甲基营养型大肠杆菌构建策略的研究进展
2021-07-28陶雨萱张尚杰景艺文信丰学董维亮周杰蒋羽佳章文明姜岷
陶雨萱,张尚杰,景艺文,信丰学,2,董维亮,2,周杰,2,蒋羽佳,章文明,2,姜岷,2
(1 南京工业大学生物与制药工程学院,材料化学工程国家重点实验室,江苏南京 211816;2 南京工业大学,江苏先进生物与化学制造协同创新中心(SICAM),江苏南京 211816)
利用微生物发酵生产生物基产品符合“既要金山银山,也要绿水青山”的可持续发展理念。近年来,生物制造产业蓬勃发展,已成为我国有经济竞争性的新型战略产业,前景光明。但是传统的微生物发酵通常以谷物等农作物为原料,不仅原料成本高昂,而且面临着“与民争粮”、“与粮争地”的争议或技术经济性问题。因此,寻找可再生、低成本的生产原料迫在眉睫。木质纤维素曾被认为是有效的替代品,但是木质纤维素的预处理成本高、水解抑制物去除难、发酵产物的产量低,制约了其进一步发展。作为化工业的重要产品,甲醇、甲烷、甲酸等一碳原料具有储量丰富、来源广泛、价格低廉等优点,深具改变传统生物发酵生产模式的潜力[1]。此外,甲醇的还原度高于木质纤维素糖(葡萄糖、木糖等),其代谢过程中可以产生大量的还原力,是微生物生产还原性高附加值产品的优良底物之一,对实现谷物原料向“非粮”的转移目标具有重要意义。
自然界中天然代谢碳一化合物的微生物包括真核和原核类的甲基营养菌。这些天然甲基营养菌能以甲醇等碳一化合物为唯一碳源,即通过同化代谢途径将碳一化合物转化为生物质,并通过异化代谢途径从中获得能量。甲醇的同化代谢途径有核酮糖单磷酸循环途径(RuMP cycle)、丝氨酸循环途径(serine cycle)和木酮糖单磷酸循环途径(XuMP cycle)3 种,后者仅存在于真核甲基营养菌中[2]。目前已在天然甲基营养菌中实现有机碳一原料到氨基酸、甲羟戊酸、正丁醇、戊二胺以及单细胞蛋白等多种化学品的生产(见图1)。早在1986 年,研究者就以甲醇为底物,利用扭脱甲基杆菌(M.extorquensAM1)生产丝氨酸[3]。在2000 年,研究者们利用毕赤酵母(Pichia pastoris)、汉逊酵母(Hansenula polymorpha)等甲基营养酵母菌,以甲醇为底物生产单细胞蛋白用于动物饲料[4]。Liang等[5]通过在甲基营养菌M.extorquensAM1 中引入甲羟戊酸模块,并通过代谢模块优化、培养基优化和转录调节工程,最终的构建菌株可合成2.67g/L 的甲羟戊酸。Zhang 等[6]通过在毕赤酵母中引入番茄红素的合成模块,结合启动子工程调控策略,使其番茄红素产量可达0.714g/L,为目前报道的毕赤酵母最高水平。然而,由于大多数天然甲基营养菌的生长速率慢、异化代谢能力强,导致代谢物的生产效率低,并且缺乏有效的遗传操作工具,限制了其在化学品生产方面的潜力[7]。
近年来,研究者致力于利用代谢工程、合成生物学以及进化代谢等技术手段,将大肠杆菌(Escherichia coli)、谷氨酸棒状杆菌(Corynebacterium glutamicum)和酿酒酵母(Saccharomyces cerevisiae)等模式微生物改造为人工甲基营养菌,进而实现甲醇到各种化学品的生物合成。其改造策略的本质是将天然甲基营养菌中各种甲醇同化代谢途径(包括核酮糖单磷酸途径、丝氨酸循环途径等)的关键催化元件和模块整合到上述模式微生物中。本文介绍了天然甲基营养菌的多种代谢路径,综述了引入天然甲醇代谢路径构建甲基营养型大肠杆菌的多种方法,总结归纳了不同代谢路径的优化手段与改造成果,并对近年来基于计算机辅助设计合成的人工路径进行了介绍,进而提出对合成路径适配性改造和工程菌株耐受性提高的展望。
1 基于核酮糖单磷酸循环的甲基营养型大肠杆菌构建策略
甲基营养菌代谢甲醇的第一步是将甲醇氧化为甲醛。根据电子受体的类型,可将该酶分为依赖吡咯喹啉醌(PQQ) 的甲醇脱氢酶(methanol dehydrogenase,MDH)、依赖烟酰胺腺嘌呤二核苷酸(NAD) 的甲醇脱氢酶和依赖O2的甲醇氧化酶(alcohol oxidase,AOX)3类[8]。随后甲醛进入3种不同的同化路径,即核酮糖单磷酸循环、丝氨酸循环和木酮糖单磷酸循环。基于生物量、能量和辅因子的产生能力分析发现,依赖NAD 的甲醇脱氢酶和磷酸核酮糖途径(RuMP)被认为是在模式菌株中组装甲醇代谢途径的最优选择[9]。
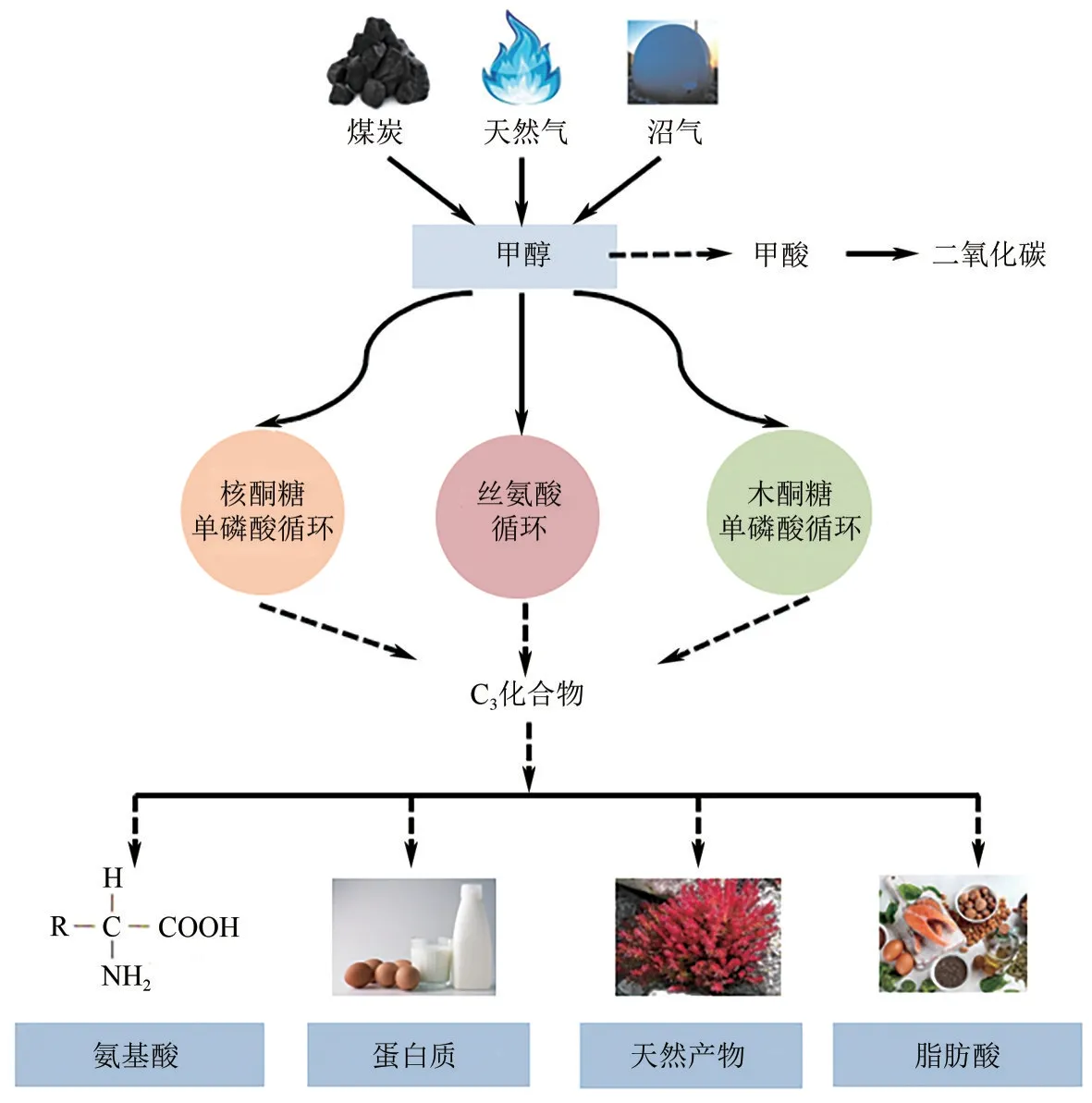
图1 以甲醇为原料生产各种化学品示意图
RuMP 途径是目前甲醛同化效率最高的途径,可分为3个部分——氧化部分、裂解部分和重排部分[9]。氧化部分,甲醛与5-磷酸核酮糖(Ru5P)缩合并异构化生成关键产物6-磷酸果糖(F6P),该反应由两个关键酶3-己酮糖-6-磷酸合成酶(3-hexulose-6-phosphate synthase,HPS)和6-磷酸-3-己糖异构酶(6-phospho-3-hexuloisomerase, PHI)催化[10]。F6P 在细胞内一般通过糖酵解途径(EMP)、Entner-Doudoroff 途径(ED)和磷酸戊糖途径(PPP)进行代谢。随后F6P被裂解成3-磷酸甘油醛(GAP)和二羟丙酮磷酸(DHAP),最后GAP 和景天庚酮糖-7-磷酸(S7P)被转酮醇酶(transketolase,TK)重排为两个五碳糖磷酸酯,后者进一步转换为Ru5P(图2)。相关研究表明,Ru5P的再生是维持循环运行的关键[20]。
1.1 优化关键酶的催化能力
Müller 等[11]最先在工程菌株中实现甲醇异化和同化,并利用合成生物学的方法将来源于甲醇芽孢杆菌(Bacillus Methanolicus)的甲醇代谢模块成功引入大肠杆菌。研究者们认为实现甲醇利用的最有效方法是组装NAD 依赖的甲醇脱氢酶以及基于HPS-PHI 的甲醛同化模块,通过在大肠杆菌中表达甲醇芽孢杆菌(B.Methanolicus)的MDH2、HPS和PHI 基因,Müller 等[11]构建了RuMP 循环,使甲醇在中心代谢中的通量达到了40%。Whitaker 等[12]根据大肠杆菌的最适生长温度,筛选出嗜热脂肪芽孢杆菌(Bacillus Stearthermophilus)的MDH与来自甲醇芽孢杆菌的HPS和PHI,并引入工程大肠杆菌中,大幅提升了甲醛消耗量,且其构建后的工程大肠杆菌菌株MDH 活性与天然甲基化营养菌的活性相似。
以往对甲基营养型大肠杆菌进行改造的研究中,大多集中在酶方向,其改造效果受到甲醇脱氢酶动力学不利因素的影响。Wu 等[13]提出甲醇芽孢杆菌的MDH依赖于内源激活蛋白(ACT),并且对甲醇表现出较低的酶特异性。研究者们从Cupriavidus necatorN-1 分离纯化出对甲醇亲和力高、中温条件下无需活化的MDH2,以此为基础构建了MDH2的变异体(CT4-1),使Kcat/Km值(酶的二级常数)提升了6倍,大幅提升了催化甲醇的效率[13]。针对依赖NAD 的MDH 中温条件下热力学特性较差的问题,Price 等[14]发明了超分子酶组装方法,利用SH3-配体相互作用将MDH、HPS 和PHI组装成工程超分子酶复合物,并在大肠杆菌中进行酶组装,将甲醇消耗率提高了9倍,为工程菌株的MDH酶活改善开辟了新的方向。

图2 基于RuMP路径的优化改造[11-19]
1.2 增强Ru5P再生与利用率
在天然RuMP 路径中,Ru5P 不足是微生物无法拥有高甲醇通量的主要制约条件。Ru5P 耗尽一方面导致甲醛的积累,细胞无法耐受高浓度的甲醛毒性,另一方面无法维持循环,限制了甲醇的持续利用。因此,提升Ru5P 的再生能力是构建工程菌株的关键问题。目前解决此问题有多种研究策略,一种是抑制F6P 进入糖酵解途径以提升F6P 的含量,进而增强Ru5P 的再生,另一种是引入甲基营养菌的优势转化基因以驱动大量Ru5P的生成。
在底物方面,工程菌株不能像天然甲基营养菌那样以甲醇为唯一碳源进行代谢,为了提高甲醇的掺入量以增加甲醇利用率,通常加入碳源或氨基酸作为共底物。Gonzalez等[15]评估了25种不同的共基质对甲醇的同化作用,比较了多种氨基酸底物对甲醇利用的影响,发现苏氨酸的利用可提高甲醇利用率,并且其调节机制受亮氨酸反馈调节蛋白(LRP)的影响。通过敲除LRP,工程菌株在甲醇中的生物量比对照组高出34%,间接改善了甲醇的同化效率[15]。Meyer 等[16]选用葡萄糖酸盐和甲醇共底物以提供5-磷酸戊糖,进一步提高中间体5-磷酸核酮糖的利用率,使高达24%的甲醇进入中心代谢途径。He 等[17]通过氧化肌氨酸生产甲醛,外源添加木糖生成5-磷酸木糖,进一步合成5-磷酸核酮糖以避开再生循环,增强了Ru5P 的直接利用率。
在代谢通量方面,Bennet等[18]通过敲除磷酸葡萄糖异构酶基因(PGI),重建了G6P 的代谢途径,在一定程度上降低了糖酵解的代谢通量,改善了甲醇和葡萄糖的共同利用。研究者们通过引入甲醇芽孢杆菌的5 种酶基因,即核酮糖磷酸异构酶(RRE)、果糖-二磷酸醛缩酶(FBA)、磷酸果糖激酶(PEK)、转酮醇酶(TKT)和景天庚酮糖二磷酸酶(GLPX),强化了大肠杆菌F6P 到Ru5P 的转化,提升了甲醇的同化能力,使PGI缺陷菌株的丙酮酸效价提升了110%[18]。Woolston等[19]使用抑制剂碘乙酸降低糖酵解通量,并在大肠杆菌中过表达GLPX,激活了RuMP 的景天庚酮糖磷酸酶(SBPase)变体,以提升S7P 的合成量进一步提升Ru5P 的再生能力(图2),维持循环的同时降低了细胞内的甲醛浓度,并使甲醇通量增加了2倍。
这些实验结果从底物、酶、循环维持等多个方向着手,在大肠杆菌中优化引入RuMP途径的实践中有了突破性的进展(见表1),优点是大幅度提高了甲醇氧化的代谢通量,缺点是工程菌株始终无法直接高效地利用甲醇。而在2020 年,Chen 等[21]通过基因编辑技术与拷贝数变异(CNVs)缓解了大肠杆菌在甲醇中的生长障碍——DNA-蛋白质交联(DPC),并通过适应性进化使菌株突变,平衡了代谢通量,最终实现了以甲醇为唯一碳源生长;其合成甲基营养型大肠杆菌的倍增时间为8.5h,甲醇耐受性可达1.2mol/L。

表1 基于RuMP路径的大肠杆菌构建策略
2 基于丝氨酸循环的甲醇代谢路径构建与优化
在丝氨酸途径中,甲醛和四氢叶酸(H4F)可自发转化为亚甲基四氢叶酸;也可先转化为甲酸盐,再通过H4F依赖型酶转化为亚甲基四氢叶酸。丝氨酸羟甲基转移酶(serine hydroxymethyltransferase,SHMT)催化甲醛与甘氨酸的加成,从而生成丝氨酸[24]。循环的碳同化部分包含3个部分,涉及碳二(C2)、碳三(C3)和碳四(C4)代谢物。碳二代谢物包括乙醛酸、甘氨酸和输出的乙酰辅酶A。在碳三化合物生成部分,SHMT 催化甘氨酸和5,10-亚甲基四氢叶酸反应生成丝氨酸(C3),丝氨酸进一步转化为磷酸烯醇式丙酮酸(PEP)(C3)。随后PEP 经羧化酶羧化后生成碳四化合物草酰乙酸(OAA),最后草酰乙酸转化成两个碳二化合物并产生乙酰辅酶A[图3(a)][24]。
Methylobacterium extorquensAM1 是一种兼性甲基营养细菌,也是研究甲基营养菌生物化学和遗传学特性的模式菌株。Kalyuzhnaya等[25]从该菌株中发现了QSCR 能够调节丝氨酸循环中关键酶的活性,并影响丝氨酸循环基因的转录,实验证明其与编码大部分丝氨酸循环酶的两个操纵子的启动子(QSC1 操纵子的SGA-HPR-mtDA-FCH 和QSC2 操纵子的mtkA-mtkB-PPC-MCLA)相互作用。后续实验预测了QSCR 的基因结合位点,表明H4F参与的循环模块受到QSCR的激活调节,并证明了甲酰四氢叶酸的积累能直接激活QSCR以结合调节的启动子,使丝氨酸C1转移途径基因的转录增加,进一步提高了转化率[26],为后续构建大肠杆菌的丝氨酸代谢奠定了基础。
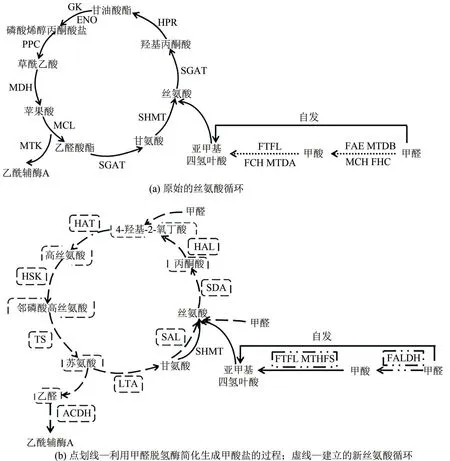
图3 基于丝氨酸循环的优化改造[24,26-28]
Yu 等[24]因大肠杆菌难以直接利用甲醇,对AM1 中的天然丝氨酸循环进行了修改。一方面为缩短甲醇的氧化流程,研究者利用依赖NAD 甲醛脱氢酶(FALDH)来简化甲醛氧化成甲酸盐的过程,并将甲酸盐转化为亚甲基四氢叶酸以提供碳一载体;另一方面,丙氨酸转氨酶(AGT)催化乙醛酸转氨生成甘氨酸,取代了天然途径的氨基供体丝氨酸,将甘氨酸与上述亚甲基四氢叶酸合成完成修饰途径丝氨酸的生成[图3(b)]。为了避免天然丝氨酸循环中有毒中间体羟基丙酮酸的形成,减少对中心代谢通量的干扰,Leander 等[27]通过引入天然苏氨酸裂解酶,使大肠杆菌中整个细胞的甘氨酸需求由苏氨酸的生物合成和降解来提供,构建了丝氨酸-苏氨酸循环,为实施甲酸在大肠杆菌中代谢提供了可行的方向。
为最大限度地提高生物质量,提升热力学优势,He 等[28]进一步优化了丝氨酸循环以提高大肠杆菌对甲醛的利用率。通过丝氨酸醛缩酶(serine aldolase, SAL)将甲醛与甘氨酸缩合生成丝氨酸,再由丝氨酸脱氨酶(serine deaminase, SDA)脱氨为丙酮酸,此过程取代了原有复杂、多ATP 消耗的甲醛同化路径;随后由4-羟基-2-氧丁酸(HOB)醛缩酶催化生新C4中间体HOB,氨化生成丝氨酸。此路径以高效简洁的甲醛同化反应取代羧化反应,所需酶数量(8种)仅为天然路径的1/2,在节省能量、提升循环效率、依赖性小、符合热力学驱动力等多方面具有绝对的优势。
虽然丝氨酸循环的同化效率不如RuMP 路径,但是引入丝氨酸循环能够从产物乙酰辅酶A衍生更多的高价值产物,如乙醇、丙酮、丁酸等。此循环更有利于合成多碳化合物,对于衍生物的开发也更具有研究性。此外,与天然好氧的甲醇同化途径相比,丝氨酸循环不依赖氧气,是非常有价值有竞争力的甲基化工程菌株构建路径(见表2)。但是天然丝氨酸循环由于存在中心代谢反应较多、循环ATP低效等问题,在热力学、动力学、同化效率等多个方面,仍然存在着许多缺点需要攻克。

表2 基于丝氨酸循环的大肠杆菌构建策略
3 基于计算机辅助设计的人工合成路径
尽管在缺乏一碳合成代谢途径的生物中(如大肠杆菌、酿酒酵母)构建天然甲基营养型菌株的代谢路径已经取得了许多突破,但天然甲醇代谢路径存在的固有缺陷无法完全克服,这些问题包括路径复杂性、环境敏感性、氧气依赖性、同化低效性、动力学热力学劣势等。在合成生物学中利用碳一原料的最理想途径应该具有如下特点:①依赖的酶数量少;②反应步骤短,并且与其他代谢途径分离,避免中心代谢流通量的冲突;③驱动力强;④在好氧和厌氧条件下都能高效地发挥功能[27];⑤能快速代谢有毒中间产物等。研究者们以此为目标,设计了比天然路径更为合理的人工一碳利用路径(图4)。

图4 人工合成甲醇代谢路径[29-32]
Siegel 等[33]利用计算机设计了一种甲醛酶(FLS),是设计催化酶活性的全新方法,它能够实现碳一到碳三化合物的合成,可以催化3个甲醛分子合成一个二羟基丙酮分子。以此酶可以构建一种新的甲醛酶途径,使甲醛转化为中心代谢物二羟丙酮磷酸(DHAP)。使用计算机辅助蛋白质设计开辟了新的酶设计领域,适用于在体外构建固定碳的人工途径。
为简化代谢步骤,Wang 等[29]在大肠杆菌内高效表达了甲醇芽孢杆菌依赖NAD+的MDH和一种计算机设计的甲醛酶(FLS),组装了人工线性甲醇同化途径。此线性途径不需要甲醛受体,由两个酶主导完成反应:先利用甲醇脱氢酶将甲醇氧化成甲醛,再利用甲醛酶将甲醛碳化成二羟丙酮[29]。以上述最优途径法则来评估,此线性途径虽在热力学上不如天然RuMP途径有利,但能直接生产C3中间体DHA,可以快速进入糖酵解并完成代谢以提供强大的驱动力,在工程大肠杆菌进一步提高甲醇利用率的探索中实现了全新的构想。
乙酰辅酶A作为乙酰基的唯一供体,是许多工业化学品C2基团的供体。前文有提到利用改进的丝氨酸循环还将甲醇和碳酸氢盐生产乙酰辅酶A,但Lu 等[30]设计了一条全新的、不与已知代谢途径重叠的合成乙酰辅酶A(SACA)的途径。SACA途径对乙醇醛合成酶和乙酰磷酸合成酶进行了改造,将乙醇醛合成酶的催化活性提高了70 倍以上。通过此途径,可以实现由甲醛等多种碳一生物合成乙酰辅酶A[30]。SACA 途径具有合成途径短、热力学化学驱动力强劲、不依赖氧气等多种优势,为将来使用多种碳一底物生产乙酰辅酶A及其衍生物创造了可能性。
Yang 等[31]利用COMB-FBA 的组合算法筛选出最合适的苯甲酰甲酸酯脱羧酶(Gals)和转醛缩酶(TalBF178Y)两种工程酶进行人工反应,通过新的组合计算方法,从选择酶和改进工艺方面入手,研究者们在体外构建了一条新的C1同化途径即乙醇醛同化(GAA)途径,使其碳转化率达到了88%。
碳一人工合成路径的创建目标明确,在已知天然路径缺陷的基础上,设计全新的简化路径以提升合成效率,往往以最适用于工程菌株、最简单化代谢路径、最符合动力学驱动力等为标准,以计算机以及各种新型算法为辅助筛选最适用反应的酶以大幅提升酶活性,将在甲醛代谢通量方面取得较大的突破(见表3)。

表3 人工合成甲醇代谢路径的构建策略
4 结语及展望
全球气候变化引起了人们对当下能源结构的担忧,大量化石燃料的使用产生的温室气体严重破坏了环境。在环境友好的同时保证能源的供应,依靠微生物利用可再生资源合成生物燃料是一个重要的解决方案。
生物制造业迫切需要改善微生物中原有碳同化途径,然后引导这些途径的代谢流通过各种自然或设计的途径高效地生产目标产物。就微生物发酵底物而言,碳一化合物以其丰富的来源而颇具优势。但以甲醇为例,虽然其中一些化合物或中间体可以从天然的甲基营养菌的各种代谢途径中产生,但这些天然代谢途径往往需要优化或重新设计,将繁琐的长路径简化并提高甲醇的同化效率。本文主要综述了将天然甲醇代谢路径引入大肠杆菌的研究进展,并概述了与之相关的一些人工合成路径。
针对菌株的耐受性问题,可从以下3个方面考虑。第一,提升菌株的甲醇耐受性。一方面对天然甲基营养型菌株的甲醇耐受性进行比较,挑选出耐受性较好的菌株分析其基因组信息,获得潜在的耐受元件并导入工程菌株中验证;另一方面,采用等离子体、常压室温等离子体(ARTP)等诱变技术,筛选出甲醇耐受性强的菌株。第二,为减少甲醛对细胞的毒性,可强化已有代谢酶的表达,提升甲醛的代谢效率;也可外源引入甲醛代谢途径,多途径提升甲醛的代谢通量;亦可通过人工合成酶简化甲醛原有的代谢路径。第三,通过工程菌进化以进一步适配天然代谢路径,比如可采用适应性实验室进化(ALE),通过提高细胞对甲醇的耐受性来提高合成的甲醇营养菌的甲醇生物转化率。原始的天然合成路径需要从各个方面不断优化,如酶活性、代谢通量、动力学优势等,从底物优化、代谢路径简化、酶活提升等各个方面强化甲醇的利用。最后,随着计算机算法的先进与科研的不断发展,人工合成的新路径是一个突出的研究方向,其目的性明确、筛选精准、对于特定产物有针对性的产量强化,在未来的工业应用方面具有很强的可塑性,可将高效的人工合成酶与耐受性强的工程菌株相结合,引入优化的甲醇代谢路径,从多方面着手,突破现有的甲基工程菌株的构建瓶颈。
