高分泌型K326近等基因系的创制和鉴定
2021-08-04任书乐王召军闫筱筱张洪映李雪君孙计平崔红
任书乐,王召军,闫筱筱,张洪映,李雪君,孙计平,崔红*
1 河南农业大学,烟草学院,郑州市文化路95号 450002;
2 河南省农业科学院烟草研究所,河南省许昌市魏都区青梅路与永昌大道交叉口 461000
烟草腺毛丰富,其分泌物的主要成分为二萜化合物和蔗糖酯[1],二者都是烟草重要的香气前体物质[2-3]。在不同类型烟草中,香料烟腺毛分泌物含量为最高,其次是雪茄烟,烤烟普遍较低[4]。二萜化合物包括西柏烷化合物和赖百当化合物两大类,西柏烷二萜在不同类型烟草中广泛存在,赖百当二萜则主要存在于香料烟和雪茄烟中[5]。烟草蔗糖酯根据分子量及酯化位置的不同可以分为六种类型(I-VI),但III-VI型蔗糖酯只在香料烟和雪茄烟中存在,烤烟一般只含有I-II型蔗糖酯[4,6]。可见,腺毛分泌物与烟叶香气品质和风味密切相关。
腺毛分泌物含量由腺毛密度、腺毛类型、分泌能力所共同决定。目前,本氏烟细胞周期蛋白基因NbCycB2对腺毛密度的调控机制已取得突破性进展[7],烟草NtCycB2基因对烟草腺毛发生的负向调控作用也已得到证明,K326中NtCycB2基因敲除可以显著提高分泌型腺毛的密度和分泌物的含量[8-9]。但是,关于烟草腺毛类型、腺毛分泌能力的分子机制尚不明确,还无法进行精确的分子定向改良。T.I.1068是美国农业部收集的一份野生烟草种质。其腺毛类型为分泌型,腺毛密度高,分泌能力强,分泌物成分齐全,包括西柏烷二萜、赖百当二萜及I-VI型蔗糖酯,分泌物可达烟叶干重的16%[10],是高分泌型烟草品种选育及腺毛发育及物质代谢相关基因克隆的理想材料。
K326是广泛栽培的烤烟品种,但因育成年代较早,其烟叶品质也需要进一步改良,以适宜烟草行业发展及消费者的需求。在不同烤烟品种腺毛分泌物的比较研究中,K326腺毛分泌物含量高于红大、翠碧1号和中烟100[11],但仍远远低于香料烟和雪茄烟品种[12]。由此推测,进一步提高腺毛分泌物的含量,有可能是K326品质改良的新的突破口。为此,本研究以K326为轮回亲本、以高分泌型种质资源T.I.1068为供体亲本进行多代回交,拟通过腺毛形态、叶面分泌物成分及农艺性状的综合定向筛选,获得高分泌型K326近等系,为K326叶面分泌物定向调控和品质改良奠定基础。
1 试验材料和方法
1.1 试验材料
供试材料为烟草品种K326和高分泌烟草种质T.I.1068。种子由本实验室和河南省农科院烟草研究所提供,品种杂交与回交试验地点设置在河南省农业科学院烟草研究所试验田(河南省许昌市),按照当地优质烤烟栽培规程进行田间管理。
1.2 试验方法
1.2.1 材料的选育过程
以K326为轮回亲本,以高分泌型种质资源T.I.1068为供体亲本,进行杂交和连续回交,对回交后代进行植株表型及腺毛形态的筛选,BC5F1与K326的农艺性状已趋于一致。BC5F1自交后,选取长柄分泌型腺毛腺头增大,罗丹明染色加深的BC5F1单株,进行腺毛分泌物成分的检测分析,从中获得了高分泌型K326近等系HSK326(图1)。
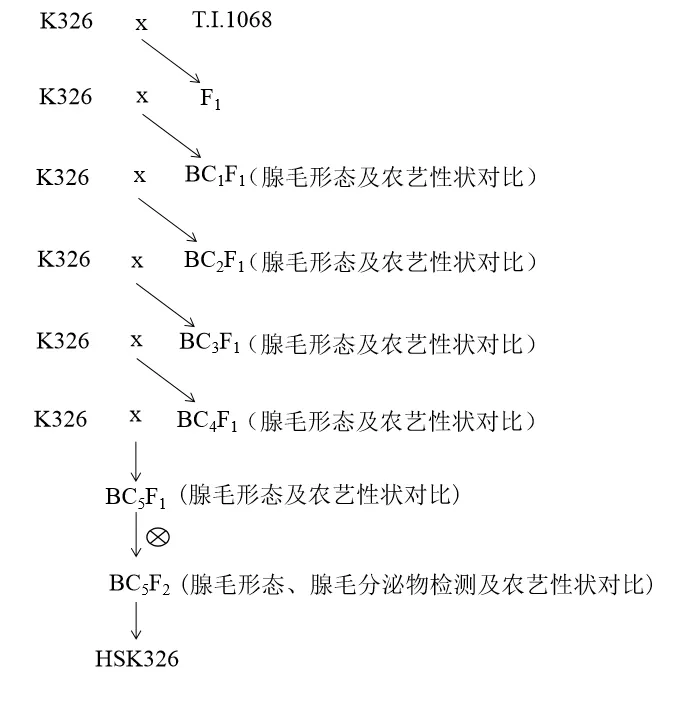
图1 HSK326选育过程Fig.1 HSK326 selection process
1.2.2 腺毛形态的观察
于烟株旺长期取顶部10 cm叶片进行腺毛染色与形态观察。试验操作参考李艳华等[11]使用的方法,将叶片置于0.2%(W/V)的罗丹明B水溶液中浸染60 min,染色结束后依次放到4个装有清水的烧杯中,每次5 s,涮去未结合的染料,用滤纸吸去多余水分,并置于室温下,自然晾干。用剪刀在叶片中部沿主脉两侧1 cm处剪取6 mm×8 mm组织块,置于超景深显微镜(VHR-5000,日本基恩士公司)下进行长柄分泌型腺毛腺头大小以及罗丹明染色的观察和长柄分泌型、短柄分泌型以及非分泌型腺毛的密度统计。
1.2.3 叶面分泌物检测
于烟株盛花期取下部5~10叶位叶片进行分泌物的提取和检测,样品制备以及前处理参考韩锦峰[13]的方法。每株材料取5片叶,每片叶取直径为7 cm的叶圆片4片,每株材料共20片叶圆片。将叶圆片在二氯甲烷溶液中浸提,每个叶圆片浸提8次,每次2 s。在浸提液中加入1 mL内标(2.020 mg/mL的蔗糖八乙酸酯和2.542 mg/mL的正十七烷醇的混合溶液),混合均匀后进行过滤,过滤后进行旋转蒸发仪浓缩,硅烷化处理后,利用气相色谱质谱联用仪TRACEGCULTRA—DSQ IIMS(美国赛默飞世尔科技公司)进行检测,色谱参数检测参考王霄龙等[14]的方法。采用内标定量法(相对校正因子为1)进行定量分析。
1.2.4 qRT-PCR 分析
在五叶一心期,选取生长状态一致的K326、T.I.1068及HSK326各一株,取自下往上第三叶位且发育正常的叶片,采用Trizol法进行RNA的提取。利用M-MLV Reverse Transcriptase试剂盒(美国Invitrogen公司)将提取的RNA反转录为cDNA。以烟草核糖体蛋白基因L25为内参基因,采用实时荧光定量PCR(qRT-PCR)对腺毛中西柏烷二萜和赖百当二萜合成基因表达水平进行定量分析,每个材料进行3次技术重复,所用引物见表1。qPCR采用LightCycler® 480 SYBR Green Ⅰ Master试剂盒,具体反体系如下:SYBR Green Mix 10.0 μL,上、下游引物(10.0 μmo/L)各0.5 μL,cDNA 1.0 μL,RNase free H2O 8.0 μL。在LightCycler® 480 Ⅱ型 荧 光 定量PCR仪上进行反应,反应条件为:95℃ 10 min;95℃ 15 s,60℃ 15 s,72℃ 30 s,40次循环,扩增结束后,添加溶解曲线程序:95℃,15 s,60℃,15 s,20 min内逐步升温至95℃,最后95℃,15 s。采用2-ΔΔCt法计算基因的相对表达水平。

表1 扩增引物序列Tab. 1 Amplification of primer sequences
1.2.5NtCPS2基因测序分析
在八叶一心期取K326、T.I.1068及HSK326发育正常的叶片,采用CTAB法进行DNA的提取。根据NtCPS2基因第4外显子中G/T SNP的侧翼序列设计特异性引物(F:TGGTGTTTTGATTGGCGAGTG;R:GGGGGCATAAAGGTGTCA)进行PCR扩增,反应条件为:94℃10 min;94℃ 30 s,58℃ 30 s,72℃30 s,35次循环;72℃延伸5 min;4℃保存。回收扩增产物进行序列测定,利用BioEdit 7.0 软件对测序结果进行序列比对。
1.2.6 SSR分子标记鉴定
在八叶一心期取K326、T.I.1068及HSK326发育正常的叶片,采用TIANGEN Plant Genomic DNA Kit试剂盒(离心柱型)进行DNA提取。利用BMVSE基因紧密连锁的分子标记PT30354(F:TCGGTTTCTGCTCCAATTTC;R:TGTTCTACTGCTGG GTCGAA)和PT20315(F:ACACGACTTTTCATCTCCC;R:CGCATGAAATT GTAAGGG-30)对所提DNA进行PCR扩增[18]。取PCR扩增产物1 µL上样于8%非变性聚丙烯酰胺凝胶单泳道,选用宝日医生物技术(北京)有限公司DL系列500 bp DNA Markers 作为参照,在1×TBE电泳缓冲液中电泳分离。电泳结束后,参照任民等[15]改良的NaOH银染方法快速显影。试验所用电泳仪器为DYY-12C 型(北京市六一仪器厂),仪器运行参数为恒电压1200 V,电流200 mA,功率60 W,电泳2 h。
2 结果与分析
2.1 高分泌型K326的选育和形态比较分析

图2 株型及腺毛形态比较Fig.2 Comparison of botany properties and trichome morphology
K326与T.I.1068的植株表型明显不同。K326株型为筒形或塔形,叶形为长椭圆形,叶数24~26片,茎叶夹角较大(图2-1);T.I.1068株型为塔形,叶形为披针形,叶数28~30片,茎叶夹角较小(图2-2)。二者叶面腺毛形态也具有明显差异。K326腺毛类型为混合型,以长柄分泌型为主,也有部分短柄分泌型和少量的非分泌型;长柄腺毛腺头较小,经罗丹明染色后呈现浅粉色(图2-4)。T.I.1068腺毛为分泌型,包括长柄分泌型和短柄分泌型,缺少非分泌型;长柄腺毛具有明显的分泌囊,经罗丹明染色后呈现玫红色(图2-5)。筛选获得的高分泌型K326近等系HSK326的株型为塔形,叶形为长椭圆形,叶数24片,茎叶夹角较大(图2-3);HSK326腺毛类型仍为混合型,但非分泌型腺毛比K326比例明显减少,长柄分泌型腺毛腺头明显增大,罗丹明着色加深(图2-6)。从腺毛密度统计来看,HSK326腺毛总量介于K326与T.I.1068之间。与K326相比,HSK326腺毛密度提高了0.3倍。其中,长柄分泌型腺毛密度提高了0.38倍,而非分泌型腺毛密度有所降低(图3)。

图3 腺毛密度比较Fig.3 Comparison analysis of trichome density
2.2 高分泌型K326叶面分泌物比较分析
在盛花期分别取K326、T.I.1068及HSK326的下部叶片,进行叶面分泌物的萃取和GC/MS分析,结果如图4所示。从组分上来看,HSK326与K326叶面分泌物组成成分一致,与T.I.1068具有较大差异。T.I.1068腺毛分泌物由西柏烷类二萜、赖百当类二萜、烷烃类和I-V型蔗糖酯构成;K326和HSK326的腺毛分泌物则只含有西柏烷类二萜、烷烃类以及I-II型糖酯,缺少赖百当类二萜和III-V型糖酯。从含量上看,T.I.1068叶面分泌物总量远远高于K326,HSK326介于二者之间。HSK326的分泌物总量是K326的1.88倍,其中,西柏烷类二萜提高了0.91倍,蔗糖酯含量提高了1.27倍,烷烃含量并无显著变化。
2.3 高分泌型K326二萜及蔗糖酯合成基因分析
2.3.1 二萜合成基因表达水平及序列分析
采用qRT-PCR技术,对K326、T.I.1068以及HSK326中赖百当类二萜合成基因NtABS、NtCPS2以及西柏烷类二萜合成基因NtCBT、NtCYP71D的表达水平进行了定量分析,结果如图5所示。可以看出,T.I.1068中二萜合成基因的表达水平普遍高于K326;HSK326中西柏烷二萜合成关键基因NtCBT显著高于K326,这与其西柏烷类化合物的大量积累相一致。赖百当二萜合成关键基因NtCPS2虽在T.I.1068中表达占据绝对优势,但在HSK326和K326中也有一定的表达量,但二者中并没有检测到赖百当二萜的积累。
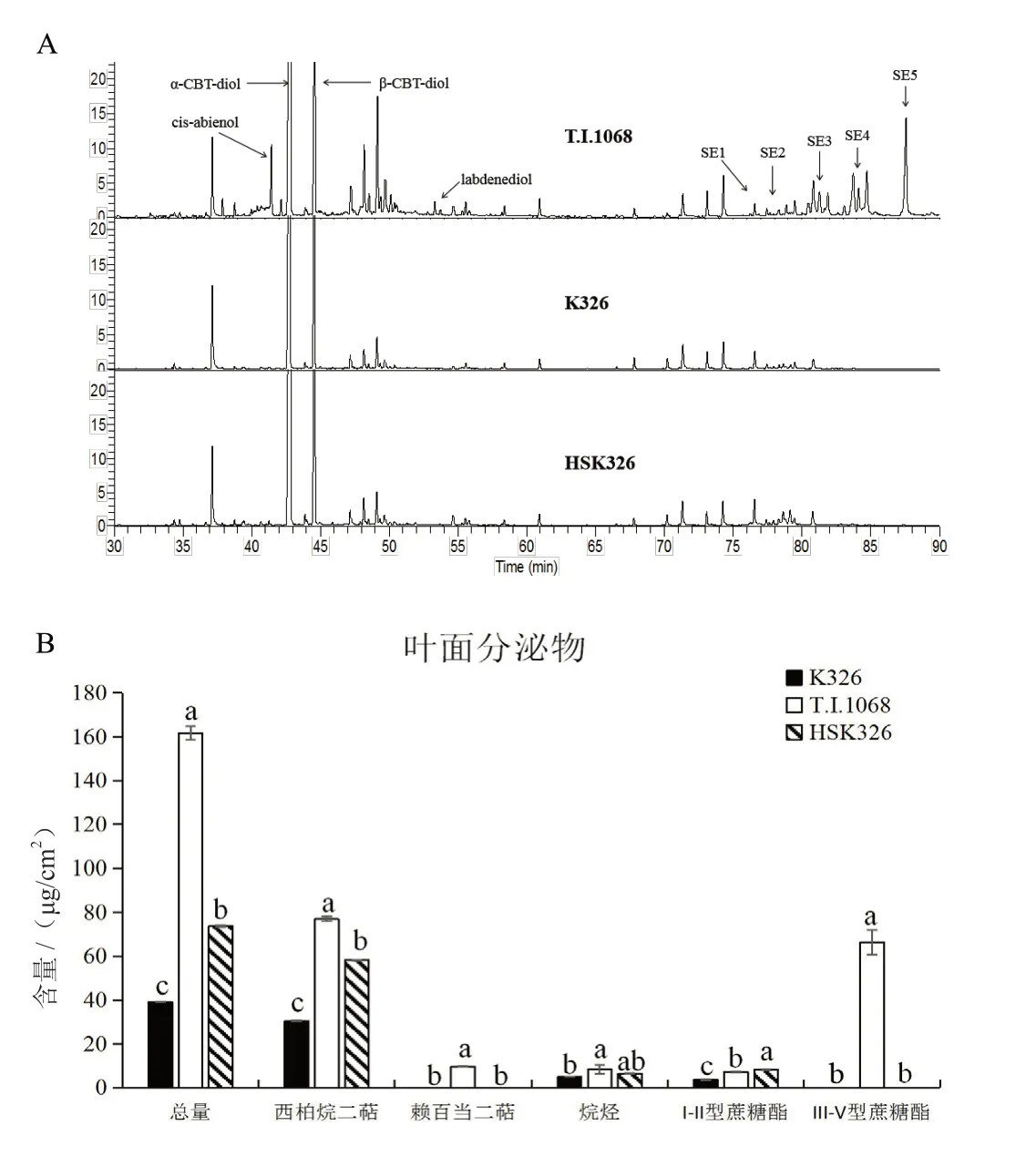
图4 叶面分泌物检测分析Fig.4 Chemical Analysis of leaf surface chemicals
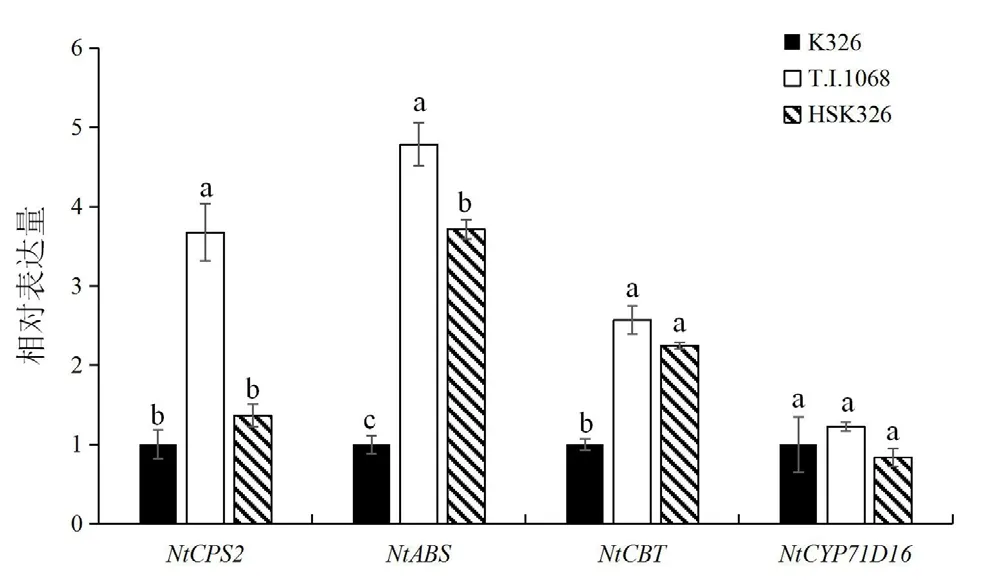
图5 二萜合成基因表达水平分析Fig.5 Analysis of the expression level of diterpene synthesis genes
根据Sallaud等人的研究结果[16],NtCPS2基因第4外显子中存在一个G/T的SNP,引起翻译提前终止,是造成大部分烤烟中赖百当二萜合成缺陷的主要原因。我们据此设计引物,分别对HSK326、K326及T.I.1068中NtCPS2基因目标序列进行PCR扩增和序列测定,结果如图6所示。可以看出,与T.I.1068相比,HSK326和K326在1542 bp处都发生了T碱基突变,这可能是二者赖百当二萜缺失的主要原因。

图6 HSK326、K326及T.I.1068 NtCPS2基因及蛋白序列比对分析Fig.6 NtCPS2 gene and protein sequence alignment analysis of HSK326, K326 and T.I.1068
2.3.2 III-VI型蔗糖酯合成连锁分子标记分析
BMVSE基因与烟草III-VI型蔗糖酯的生物合成密切相关[17],但至今该基因的序列和功能还不清楚。Vontimitta等[6]对BMVSE基因进行了遗传定位,并开发了一系列与BMVSE连锁的SSR分子标记。其中PT20315和PT30354两个标记多态性稳定,且与糖酯类型高度一致[18]。因此,我们选用这两个标记对HSK326、K326及T.I.1068进行了基因型鉴定,结果如图7所示。可以看出,PT20315和PT30354两个标记可以把亲本K326和T.I.1068区分开来,而HSK326的带型均与K326相同,说明HSK326与K326中缺少III-VI型蔗糖酯合成的关键基因。这与蔗糖酯的检测结果相一致。
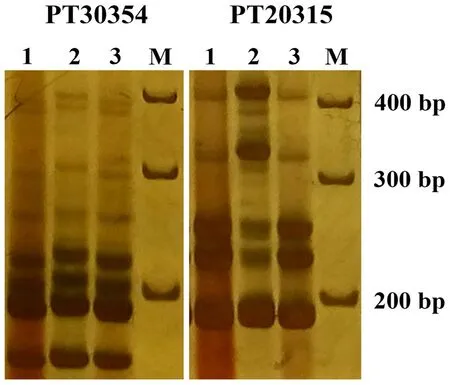
图7 HSK326、K326及T.I.1068中III-VI型糖酯合成基因分子标记鉴定Fig.7 Molecular marker identification of type III-VI sugar ester synthesis genes in HSK326, K326 and T.I.1068
3 讨论
烟草腺毛作为表皮细胞的特化结构,不仅是烟株与环境间的物理屏障,其分泌物更是天然的化学防线,与烟株抗性和烟叶品质密切相关[19]。叶面分泌物含量的高低作为品种的遗传特性,由腺毛密度、腺毛类型、腺毛分泌物合成及分泌能力等因素所共同决定。烤烟腺毛为混合型,分泌型与非分泌型共存。腺毛密度,尤其是长柄分泌型腺毛的密度普遍较低,分泌能力不高,腺头分泌囊较小,导致了其叶面腺毛分泌物含量在不同类型烟草中处于较低水平[12]。由于烟草腺毛发育及物质代谢的分子调控机制并不十分清楚,采用常规选育技术,将种质资源中的高分泌特性导入栽培品种,仍是目前行之有效的途径之一。T.I.1068作为典型的高分泌型烟草资源,具有腺毛密度高、分泌旺盛、成分齐全等特点,是烟草叶面分泌物研究的典型材料[10]。本研究将T.I.1068作为供体亲本与K326进行杂交,经过多代回交后,将其高分泌特性成功导入K326,获得了高分泌型K326改良品系HSK326。
HSK326的一个明显特征是腺毛密度较高。虽然,HSK326与K326的腺毛类型一致,但分泌型腺毛,包括长柄分泌型和短柄分泌型的密度,比K326都有明显增加。T.I.1068高腺毛密度的分子机制并不清楚。已有的研究证明,NtCycB2基因的突变会提高长柄分泌型腺毛的密度[8],但T.I.1068中该基因的序列和表达水平与K326相比,并无明显差异(未发表)。从T.I.1068中非分泌型腺毛缺失,推测其控制非分泌型腺毛发生的基因通路受阻,这或许在某种程度上促进了分泌型腺毛的发生。HSK326在分泌型腺毛密度提高的同时,非分泌型腺密度有所降低,预示烟草分泌型腺毛与非分泌型腺毛发生之间存在某种交叉调控机制。
腺毛分泌能力强是HSK326的另一个特征。形态观察可以看出,HSK326腺头分泌囊比K326明显增大,罗丹明着色程度明显加深,说明其腺毛分泌物,尤其是糖酯的合成和分泌更为旺盛。GC/MS检测结果也表明,HSK326腺毛分泌物总量虽然低于T.I.1068,但比K326提高了接近一倍,这与二萜合成关键基因的表达水平普遍提高相一致。腺毛密度的增加可能是导致基因表达水平提高、分泌物含量增加的一个原因。然而HSK326的腺毛密度与K326相比仅提高了38%,但西柏烷二萜含量比K326提高了91%,蔗糖酯含量提高了127%,此外HSK326还表现出腺毛分泌囊增大、着色加深的表型,说明单个腺毛中物质合成和分泌能力的增强也促进了腺毛分泌物总量的提高。
采用常规育种手段对腺毛分泌物含量进行遗传改良的一个难点是表型鉴定。前人研究表明烟草腺毛分泌物被腺头细胞分泌出来后会储存在腺头部分,形成一个分泌囊[20],因此分泌囊的大小反应了腺毛分泌能力及腺毛分泌物的含量,利用罗丹明B对腺毛染色能够较为准确的对分泌物中的蔗糖酯进行定量[21],染色越深,糖酯含量越高。在HSK326选育过程中我们采用了比较腺毛分泌囊的大小结合化学染色的方法,有效解决了表型鉴定困难的问题。利用这两种观察方法在苗期就可以完成表型观察及候选单株的确定,从而省去了大群体的田间种植及分泌物提取检测的过程,有效提高了烟草腺毛分泌能力品种选育的效率,对于烟草高香气育种工作具有重要的参考意义。
从分泌物组成成分来看,HSK326与K326的二萜组成一致,都只含有西柏烷化合物,缺少冷杉醇、赖百当二醇等赖百当化合物。虽然,赖百当二萜合成途径关键基因NtCPS2在HSK326与K326中都能正常表达,但序列测定发现T.I.1068和K326在NtCPS2基因1542 bp处存在一个G/T的SNP,K326中的T碱基能引起翻译提前终止,造成腺毛分泌物中不含有赖百当二萜。HSK326中对应位置与K326相同,也为T碱基,使HSK326获得了K326中赖百当化合物缺失的特性。赖百当类物质会影响烤烟的香气风格,例如含有冷杉醇的“豫烟11”[11]、“大白筋”[22]都具有明显的晾晒烟风味。因此赖百当类物质的缺失对HSK326保持烤烟的香气风格至关重要。另外,HSK326与K326都只含有I-II型蔗糖酯,缺少III-VI型蔗糖酯。据Vontimitta等人报道[6],III-VI型蔗糖酯合成基因BMVSE与冷杉醇合成基因NtCPS2呈现紧密连锁状态。因而,III-VI型蔗糖酯与冷杉醇常常共存于香料烟、雪茄等晾晒烟中,而在烤烟、白肋烟中则不含这两类物质。蔗糖酯对卷烟具有增香保润、增加甜感、改善吃味的作用[23],至于III-VI型蔗糖酯对烤烟风格的影响还未见有报道。我们在BC5F2群体的筛选中,也发现了打破BMVSE与NtCPS2连锁的材料,不含冷杉醇但含有III-VI型蔗糖酯。对该材料的进一步分析,有望阐明蔗糖酯不同组分对烤烟风格的影响。
4 结论
本研究将K326与T.I.1068进行杂交,以K326为轮回亲本进行连续回交,最终筛选获得高分泌型K326近等系HSK326。HSK326在保持与K326植物学性状基本一致的同时,叶面腺毛密度增加、分泌能力增强、分泌物含量显著提高;而且,腺毛分泌物组分与K326高度一致,保持典型的烤烟叶面化学特征,具有较大的应用潜力。
