Berberissargentiana)、扁刺蔷薇(Rosasweginzowii)、陕甘花楸(Sorbuskoehneana)、高山柳(Salixparaqplesia)等。
1.2 样地设置与土样采集
2019年7月下旬,按照地域毗邻和生境相似性原则,在米亚罗林区选取25、40、50a和60a的粗枝云杉(Piceaasperata)人工林(见表1),每种林地随机布设3个20 m×20 m的标准样地,进行每木检测,并记录地理坐标、海拔、坡度、郁闭度等指标。然后对每块标准样地按照“田”字形分为4个10 m×10 m的样方,采用“梅花5点”法布点采集表层(0—20 cm)土壤样品,采样前小心除去地表枯落物,用不锈钢土钻(Eijkelkamp,型号 07.53.SC)进行取样,将所有土样充分混合均匀,按四分法分取1 kg样品放入密封袋,置于4℃保温箱运回实验室,立即去除动植物残体和石砾等杂质,将其土样分为2份,1份鲜样用于测定土壤微生物生物量碳氮和酶活性;另1份自然风干,分别过1、0.25 mm筛,用于测定土壤理化性质。
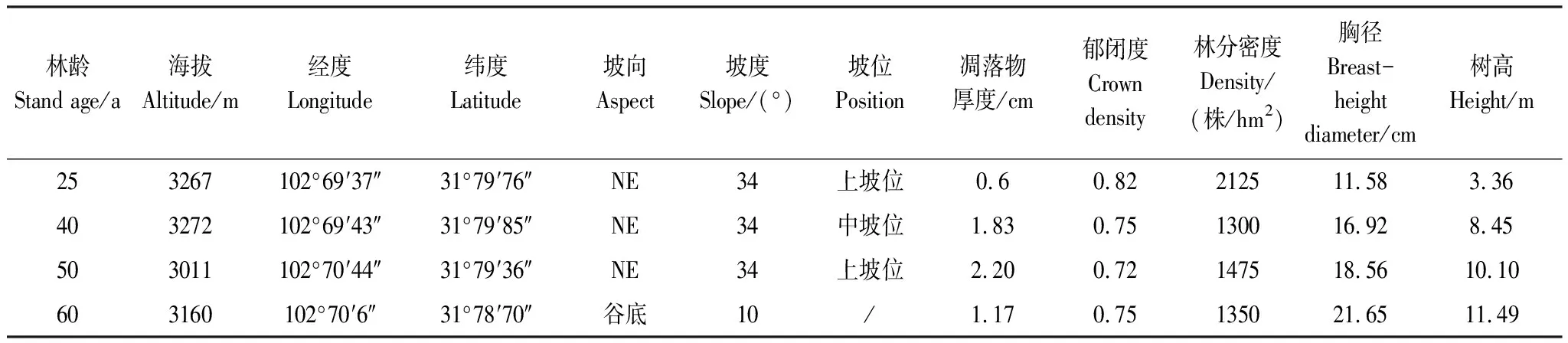
表1 粗枝云杉人工林样地基本情况
1.3 样品分析
1.3.1土壤基本理化性质的测定
土壤有机碳(SOC)和全氮(TN)分别采用重铬酸钾氧化外加热法、硫酸-催化剂消解流动分析仪法测定;全磷(TP)采用钼锑抗比色法,利用 UV- 2450紫外分光光度计测定;碱解氮(AN)采用滴定法测定;土壤pH值用赛多里斯PB- 10型pH计测定,水土体积质量比为2.5∶1;采用烘干法测定土壤质量含水率。
土壤可溶性有机碳(DOC)和可溶性有机氮(DON),用蒸馏水以水土比为5∶1浸提,室温振荡30 min,用0.45 μm滤膜抽滤,用日本岛津TOC-VCPH/CPN分析仪测定滤液中DOC含量,用紫外分光光度计(岛津 UV- 2450)测定可溶性总氮(DTN),用AA3-连续流动分析仪测定可溶性无机氮(DIN),利用差值法DON=DTN-DIN计算;轻组有机碳(LFOC)采用碘化钠浸提法,用比重为1.7 g/cm3的碘化钠溶液浸提,振荡1 h后离心,取轻质部分用0.45 μm 纤维滤膜过滤,再用0.01 mol/L CaCl2和蒸馏水抽滤冲洗,烘干(60℃)称量后过0.15 mm筛,最后用岛津TOC-SSM- 5000A 碳分析仪测定
1.3.2土壤微生物生物量碳(MBC)、生物量氮(MBN)浓度测定
微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸-K2SO4直接浸提法测定:称取2份7.5 g的过2 mm筛的新鲜土样,分别放置于有氯仿和无氯仿的真空干燥器中,在黑暗环境下内熏蒸24 h后取出,再分别加入土液体积比为1∶4的0.5 mol/L K2SO4水溶液,震荡30 min,过滤后,MBC用TOC-VCPH有机碳分析仪测定,MBN用AA3-连续流动分析仪测定[29],计算公式如下:
土壤微生物生物量碳 =(熏蒸土壤有机碳-未熏蒸土壤有机碳)/0.45
(1)
土壤微生物生物量氮 =(熏蒸土壤有机氮-未熏蒸土壤有机氮)/0.25
(2)
1.3.3土壤酶活性的测定
土壤酶活性的测定参照Chen等[30]和Li 等[31]的测定方法:葡萄糖苷酶采用荧光比色法,首先,取1 g新鲜土壤,加蒸馏水125 mL,震荡2 h(25℃ 180 r/min)后,吸取1 mL的悬浊液,加入荧光底物(伞形酮MUB)0.25 mL,25℃下避光培养4 h,再加入50 μL浓度为0.5 mol/L的NaOH溶液,反应终止后用多功能酶标仪(M200PRO, 瑞士Tecan)测定其荧光度;过氧化物酶和多酚氧化酶采用酶联免疫微量法,先取过50目的风干土样0.20 g,分别加入反应底物(120 mL和100 μL邻苯三酚溶液、20 μL过氧化氢溶液),振荡混匀,30℃恒温培养 1 h后取出,再分别加入50 mL磷酸缓冲液和430 mL乙醚、50 μL磷酸缓冲液和430 μL乙醚,振荡数次,25℃室温静置 30 min,用酶标仪(M200, 瑞士Tecan)测定;纤维素水解酶采用3,5-二硝基水杨酸比色法测定,称取10 g土壤,加入1.5 mL甲苯,摇匀后放置15 min,再加入5 mL质量分数为1%的羧甲基纤维素溶液和5 mL pH=5.5的醋酸盐缓冲液,于37℃恒温箱中培养72 h后,取1 mL滤液,用岛津-UV2450-紫外分光光度计测定。5种土壤酶的名称、缩写和功能见表2。
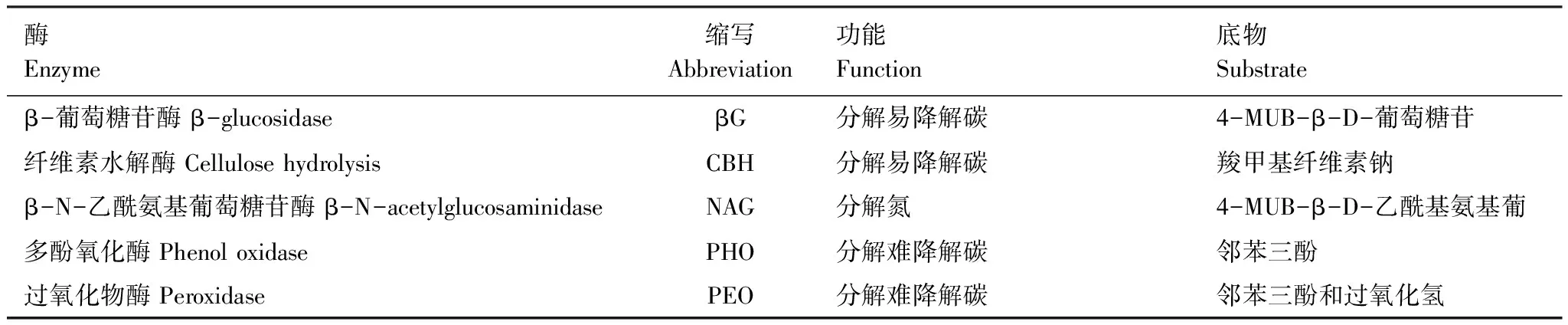
表2 土壤酶的名称、缩写、功能及底物
1.4 数据处理
采用单因素方差法(One-way ANOVA)和最小显著性差异法(LSD)检验不同林龄人工林土壤理化性质、土壤微生物生物量和土壤酶活性的显著性差异(α=0.05);利用Pearson相关性分析得到与土壤酶活性显著相关的土壤因子(数据未在文中列出);用Canoco Software 4.5软件对土壤酶活性和与其呈显著相关性的土壤因子进行冗余分析(Redundancy Analysis, RDA),探究影响土壤酶活性变化的主导因素。所有数据均采用SPSS 22.0软件进行统计分析,用SigmaPlot12.0软件制图。文中图表数据均为平均值±标准偏差。
2 结果与分析
2.1 不同林龄粗枝云杉人工林土壤基本理化性质的比较
不同林龄人工林土壤pH变幅为5.30—6.02,随种植时限增加呈显著增加趋势(P<0.05);40年生人工林的WC显著高于其他林龄阶段的人工林;随林龄增加,表层土壤SOC、TN、LFOC、AN和DOC呈现先增加后降低的变化趋势(表3),其中50年生人工林的含量最高。随着林龄的增加,DON也呈先增后减的变化趋势,但未达到显著水平(P>0.05)。土壤C/N随林龄增加呈显著下降的趋势,60年生人工林的C/N显著低于25年生和40年生人工林(P<0.05)。表层土壤TP含量在不同林龄人工林之间差异显著,表现为60a>25a>50a>40a。
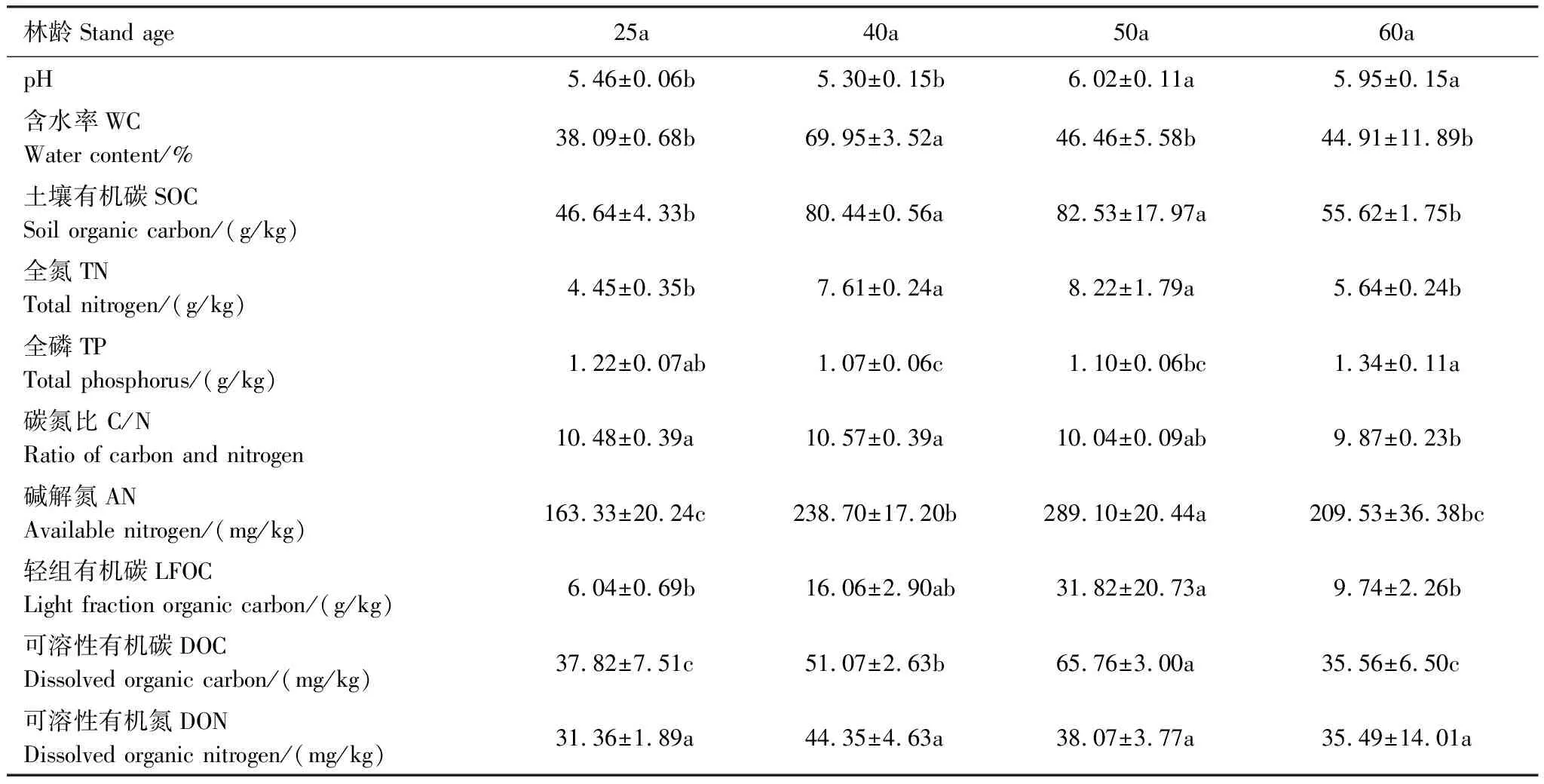
表3 不同林龄粗枝云杉人工林土壤理化性质
2.2 不同林龄粗枝云杉人工林土壤微生物生物量的比较
图1显示,不同林龄粗枝云杉人工林表层土壤MBC和MBN的含量变化趋势相似(P<0.05),其中40年生人工林的MBC和MBN含量最高(1408.50 mg/kg和437.94 mg/kg),相比25、50、60年生,分别高64.09%和52.92%、26.97%和34.71%、54.91%和70.19%,其高低排序分别为40a>50a>60a>25a和40a>50a>25a>60a。MBC/MBN随林龄呈显著增加趋势(图1),即60年生人工林分别比25、40、50年生高50.35%、31.73%和26.06%。
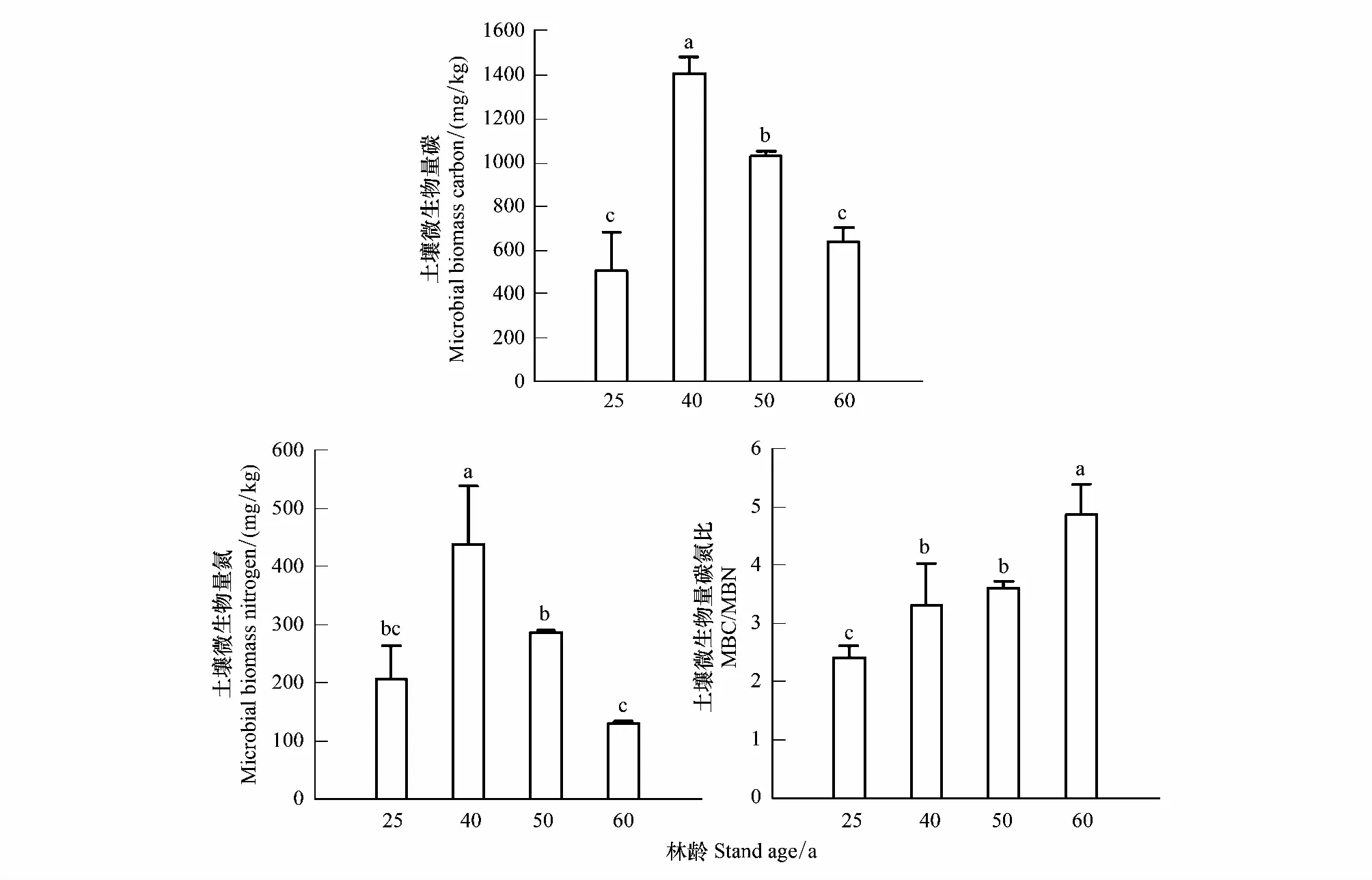
图1 不同林龄粗枝云杉人工林土壤微生物生物量碳氮含量变化Fig.1 The variation trend of soil microbial biomass contents and microbial C:N ratio in Picea asperata plantations with different ages (n=3)不同小写字母表示不同林龄粗枝云杉人工林间差异显著(P<0.05)
2.3 不同林龄粗枝云杉人工林土壤酶活性的比较
图2显示,人工林表层土壤βG活性表现为50a(30.65 nmol g-1h-1)>60a(22.71 nmol g-1h-1)>40a(21.74 nmol g-1h-1)>25a(5.28 nmol g-1h-1),其中40年生和60年生的βG活性差异不显著(P>0.05);CBH活性随林龄呈增加趋势,60年生显著高于其他3种林龄阶段的人工林(P<0.01),其中50a和40a、40a和25a之间差异不显著(P>0.05);NAG活性变化趋势与βG相似,50年生人工林表层土壤NAG活性分别较25a和60a的高77.03%和39.98%;PHO活性表现为40a>50a>25a>60a;50年生的PEO活性显著低于其他三种林龄阶段的人工林(P<0.05),分别比25、40和60年生低77.94%、63.88%和68.52%。
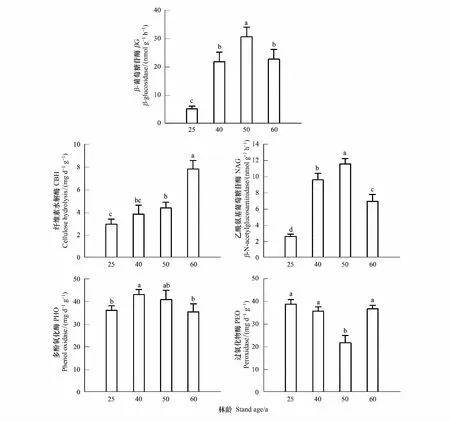
图2 不同林龄粗枝云杉人工林的土壤酶活性Fig.2 The variation trend of soil enzyme activities in Picea asperata plantations with different ages (n=3)
2.4 土壤酶活性变化的RDA分析
以5种土壤酶活性作为响应变量,选取与土壤酶活性具有显著相关关系的土壤因子作为解释变量,并结合不同林龄处理样点进行冗余分析,获得驱动土壤酶活性变化的主要因子。结果显示,不同林龄人工林样地表现出有规律的聚类(图3),说明林龄对人工林土壤酶活性的影响显著。从图3可看出,第一轴和第二轴共同解释了土壤酶活性变异的86.86%,AN、pH和DOC分别解释了土壤酶活性变异的65.4%(P=0.002)、9.7%(P=0.026)和7.6%(P=0.04),它们是引起土壤酶活性变异的显著性解释因子。AN与NAG、βG极显著正相关(P<0.01),与PEO显著负相关(P<0.05);pH与PEO显著负相关(P<0.05);DOC与NAG、βG、PHO显著正相关,与PEO显著负相关(P<0.05);NAG与βG极显著正相关(P<0.01)、与PHO显著正相关(P<0.05),PEO与NAG和βG极显著负相关(P<0.05)。
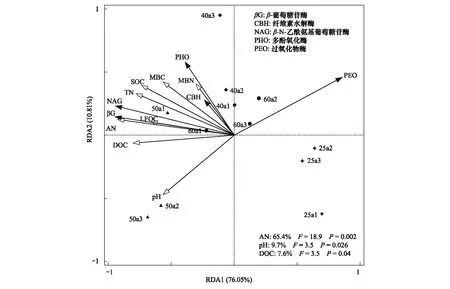
图3 不同林龄粗枝云杉人工林土壤理化性质与土壤酶活性冗余分析Fig 3 Redundancy analysis (RDA) depicting the relationship between soil physical-chemical properties and soil enzyme activities in Picea asperata plantations with different ages (n=12)25a1、25a2、25a3表示25年生粗枝云杉人工林样地的3个平行重复25a1, 25a2, 25a;pH:酸碱度; SOC:土壤有机碳;TN:全氮;AN:碱解氮;LFOC:轻组有机碳;DOC:可溶性有机碳;MBC:微生物生物量碳;MBN:微生物生物量氮;图右下角变量注释表示对土壤酶变化有显著影响作用的环境因子的相应解释比例
3 讨论
3.1 不同林龄人工林土壤理化性质变化
林龄或林分发展阶段是森林生态系统发育的结果,林龄对人工林土壤理化性质有着显著影响。土壤pH与凋落物的量和凋落物分解速率、凋落物养分归还率密切相关。本研究中,随着粗枝云杉人工林林分的生长发育,土壤pH显著增加,表明种植年限可有效改变土壤酸碱性,已有研究结果亦有证实[32]。各林龄粗枝云杉人工林C/N随林龄增大而减小,这是由于土壤有机碳和全氮的变化趋势虽然一致,但土壤有机碳的变化趋势滞后于土壤全氮的变化趋势。土壤C、N均是植物生长所必需的大量元素,在植物生长发育过程中起着重要作用。本研究发现SOC、TN、AN、DOC和LFOC等养分含量均随林龄呈先增后减的变化趋势,于近熟林(40—50a)达到最大,与已有研究结果一致[33]。人工林生态系统处于幼龄林时,植物为了满足自身生长需要,从土壤中吸收大量的养分导致土壤养分含量呈现出较低水平;人工林到达成熟期时,养分循环周期长、归还速率慢,更多的消耗林地的养分来维持自身的生长,林地土壤养分含量亦低,故幼龄林和成熟林时期人工林的生物量、营养元素积累量和年存留量较低。土壤全磷含量的高低在一定程度上反映了土壤的供磷潜力,当全磷含量低于0.8—1.0 g/kg时,土壤常出现供磷不足[34]。本研究结果显示川西亚高山不同林龄粗枝云杉人工林全磷含量为1.07—1.34 g/kg,并随着林龄的增加呈V型趋势变化,40—50年生显著低于25年生和60年生的人工林,可能是因为40—50 a的粗枝云杉处于快速生长阶段,导致树木对土壤磷素的吸收量大于土壤全磷的增加量。
3.2 不同林龄人工林土壤微生物生物量变化
随林分发育土壤微环境发生变化,使得土壤微生物生物量对环境变化的敏感性随之改变。本研究结果显示,表层土壤MBC和MBN含量变幅分别为505.73—1408.50 mg/kg、130.54—437.94 mg/kg,与谭波等[35]对该区域同海拔同时段冷杉林的研究结果相似,高于贺若阳等[36]对该区域云杉人工林有机层的研究结果。表层土壤MBC和MBN随林龄增加均呈现出先增后降的显著变化趋势,与Wang等[37]的研究结果一致,但与本研究区域境内不同林龄(9、13、31a)粗枝云杉(Piceaasperata)人工林的研究结果有所差异[38],可能是由于研究土层选择不同所致。粗枝云杉人工林生长到40—50a阶段,表层土壤MBC和MBN显著高于25年生和60年生,可能是因为40—50年生阶段人工林林下地表凋落物量和通过凋落物归还土壤的营养元素的年归还总量高于25a和60a(表1),且地表温湿度适宜微生物繁殖,提高了微生物活性,使微生物分解有机质增强,故土壤微生物生物量含量较高,这在已有研究中得到证实[39]。亦有研究表明,土壤养分含量是影响土壤微生物生物量的重要因素[40],本研究25年生和60年生人工林的SOC、TN、AN等养分含量显著低于40—50年生,可能是导致该林龄土壤MBC和MBN含量低的原因。MBC/MBN作为土壤微生物种类组成的一个重要指标,可指示土壤真菌与细菌的相对丰度,其比值越高,说明土壤中真菌数量越多[41]。土壤MBC/MBN处于3—6之间,土壤细菌占优势,反之则以真菌为主[42]。本研究中粗枝云杉人工林土壤MBC/MBN值随林龄增加而显著增大(P<0.05),为2.42—4.87(均值3.55),低于川西亚高山粗枝云杉(Piceaasperata)人工林(10.56—13.77)[36]及温带森林(7.4—7.9)[43]的MBC/MBN比值,说明随林龄增加,研究区人工林土壤微生物真菌数量有所增加但仍以细菌为主。
3.3 不同林龄人工林土壤酶活性变化
土壤微生物酶可催化土壤发生发育及肥力的形成,其活性高低可指示土壤微生物活性的强弱,可作为表征土壤质量的重要生物学指标[44]。本研究发现,随粗枝云杉人工林种植年限的增加,表层土壤与碳氮相关水解酶活性(NAG和βG)呈先增后降的趋势,均于50年生达到最大(图2),不同于该区域云杉人工林的研究结果[27],可能是林地微环境不同所致。土壤中可利用养分为土壤微生物及水解酶生产供应能量,故土壤养分含量高的林分,土壤水解酶活性也高[45]。白桦林在中龄林与近熟林阶段酶活性最高,土壤有机质也在这个时期周转最快[46];日本落叶松在11年和47年酶活性较高,此阶段人工林养分含量也较高[47],因而50年生人工林较高的SOC、TN、DOC、AN和LFOC含量促进了NAG和βG的活性;而随林龄增加CBH呈逐渐增加的变化趋势,有研究证实林龄会引起林分中温度、湿度以及凋落物现存量、厚度的差异[48],这种结果进一步表明森林生态系统的复杂性。此外,与分解难降解碳相关的氧化酶(PEO和PHO),25年生和60年生无显著差异,表明PHO、PEO活性变化较小。可能是由于粗枝云杉人工林凋落物中木质素、粗纤维、多酚类等难溶性物质含量较高,种植密度(1300—2125 株/hm2)和郁闭度较大(0.72—0.82),且温带森林凋落物年均分解速率仅为热带森林的1/3,亚热带森林的1/2左右[49-50],故研究样地凋落物引起土壤难降解碳含量的变化较小。因此,研究区人工林氧化酶活性受林龄的影响较小,与已有研究结果相似[51]。
3.4 驱动土壤酶活性变化的关键因子
土壤酶作为土壤中众多生化反应的主要催化物,其活性与温度、水分、pH 、养分有效性、土壤微生物群落结构以及植物种类等因素密切相关[52]。在本研究中发现AN是人工林表层土壤酶活性的主要影响因子,解释量高达65.4%,与已有研究结果相似[53];冗余分析显示,AN与NAG和βG显著正相关,不仅反映了底物与酶活性之间的正相关关系,也说明了土壤N素有效性促进了与土壤C循环相关的酶活性[54]。陈倩姝等[55]研究表明,在我国西部地区的亚高山森林生态系统普遍受氮限制的背景下,增施土壤氮肥可有效促进土壤NAG和PEO活性。土壤AN是铵态氮、硝态氮、氨基酸、酰胺和易水解的蛋白质的总和,能有效衡量土壤供氮能力,在川西亚高山森林土壤氮限制背景下,AN是研究区人工林表层土壤酶活性变化的主要影响因素,与已有研究结果相似[24]。土壤pH的高低,不仅影响土壤微生物的种类、活性和氧化还原反应等过程,而且对土壤酶参与生化反应的速率有直接影响。有些酶促反应对 pH 值变化很敏感,研究显示,当 pH 值超过其最适宜范围时,就会对酶活性产生抑制作用[56]。本研究中,pH与PEO活性显著负相关,对土壤酶活性变化解释了9.6%,亦是影响土壤酶活性的重要因素之一。此外,DOC也是影响土壤酶活性的显著性解释变量,随林龄增加,DOC先增后减,与参与C、N循环相关的NAG、βG、PHO呈现正相关关系,与PEO呈负相关,有研究表明土壤微生物利用有机碳往往有一定的选择性, 更倾向于利用易分解的有机质[12],与本研究结果发现DOC对土壤酶活性影响较大的结果相似。可溶性有机物是森林生态系统土壤养分迁移及损失的主要形式,既可源自于土壤微生物的生命活动,也可来源于有机物质的直接溶解[7],本研究中DOC与MBC变化趋势较一致,说明人工林土壤中DOC含量可能更多地来自土壤微生物生命活动。
4 结论
不同林龄粗枝云杉人工林表层土壤微生物生物量和酶活性存在显著差异;土壤养分含量(SOC、TN、AN、DOC、LFOC)、微生物生物量(MBC、MBN)和酶活性(NAG、βG、PHO)总体上随林龄增加呈先升后降的显著变化趋势,40—50a显著高于25年生和60年生阶段,说明此生长阶段的粗枝云杉人工林具有较好的土壤C和N周转速率。AN、pH和DOC是驱动土壤酶活性变化的主要因素,这源于土壤微生物更倾向于利用土壤中易分解的有机碳和氮。因此,在粗枝云杉人工林经营管理中应适当增施氮素肥料及凋落物维护,以提高土壤酶活性,进而提升土壤质量,以保证林木良好生长。
