重庆綦江早奥陶世特马豆克阶藻类增长及其指示意义
2021-07-27张虹瑞穆财能刘欣春杨丽君范程华
张虹瑞, 刘 滔, 穆财能, 刘欣春, 杨丽君, 范程华
(1. 油气藏地质及开发工程国家重点实验室(成都理工大学),成都 610059;2.彝良驰宏矿业有限公司,云南 昭通 657600;3.中海石油(中国)有限公司 天津分公司,天津 300450)
奥陶纪生物大辐射从奥陶纪早期特马豆克阶一直贯穿到奥陶纪晚期凯迪阶,即485.4~444 Ma B.P.,历经41.4 Ma,是地质历史时期最大的一次生物辐射事件,其后2亿年时间里海洋生物总体多样性基本不变[1-3]。并且生物辐射最直观的表现就是在科、属、种等较低级别的分类单元上成倍增加(如华南上扬子地区中华正形贝动物群的大量繁盛 )[4]。生物活动的范围也从原来的陆棚浅海区不断向深水区域发展,达到了前所未有的繁盛景象,如华南、塔里木等地区的上奥陶统宝塔组和坎岭组的混积陆棚相沉积中有大量的震旦角石,在五峰组和印干组黑色页岩中有大量的笔石化石,为较深水沉积[5]。随之而来的是,由于生物活动区域的增大,导致生物地理分区性不断增强,如华南上扬子地区奥陶纪弗洛期早期出现的一系列较深水沉积的正形贝类,经过不断地向上扬子台地拓展生态空间,在台地中心多个地点形成了新的底栖生态群落[6-7]。
如图1所示,华南奥陶纪腕足动物的生物大辐射出现了多个多样性急速增长值(峰值),大部分腕足动物的生物类群急速增长峰为3个,分别出现在早奥陶世弗洛期、中奥陶世达瑞威尔期早中期和晚奥陶世凯迪期晚期。尽管全球不同地区海洋无脊椎动物生物类群出现的峰值时间存在一些差异,但通过大量资料研究对比,这三个峰值较为符合全球生物辐射规律[7-10]。并且生物辐射的规模有逐渐递增趋势,凯迪晚期规模最大,实现了对寒武纪澄江生物群的全面替代,如腕足动物群和苔藓动物群对三叶虫I-bex动物群和磷酸盐质壳腕足动物群的替代,从此海洋无脊椎动物的基本演化框架得以建立[4]。
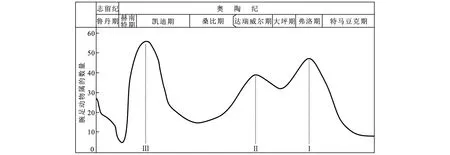
图1 华南奥陶纪腕足动物的生物辐射曲线Fig.1 Biological radiation curves of Ordovician brachiopods from South China(修改自文献[7])
本文在前人研究的基础上,对奥陶纪生物辐射事件的触发机制进行研究。奥陶纪初期,全球及区域构造运动加剧,导致Gondwana超级大陆逐渐解体,华南、阿根廷、南欧、伊朗等[7]多个地体逐渐向赤道附近聚集,从而导致了地理隔离的形成,为该时期生物辐射提供了足够的推动力。起始于晚寒武世,并在奥陶纪持续进行的海洋“底质革命”[1],以及混积陆棚海沉积区域在早-中奥陶世达到了显生宙以来的最大规模,进一步扩大了滤食生物的生存场所,为其辐射演化提供了必要的先决条件。在此期间,全球古海洋温度也在30~40 ℃波动[5],如此温暖的气候有利于底栖生物(如双壳类、珊瑚、三叶虫等)在奥陶纪的持续繁盛。与此同时,大气中氧气含量上升[11],海平面也达到了古生代的最高水平[12],加上海洋环境的流通[13]和深层陆架以及斜坡区域的渐进氧化[14-15],包括陆源物质的输送[16]等因素的协同作用,为生物生存、繁殖和壮大创造了绝佳的环境条件。
对于早奥陶世生物辐射事件的研究,更多的着眼于生物量演化及初步启动机制等方面,目前所获得的共识是奥陶纪早期海洋氧含量及营养物质的增加导致了生物辐射事件的发生,但其中究竟是何种因素引发了氧含量的增加尚需进一步研究探讨。K.Pehr等[17]的研究结果表明,藻类可以作为海洋宏体生物的食物,而且藻类能吸收无机的氮和磷等物质并大量释放氧气,促进生物圈其他圈层的壮大。作为连接宏体生物和微生物的纽带,藻类在生物辐射过程中的影响不可忽视,而其与宏体生物间及大洋氧气演化的协变关系也极具研究价值。因此,本文利用薄片观察及生物量统计等手段,对重庆綦江观音桥剖面早奥陶世特马豆克阶和弗洛阶进行研究,重点探讨藻类变化与大洋氧含量演化的相对关系。
1 剖面介绍
本文研究剖面位于重庆市綦江区安稳镇的观音桥村附近(图2),在地质上为四川盆地东南边缘,分布于黔中隆起北侧和雪峰山隆起西侧构造带之间的过渡带上。张道光[18]指出,奥陶纪该地区为被动大陆边缘的海相沉积,受全球加里东运动影响,上扬子地台大部分抬升成陆,开始遭受风化剥蚀,导致该区大面积的区域缺失泥盆系、石炭系等地层。该区域于二叠纪再一次沉降开始沉积,而到了中三叠世主要发育海相碳酸盐沉积,印支运动晚期以后该区域演变为陆内湖盆沉积。研究区在构造上属于上扬子板块(图2),奥陶纪-志留纪时期上扬子地台经历了多期构造作用[19-20],特别是中奥陶世后受南东方向的华南板块向西北方向扬子板块的挤压增强,除扬子北缘仍属于被动大陆边缘外[21-22],其他地区均表现为构造挤压隆升的背景。此时许多古隆起,如川中隆起、黔中隆起和雪峰山隆起的迅速扩大导致上扬子地区的海平面上升。加里东运动的众多隆起起到了阻隔海水的作用,盆地底部的山地和丘陵也使得水体和大洋无法进行有效的交流,这种情况在奥陶世晚期变得更为明显[23]。
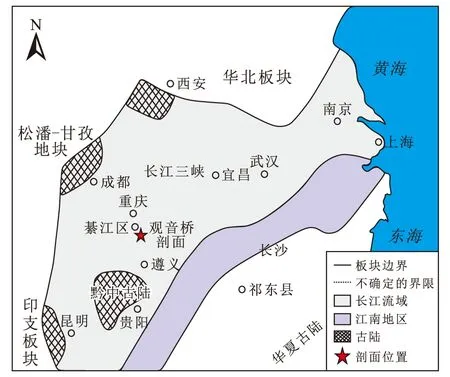
图2 上扬子地区大地构造背景Fig.2 Tectonic setting of the upper Yangtze Region(修改自文献[20 ])
观音桥剖面北距綦江县90 km,渝黔高速与G210国道交叉处[24]。剖面起点为中上寒武统娄山关群与下奥陶统桐梓组的分界点,坐标为28°37′12.36″N、106°47′10.68″E;剖面终点为上奥陶统五峰组观音桥段与下志留统龙马溪组的分界点,坐标为28°37′51.90″N、106°47′9.50″E。
剖面整体构造简单,出露较好,层序清楚,生物化石丰富。剖面起点由于工程施工,娄山关群被破坏不可见。对于奥陶纪生物事件与藻类变化的耦合关系这一研究,样品采集包括后续的生物量统计都是特别重要的工作。样品采集的密度、数量以及新鲜度等都会直接影响到生物量统计工作,进而会对我们判断奥陶纪生物事件与藻类的耦合关系时造成影响。因此,野外采集样品时,全部选择新鲜面敲取样品。根据露头情况,在桐梓组、红花园组和湄潭组,大致为1~2 m的采样密度(图3)。
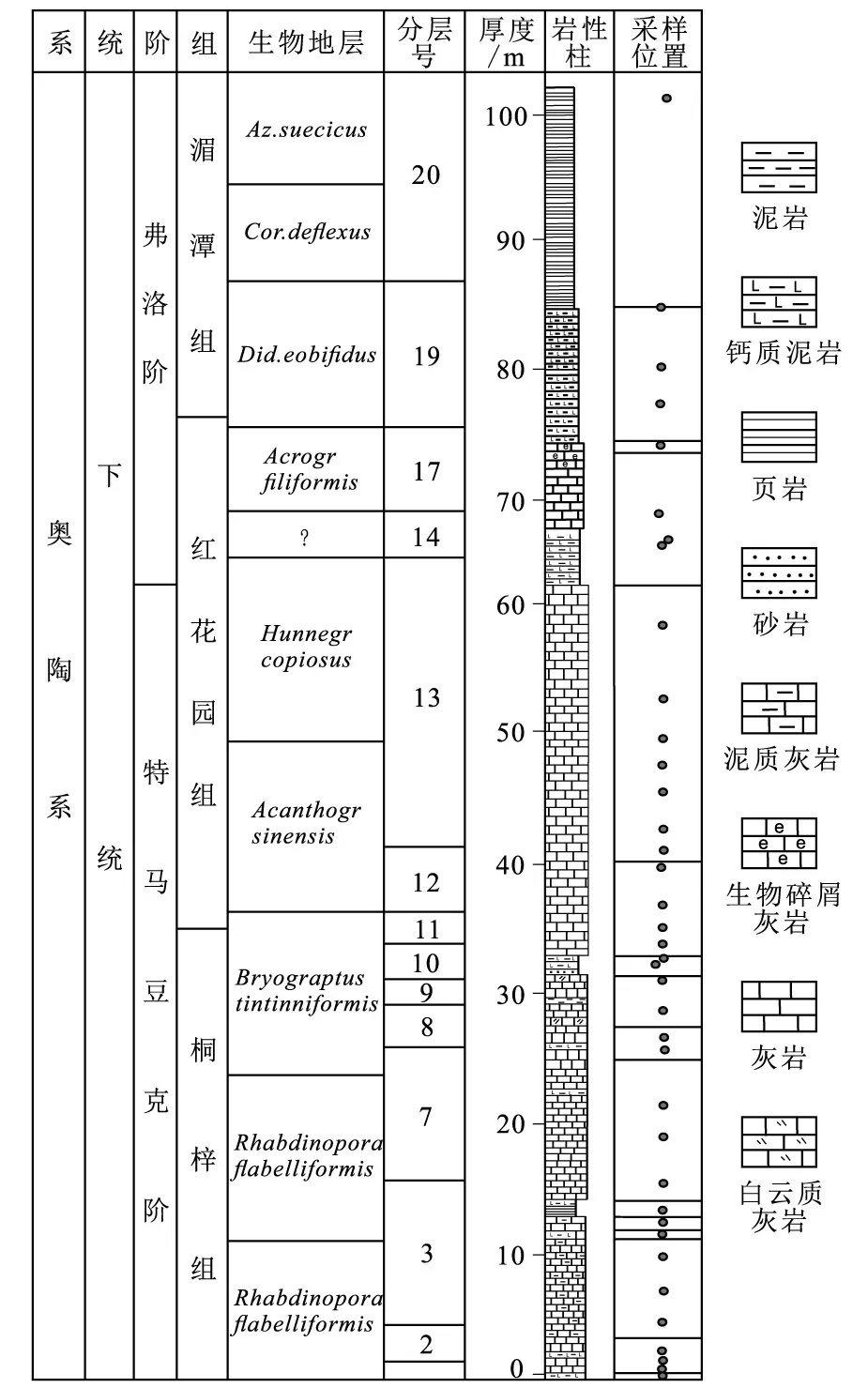
图3 观音桥剖面采样位置Fig.3 Sampling positions of Guanyinqiao section
2 实验方法
在观音桥剖面共采集样品68个,选取其中40个奥陶系样品制备光片。生物量统计的步骤和方法如下:
a.野外样品采集。找寻奥陶纪地层的顶和底,通过和桐梓地区同层位岩性对比划分组段。保证采样间距大致为1~2 m。
b.磨制光片。将选出的40个样品用地质锤敲成适宜磨制薄片的小块,分别编号标记,然后制作薄片。
c.生物量统计。在偏光显微镜下,放大200倍分别对介壳生物碎片和藻类进行生物量统计工作,每个薄片统计20个视域,记录20个视域的生物量总数和计算每个视域的平均生物量(表1)。记录结果统计列入Excel表,并生成纵坐标为生物量、横坐标为以地层由老到新为顺序的样品编号,分别代表藻类和介壳生物含量的折线图,观察它们之间的耦合关系。
另外,由于生物量统计工作中观察到有较多的大型藻(直径>150 μm),如丛生藻和大型红藻等,特将大型藻类所在薄片的采样位置在观音桥剖面柱状图中标记,讨论藻类-环境变化-生物演化间的响应机制。
3 统计结果
根据统计结果可以发现,桐梓组主要有三叶虫、腕足和腹足动物;红花园组主要有腕足、头足和腹足动物,还有一些介形虫(图4-F)、海绵和苔藓;湄潭组主要有三叶虫(图4-D)、腕足、腹足(图4-I)和头足动物(图4-H),还有一些笔石。此外,在桐梓组中部开始出现一些大型蓝绿藻,如丛生藻;到红花园组藻类辐射达到高潮,并在红花园组中部发现了一些藻鲕(图4-A);直至下奥陶统湄潭组藻类辐射开始衰退,与此同时,大型藻类的数量也开始减少。主要的藻类有蓝绿藻(图4-C)、红藻、硅藻和轮藻;大型藻类有丛生藻(图4-B)和前管孔藻(图4-E)。介壳生物从桐梓组开始辐射,到湄潭组介壳生物开始增多。下面选取一些典型的单偏光下生物照片进行介绍。

图4 奥陶纪地层中生物化石显微特征Fig.4 Microscopic characteristics of fossils in Ordovician strata照片均在单偏光下拍摄
图4-A:样品G7-2的显微照片,层位为桐梓组。按显微镜下的面积,微晶方解石占75%,有机质占10%,泥晶方解石胶结物占5%。其中鲕粒被泥晶方解石充填,斑点藻被泥质充填显黑色。
图4-B:样品G8-1的显微照片,层位为桐梓组顶部。按显微镜下的面积,微晶方解石占85%,有机质占2%,泥晶方解石胶结物占13%。其中藻类主要为大型蓝绿藻中的丛生藻,未发现介壳碎片。
图4-C:样品G9-1的显微照片,层位为桐梓组顶部。按显微镜下的面积,微晶方解石占55%,有机质占30%,硅质胶结物占15%。藻类主要为斑点藻,还有极少量的三叶虫碎片;藻球粒主要由圆形或椭圆形的团粒组成。此外有少量砂屑,为硅质磷块岩。
图4-D:样品G13-5的显微照片,层位为红花园组底部。按显微镜下的面积,微晶方解石占77%;介壳碎片占3%;还有少量深褐色有机质团块,占2%;泥晶方解石胶结物占18%。介壳碎片来自角石和三叶虫等,未发现藻类。
图4-E:样品G14-2的显微照片,层位为红花园组顶部。按显微镜下的面积,微晶方解石占75%;生物介壳骨架占10%;少量有机质,占5%;泥晶方解石胶结物占10%。介壳碎片来自三叶虫、腹足类、头足类、苔藓虫和海百合茎等,有大型红藻中的前管孔藻。
图4-F:样品G15-1的显微照片,层位为红花园组顶部。按显微镜下的面积,微晶方解石占75%;生物介壳骨架占20%;少量有机质,占1%;铁泥质胶结物占4%,风化后显褐色。介壳碎片来自苔藓虫、三叶虫、腹足类、头足类、海百合茎和筛管虫等。
图4-G:样品G17-1的显微照片,层位为红花园组顶部。按显微镜下的面积,微晶方解石占79%;介壳碎片占2%;还有少量深褐色有机质团块,占2%;泥晶方解石胶结物占17%。介壳碎片来自角石和三叶虫等。
图4-H:样品G19-1的显微照片,层位为湄潭组底部。按显微镜下的面积,微晶方解石占75%;生物介壳骨架占20%;少量有机质,占1%;铁泥质胶结物占4%,风化后显褐色。介壳碎片来自苔藓虫、三叶虫、腹足类等,未发现藻类。
图4-I:样品G19-2的显微照片,层位为湄潭组。按显微镜下的面积,微晶方解石占75%;生物介壳骨架占12%;少量有机质,占3%;泥晶方解石胶结物占10%。介壳碎片来自三叶虫、腹足类、头足类和棘皮动物等,有少量丛生藻。
4 讨 论
为了能直观地表现本次生物量统计的结果,特把藻类与介壳动物随时间变化的曲线在柱状图中表示,并把大型藻类出现的位置也作了标示(图5)。
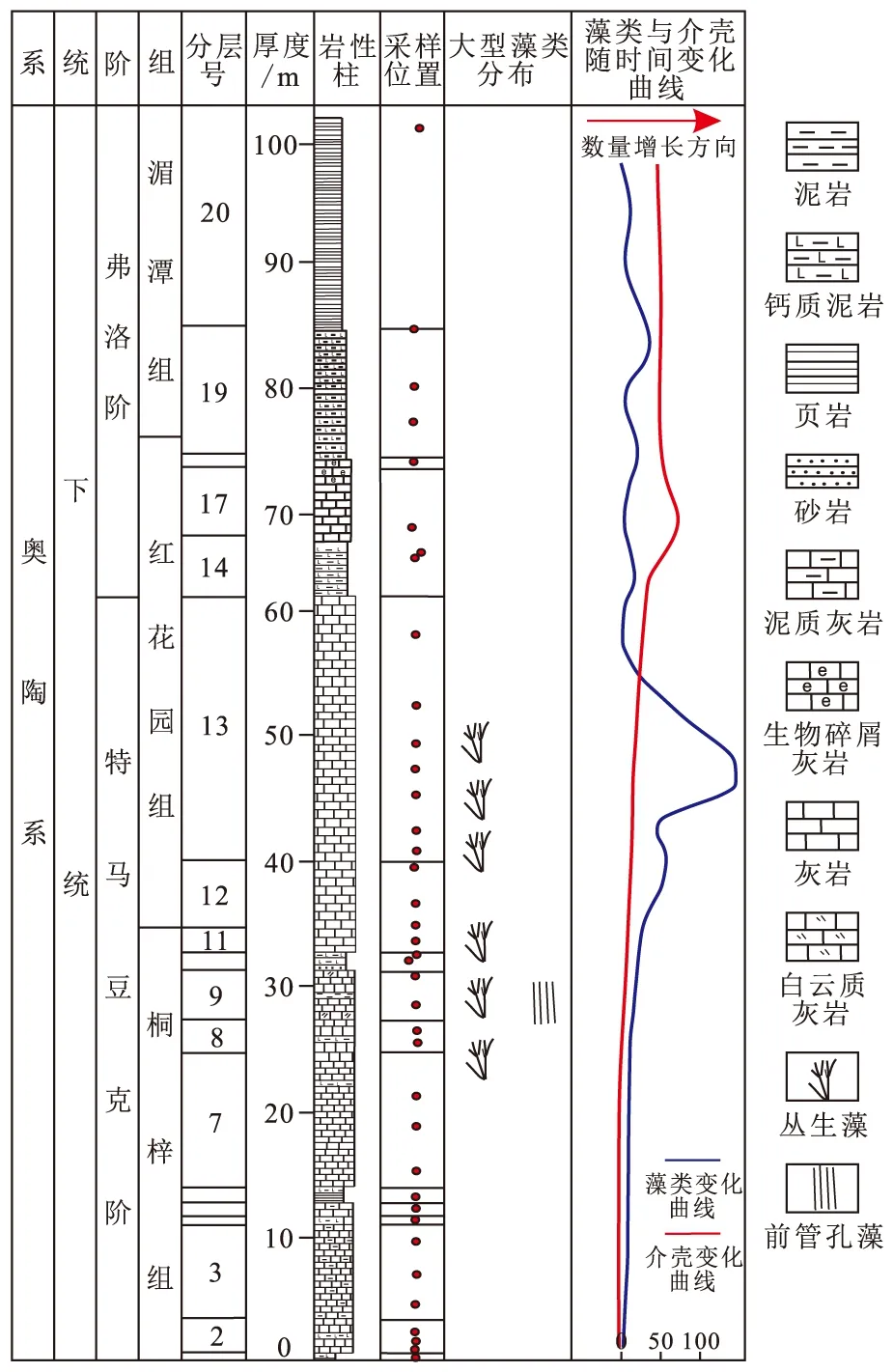
图5 藻类与介壳类生物随时间变化曲线Fig.5 Time curve of algae and shells
在现代及地质历史时期,藻类均为初级生产力及食物链的最低部,对生态环境的稳定性有着决定性的作用。当藻类发生突然变化时,必然也指示与其相关的环境发生了极大变化,从而可能与地质历史时期的生物绝灭事件或生物辐射事件发生耦合。如埃迪卡拉生物群辐射初期,生物标志物指示同时期藻类数量发生了极大增长[18],为开始大量出现的埃迪卡拉生物提供了丰富的食物来源,从而推动了同时期生物辐射事件的进一步发展。在泥盆纪弗拉期-法门期发生的F-F生物绝灭事件,根据生物标志物数据分析[25],指出在生物绝灭发生时期,藻类数量极大增长,可能说明生物绝灭开始之后,大洋发生富营养变化,从而引发赤潮,进一步加大了同时期生物绝灭的强度。而在此次研究中,观音桥剖面所采集样品,有机碳含量不高,不适合进行生物标志物研究,那么是否能利用更加直接的指标(如薄片藻类生物量统计)进行类似研究?下文将利用我们所获得的藻类统计结果与其他研究者关于奥陶纪生物变化曲线、碳同位素曲线、硫同位素曲线、温度曲线及氧气含量变化曲线进行分析,探讨藻类(初级生产力)的变化与早奥陶世生物辐射事件的启动是否具备协同性变化。
如图5所示,第一次生物辐射高潮阶段位于弗洛阶上部区域,生物量在属一级变化上,从990增长至1 180。而在其以下层位,特马豆克阶上部即观音桥剖面红花园组中部,藻类数量持续增长,从200增长至460;之后从特马豆克阶顶部至弗洛阶(红花园组中-上部),藻类数量从460快速减少至40。据此分析,尽管藻类数量增加与生物辐射的第一幕高潮并不具有很强的一致性,但正如K.Pehr等[17]对埃迪卡拉生物辐射事件的研究发现藻类数量增加总是早于埃迪卡拉生物增加一样,藻类是作为介壳生物和埃迪卡拉生物的食物而存在,当藻类数量快速增加之时,也正说明了介壳生物可获取的食物更加丰富,从而导致该种群的快速辐射。正如图6红色方框区域所示,藻类数量的增加或减少与奥陶纪生物辐射的第一幕高峰具有极高的耦合性:首先,藻类增加,促使介壳动物种属增加;其次,在介壳动物种属增加的过程中,作为其食物的藻类数量则开始大规模减少;最后,藻类数量降至其最低数量之时,奥陶纪生物辐射事件的第一幕也就此结束。此种藻类-介壳动物的耦合性变化正说明了藻类数量的增加导致了奥陶纪生物辐射事件的开始。

图6 藻类与生物协变关系图Fig.6 Covariant relationship between algae and organisms(生物和大气中氧含量曲线修改自文献[10];碳同位素曲线,包括硫同位素曲线修改自文献[26];温度曲线修改自文献[28])
那么是什么样的环境变化导致了藻类在此时间段的爆发?据图6所示,由于缺乏相关的硫同位素,我们无法直接进行大洋氧化-还原性的分析;但是根据奥陶纪大气氧气含量曲线[11]不难发现,同时期氧气含量较为稳定,甚至有较小幅度负漂移变化。尽管大气氧含量并无明显变化,但是蓝藻在海洋中的爆发和极速增长,其光合作用会导致大量氧气释放并部分溶于海洋之中,氧气含量曲线也有一个小幅度增长,在第一幕后又轻微下降,与藻类含量变化曲线的峰值具有极好的一致性,这也正与部分研究者所获得的特马豆克阶古海洋氧化的结果一致[26-28]。同一阶段,碳同位素(图6中碳同位素黄色曲线)频繁波动[29],但相对稳定。图6中与藻类变化具有较为明显关系的是古温度变化曲线[30],在藻类数量增加及奥陶纪生物辐射第一幕高峰时期,古温度从37 ℃降至32 ℃。根据现代生态学研究,蓝藻等藻类的最佳生存温度为30~35 ℃,在此温度条件下藻类会大量繁殖[31],因此我们认为温度的下降导致藻类的繁盛,从而为介壳生物的第一幕辐射提供了足够的氧气和食物来源。在介壳动物群种属数量增加之后,藻类被迅速消耗,并在奥陶纪生物辐射事件第一幕高潮之后,一个新的生态体系得以形成,藻类数量与介壳数量处于一个较小变化幅度的稳定时期。需要指出的是,藻类的爆发是发生在辐射之前的,正是由于藻类生物的爆发式增加,才为后续的生物辐射奠定了基础。
5 结 论
a.大型藻类丛生藻在特马豆克阶桐梓组上部及红花园组下部集中出现,明显较其他层位更为繁盛,但藻类数量持续保持一个较小的变化幅度,在丛生藻繁盛后期,藻类数量开始增多,并最终在红花园组中部达到高峰,说明大型藻类的出现对藻类的辐射可能具有极好的指示意义,这需要下一步的更为详细的研究。
b.在藻类数量增加及奥陶纪生物辐射第一幕高峰时期,古温度从37 ℃降至32 ℃。蓝藻等藻类的最佳生存温度为30~35 ℃,因此我们认为温度的下降导致藻类的繁盛,从而为介壳类生物的第一幕辐射提供了足够的氧气和食物。在介壳动物种属数量增加之后,藻类数量迅速消耗,并在奥陶纪生物辐射事件第一幕高潮之后,一个新的生态体系得以形成,藻类数量与介壳数量处于一个较小变化幅度的稳定时期。
c.氧含量变化导致了生物辐射事件的启动与前人所认为的特马豆克阶古海洋氧化的结果一致;藻类繁盛明显早于氧化事件和生物辐射事件,说明藻类促进了氧化事件,氧化事件又促进了生物辐射事件。三者之间存在的促进关系具有很好的研究意义。
目前的研究还存在一些不足。首先,藻类数量的统计存在主观因素的影响且不可消除,只能通过研究者本人对数据进行多视域多对比的操作以确保数据的准确性,工作量极大。其次,早奥陶世初级生产力引发的古海洋氧化事件是具有全球统一性,还仅仅是一种区域性的事件,这需要我们用更多合适的剖面进行同位素地球化学分析进行对比研究。最后,仅靠藻类数量统计,所获得数据较之生物标志物数据的客观性和易解读性明显较弱,因此下一步的研究则是选择一个更为适合的剖面,利用生物标志物及地球化学分析等手段,以验证本文关于藻类爆发和大洋氧含量变化协同机制。
在论文写作中得到成都理工大学何小琴同学的支持和帮助,借此表示感谢。
