配方施肥对杉木幼苗生长及抗氧化特性的影响
2021-07-22赖慧捷李惠通宋政凯侯政杰刘爱琴
赖慧捷,李惠通,宋政凯,苏 铁,侯政杰,刘爱琴*
(1.福建农林大学 林学院,福建 福州 350002;2.国家林草局杉木工程技术研究中心,福建 福州 350002)
【研究意义】氮、磷和钾是植物生长发育的必要元素,微量元素参与了植物体内多种代谢过程,均对植物生长不可缺少[1-3]。长期以来由于树种需肥量与林地土壤养分含量不同,林业生产上盲目施肥现象十分严重,盲目施肥不仅增加苗木培育成本,而且造成肥料浪费,产生环境污染[4-6]。为解决农林业生产上的盲目施肥现象,2005 年开始推广测土配方施肥技术,该技术是在综合考虑土壤供肥能力、植物需肥规律及肥料施用成效基础上,提出植物营养元素用量及其配比的新技术[7-9],在农业生产中得到了广泛应用,但在林业生产上的应用相对较少。因此如何筛选适合不同林地土壤的施肥配方成为当前林业生产中急需解决的重大课题。杉木(Cunninghamia lanceolata(Lamb.)Hook)是我国南方重要的造林树种,具有生长快、产量高、用途广等特点[9],在我国南方得到大面积的人工栽植,在林业生产上占有举足轻重的地位[10]。近年来随着杉木育种水平的提高,高世代杉木良种大量应用,这些杉木良种对林地养分的需求提高,传统的施肥配方已不能适应这些良种苗木培育的要求。【前人研究进展】虽然国内外有关杉木施肥的研究较多,但多以单种肥料的施肥研究居多,对氮磷钾不同配比施肥的研究相对较少,特别是微肥配施的研究更少,缺乏高世代杉木良种育苗的施肥配方,因此极有必要开展配方施肥对促进高世代良种杉木苗木生长的研究。植物在生长代谢及逆境环境下有较高的保护酶系统,其中超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)相互协调在一定程度上能减缓活性氧对植物细胞造成的伤害[14]。丙二醛(MDA)是细胞膜发生脂质过氧化作用而产生,可与SOD、POD和CAT一起反映出植物细胞膜的损伤程度[15-16],但目前有关配方施肥对杉木幼苗抗氧化特性方面的影响研究相对较少。【本研究的切入点】鉴于此,本研究以经二代改良的杉木良种幼苗为研究对象,在传统“3414”N、P、K 配方施肥技术基础上,增加B和Zn微量元素处理,开展杉木幼苗配方施肥盆栽试验,在施肥后测定杉木幼苗生长及其抗氧化系统SOD、POD、CAT 及MDA 含量,探讨不同配方施肥对杉木幼苗生长及抗氧化特性的影响规律。【拟解决的关键问题】筛选出适合二代改良杉木良种苗木培育的施肥配方。
1 材料与方法
1.1 试验材料
以福建省漳平市五一国有林场2代杉木种子园单株采种的半同胞家系种子培育的苗木为研究对象,选择长势一致、生长良好、无病虫害的苗木进行盆栽。
1.2 试验设计
采用“3414”配方施肥技术,选择3 个因素(氮、磷、钾)、每个因素设4 个水平(0、1、2、3),同时增加有无B 和Zn 微肥两种处理(0、2),共设28 个不同施肥处理(表1),每个处理重复4 次,共有试验处理112盆。

表1 不同配方施肥的养分水平Tab.1 Factors and levels of fertilization in different formulations g/kg
1.3 试验处理的实施
为便于控制不同试验处理的养分,采用沙培法进行不同施肥处理的实施。试验在福建农林大学林学院温室大棚进行,从2015 年8 月20 日开始,至2016 年9 月1 日结束,持续观测1 年。栽种盆采用直径19 cm、高18 cm 的聚乙烯塑料花盆。培养基质为过2 mm筛的干净河沙和吸足纯水的聚丙烯酸钠盐保水剂,按3∶1的体积比例混合,每个处理的培养基质干质量3.5 kg(表3)。

表3 试验盆栽培养基质的养分情况Tab.3 Nutrient condition of potted culture substrate
施肥处理分4 次进行,分别在2015 年10 月12 日、2016 年3 月12 日、2016 年6 月12 日和2016 年7 月12 日施入,施用量分别为总量的20%、30%、40%和10%。肥料分别为尿素(含N 46%)、磷酸钙(含P2O516%)、氯化钾(含K2O 60%)、硼酸(含有效硼11%)和EDTA-Zn(含Zn 15%)。
1.4 测定方法
1.4.1 杉木生长指标测定 在施肥12 个月后对不同试验处理杉木进行生长调查,分别测定不同处理杉木的苗高、地径,苗高用T型直尺测量,精确到0.00 cm;地径用游标卡尺测定,精确到0.00 mm。
1.4.2 杉木生理指标测定 在杉木幼苗收获前的2016年8月25日,采集不同试验处理杉木叶片、茎和根系带回实验室进行杉木生理指标测定。采用氮蓝四唑光化还原法[19]测定杉木不同器官超氧化物歧化酶活性,采用愈创木酚法[19]测定过氧化物酶活性和紫外吸收法[19]测定过氧化氢酶活性,采用硫代巴比妥酸法[19]测定丙二醛含量。

表2 不同配方施肥处理试验设计Tab.2 Experimental design of different formulations fertilization treatment
1.5 数据处理
利用SPSS 22.0 软件进行不同试验处理数据分析,采用OriginPro 9 软件绘制数据图表。苗木净生长量=试验结束时苗高/地径-试验初始时苗高/地径;相对生长速率=净生长量/试验处理总月份。
2 结果与分析
2.1 不同配方施肥对杉木幼苗生长的影响
2.1.1 配方施肥对杉木苗高生长的影响 表4可知,不同配方施肥后,杉木苗高生长以N2P2K2处理最高,N0P0K0处理最差。无论是否施微肥,杉木苗高生长均随氮磷钾含量的增加呈先升高后降低趋势,其中以N2P2K2处理的最高。微肥处理组杉木苗高生长占前3的处理分别是N1P2K1、N2P2K2、N2P3K2,无微肥处理组则是N2P2K2、N2P0K2、N2P1K1。多数处理中有微肥处理比无微肥处理苗高生长更好。杉木苗高净生长量和相对生长速率均以N2P2K2处理最高。可见在施微肥条件下氮磷钾均衡施肥对杉木苗高生长的促进效果最好。

表4 不同施肥处理杉木苗高生长的比较Tab.4 Growth condition of Chinese fir seedlings under different fertilization treatment
2.1.2 配方施肥对杉木地径生长的影响 表5可知,不同配方施肥后,杉木地径生长以N2P0K2处理最好,N1P1K2处理最小。不论施微肥与否,杉木地径生长随氮磷钾含量的增加呈先升高后降低趋势,均以N2P2K2处理生长最好。在施微肥处理组中杉木地径生长占前3的处理分别是N2P2K2、N1P2K1、N1P2K2,在没有施微肥处理组中为N2P0K2、N1P2K1、N2P2K2处理。多数施微肥处理地径生长均高于没有施微肥处理,个别处理略低可能跟其本底值偏小有关。杉木地径净生长量及相对生长速率均以N2P2K2处理最高。可见施微肥条件下氮磷钾均衡施肥对促进杉木地径生长的效果最好。
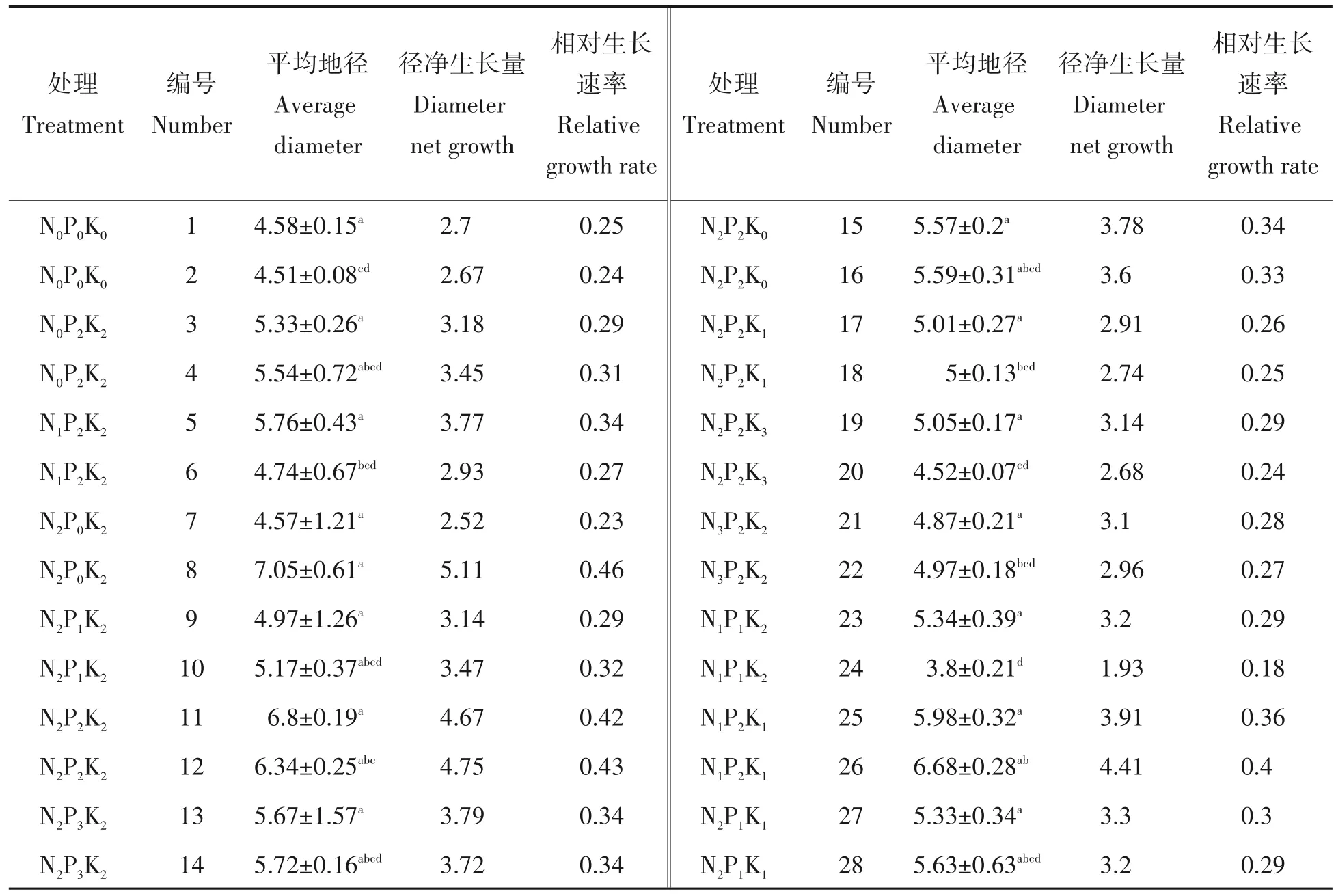
表5 不同施肥处理杉木地径生长的比较Tab.5 Growth condition of Chinese fir seedlings under different fertilization treatment
2.2 不同配方施肥对杉木抗氧化特性的影响
2.2.1 配方施肥对杉木SOD 的影响 图1 可知,不同配方施肥对杉木不同器官SOD 活性的影响达极显著水平(P<0.01),表现为叶>茎>根,杉木地上部分SOD 活性明显高于地下部分。在未施微肥处理中,杉木根、茎、叶的SOD 活性分别以N2P3K2、N0P2K2和N3P2K2处理最高。在施微肥处理中,杉木根、茎、叶SOD 活性分别以N3P2K2、N0P0K0和N0P2K2处理最高。在未施微肥条件下不同N 水平处理对杉木SOD 活性的影响达显著水平(P<0.05),随N 的增加,根系SOD 活性总体呈下降趋势;茎和叶SOD 活性均呈先降低后升高趋势,以N2P2K2处理的活性最低。不同P 水平处理对杉木根和叶SOD 活性的影响达显著水平(P<0.05),随P 的增加,根系SOD 活性总体呈上升趋势,茎和叶SOD 活性均呈先下降后上升趋势。不同K 水平处理对杉木SOD 活性的影响达显著水平(P<0.05),随K 的增加,杉木各器官SOD 活性均呈先下降后上升趋势。在施微肥处理条件下除茎外不同N、P、K 水平处理对杉木SOD 活性的影响达显著水平(P<0.05)。

图1 不同施肥处理杉木超氧化物歧化酶活性的比较Fig.1 SOD activity of Chinese fir seedlings under different fertilization treatments
2.2.2 配方施肥对杉木苗木POD 活性的影响 图2 可知,配方施肥对杉木不同器官POD 活性的影响均达极显著水平(P<0.01),表现为叶大于根和茎。在未施微肥处理中,杉木根、茎、叶POD 活性分别以N1P1K2、N2P3K2和N2P1K2处理最高。在施微肥处理中,杉木苗木根、茎、叶POD 活性分别以N2P2K1、N1P2K2和N2P1K2处理最高。在没有施微肥条件下除根POD 活性在不同P 水平下无显著差异外,不同N、P、K 处理对杉木SOD 活性的影响均达显著水平(P<0.05)。随N 的增加,根和茎的POD 活性呈先下降后升高趋势,叶则反之。随P 和K 的增加,根和叶的POD 活性均呈先升高后降低趋势,茎则反之。在施微肥条件下,除茎的POD 活性在不同K 水平下无显著差异外,不同N、P、K 处理对杉木POD 活性的影响均达显著水平(P<0.05)。随N、K 的增加,根和茎的POD 活性总体呈先下降后升高趋势,叶则反之;随P的增加,根的POD活性呈先下降后升高趋势,叶则反之。

图2 不同施肥处理杉木过氧化物酶活性的比较Fig.2 POD activity of Chinese fir seedlings under different fertilization treatments
2.2.3 配方施肥对杉木苗木CAT 活性的影响 图3 可知,不同配方施肥对杉木CAT 活性的影响均达极显著水平(P<0.01),表现为根大于茎和叶。在未施微肥条件下杉木根、茎、叶CAT 活性分别以N0P2K2、N0P0K0和N1P2K2处理最高。在施微肥条件下杉木苗木根、茎、叶CAT 活性分别以N1P2K2、N2P2K2和N0P2K2处理最高。
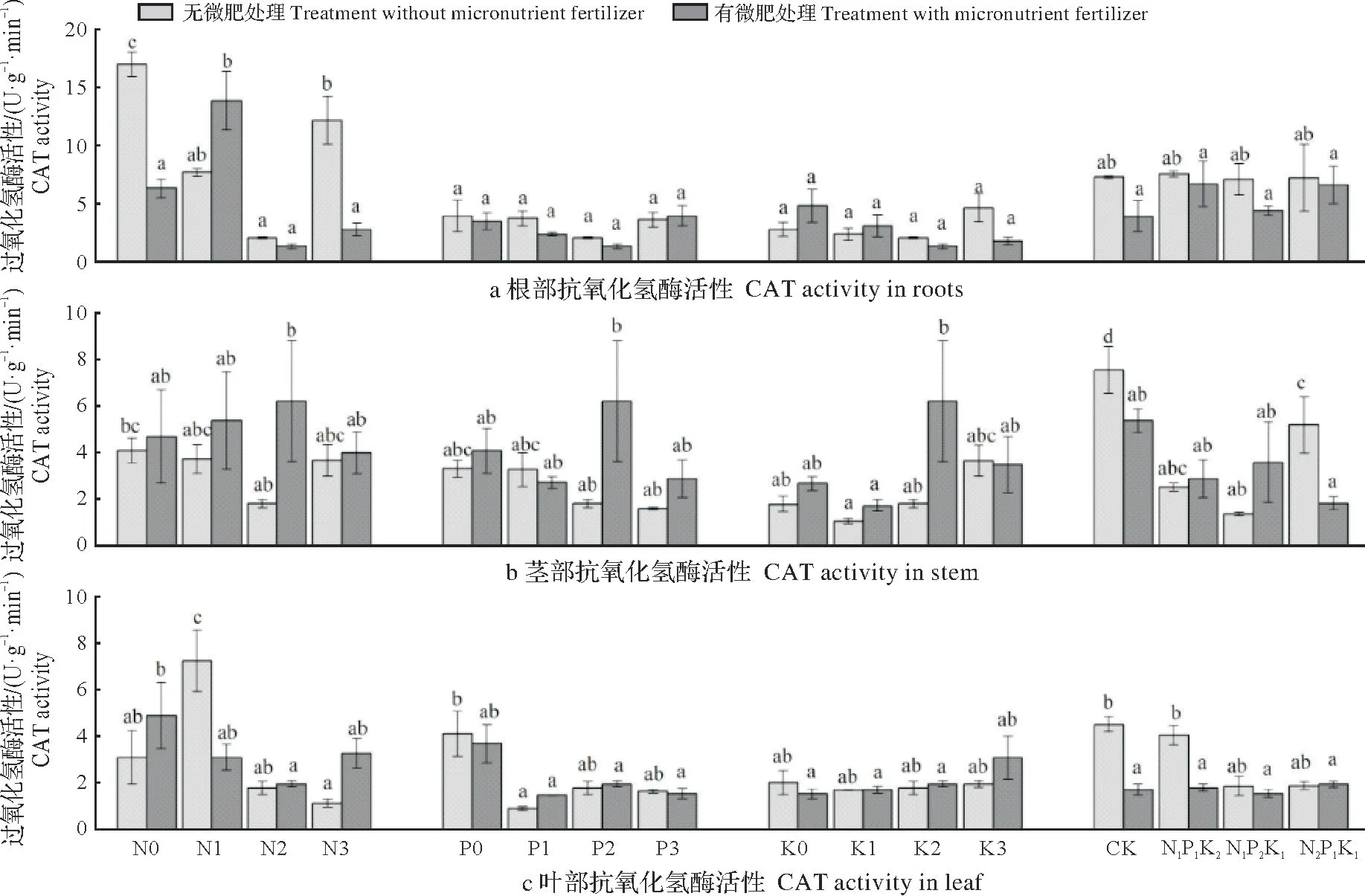
图3 不同施肥处理杉木过氧化氢酶活性的比较Fig.3 CAT activity of Chinese fir seedlings under different fertilization treatments
经方差分析,根系CAT 活性在没有施微肥条件下不同P水平条件下无显著差异,茎在施微肥条件下不同N水平下无显著差异,叶CAT活性在不同K水平下均无显著差异,其余不同N、P、K水平对杉木CAT活性的影响均达显著水平(P<0.05)。在未施微肥条件下,随N 的增加,根和茎CAT 活性呈先降低后升高趋势,叶则反之。随P 的增加,根和叶的CAT 活性呈先降低后升高趋势,茎则呈降低趋势;随K 的增加,杉木CAT活性均呈先降低后升高趋势。在施微肥处理下,随N的增加,根和茎CAT活性呈先升高后降低趋势,叶则反之;随P的增加,根和叶的CAT 活性呈先下降后升高趋势,茎则反之;随K 的增加,根CAT 活性呈先下降后升高趋势,茎则反之,叶则呈上升趋势。
2.2.4 配方施肥对杉木苗木MDA 含量的影响 图4可知,不同配方施肥对杉木MDA 含量的影响均达极显著水平(P<0.01),施肥对杉木苗木的MDA含量有较大影响。在未施微肥条件下杉木根、茎、叶MDA含量分别以N2P3K2、N0P0K0和N0P2K2处理最高。在施微肥条件下杉木根、茎、叶MDA 含量分别以N2P3K2、N2P2K3和N3P2K2处理最高。
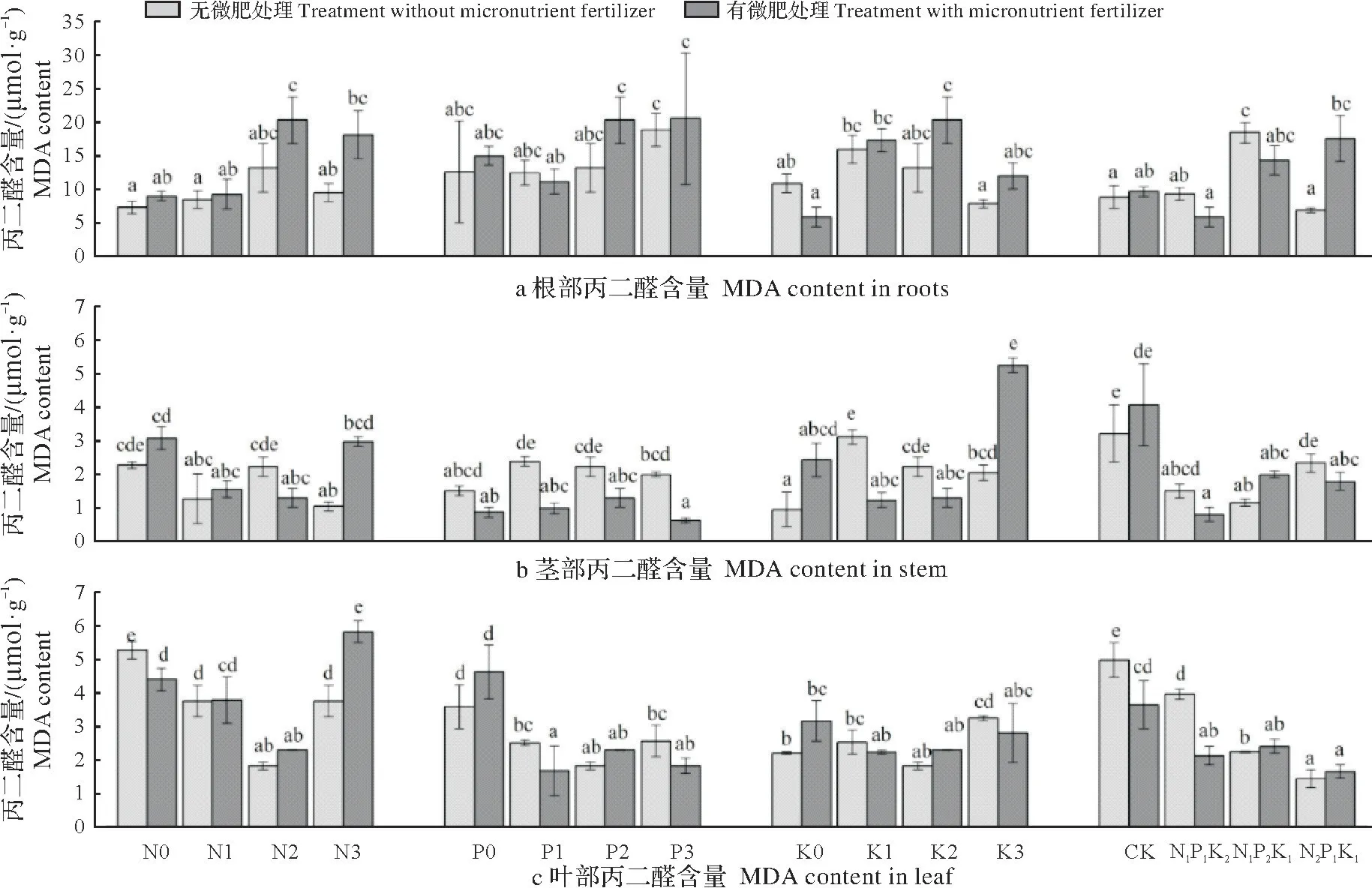
图4 不同施肥处理杉木丙二醛含量的比较Fig.4 The subjection function of Chinese fir seedlings under different fertilization treatments
经方差分析,杉木根系MDA 含量在不同P 水平下无显著差异,叶在施微肥不同K 水平下无显著差异,其余不同N、P、K 水平对杉木MDA 含量的影响达显著水平(P<0.05)。在未施微肥条件下,随N 的增加,根MDA 含量呈先升高后降低趋势,叶则反之,茎呈先降低后升高再降低趋势。随P 的增加,根的MDA 含量呈升高趋势,茎则呈先升高后降低趋势。随K 的增加,根和茎的MDA 含量呈先升高后降低趋势。在施微肥条件下,随N的增加,根MDA含量呈先升高后降低趋势,茎和叶则反之。随P的增加,根和叶的MDA含量呈先下降后升高趋势;随K的增加,根MDA含量呈先升高后降低趋势。
2.3 不同配方施肥对杉木抗氧化特性影响的主成分分析
利用主成分分析法进行不同施肥处理杉木的苗高、地径、SOD、POD、CAT 和MDA 等指标的主成分分析,综合评价不同施肥配方对杉木生长及抗氧化特性的影响。结果表明:第一主成分中承载系数较大的指标为CAT、SOD 和POD,其中POD 产生负向影响。第二主成分中承载系数较大的指标为地径和苗高。第三主成分中承载系数较大的指标为MDA,前3 个主成分方差贡献率分别为33.98%、31.22%、17.10%,累计贡献率达82.30%,说明这3 个主成分可以替代6 个指标来综合分析不同配方施肥对杉木生长及抗氧化特性的影响。数值越高表明该处理对杉木生长的促进作用及抗氧化能力越强。
表6 可知,不论施微肥与否,均以N2P2K2处理对杉木苗高和地径生长的促进效果最好。未施微肥处理条件下,不同配方施肥处理施肥效果的排序均以中量配方的处理最好。在施微肥后多数配方施肥处理的综合评价排序提高,表明在土壤养分不足时微肥能够一定程度上缓解大量元素的不足,有利于杉木生长。当大量元素过剩时,微肥则能减缓养分过量对杉木生长的影响。在不同处理中N1P1K2的施肥效果排序较高,说明杉木生长及其抗氧化能力对P的需求比对N、K的需求更大。
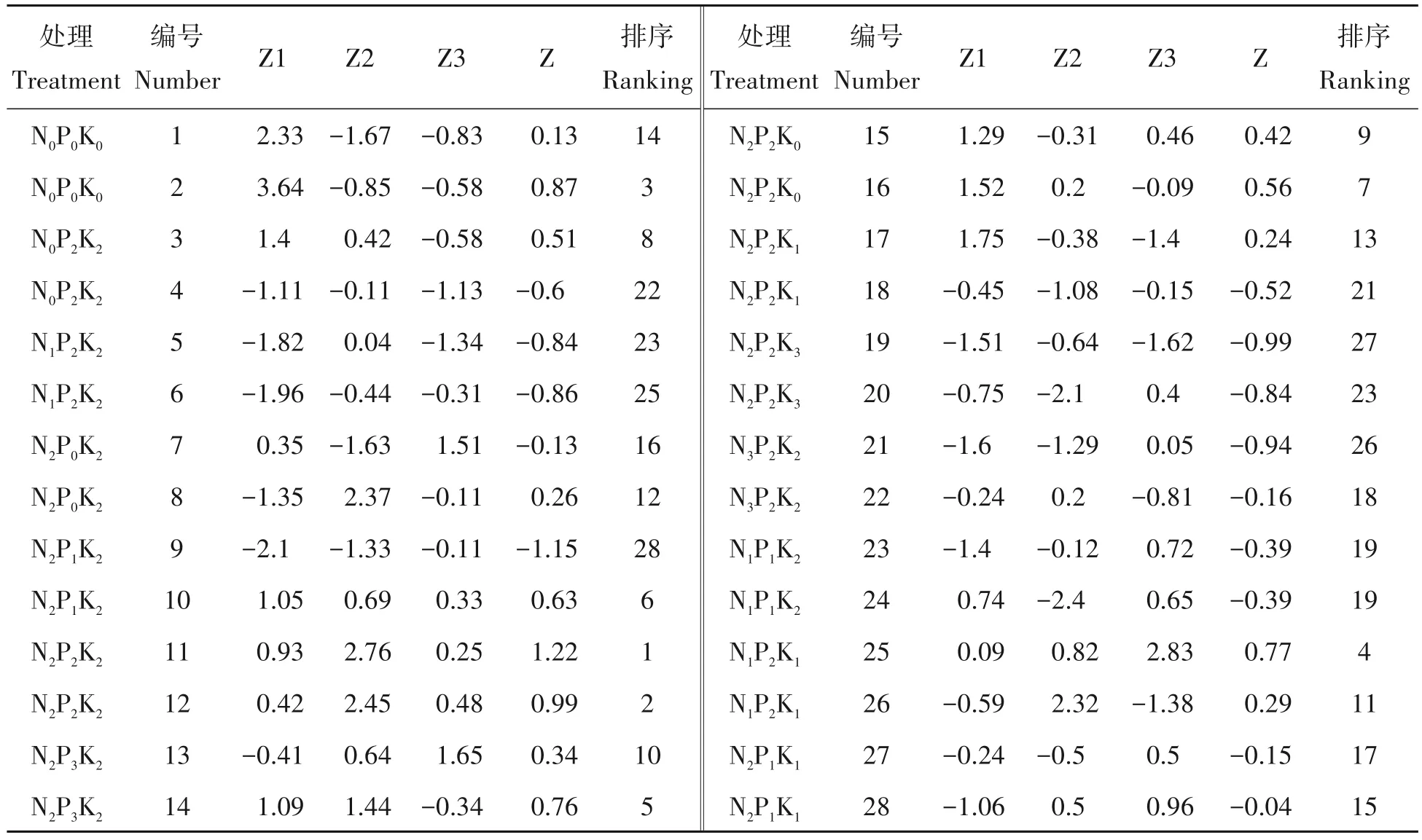
表6 不同施肥处理的主成分分析Tab.6 Score and comprehensive score of principal components
3 讨论
大量研究表明,施肥对杉木幼苗的生长有促进作用,能提高苗木质量,但过量施肥会使幼苗产生毒害,影响其生长[21]。微量元素B、Zn 能促进植物对N、P、K 的吸收利用,提高N、P、K 肥料的利用率[22]。本研究发现,在不同N、P、K配方施肥中,杉木幼苗生长以氮磷钾均衡配施处理最好,微量元素B、Zn的施用能明显提高杉木苗木质量,说明氮磷钾均衡施肥能促进杉木幼苗生长,微肥能提高杉木对大量元素的吸收,进而提升杉木苗木质量,与前人研究结果一致。
SOD、POD 和CAT 是植物在生长代谢和受到环境胁迫时消除产生的游离氧的关键酶,MDA 是膜脂过氧化过程中主要产物[24]。抗氧化酶活性及MDA 含量是评定植物对生存环境响应的判定指标。在遭受逆境时,植物细胞产生的活性氧含量增加或抗氧化酶活性降低[25-26],合理施肥对提高植物叶片抗氧化酶活性及降低MDA 含量有显著作用[27],根系对环境变化的胁迫更为敏感[28]。根和茎中起抗氧化作用的关键酶是SOD 和POD,而叶起主要作用的关键酶为CAT[29]。本研究表明,不同配方施肥下杉木SOD 和POD 活性总体表现为叶>茎>根,CAT 活性表现为根>茎>叶;MDA 含量表现为根>叶>茎,与前人研究结果一致。
当施肥过量或不足时,植物POD 活性降低,SOD 和CAT 活性则增强[30]。赵落含等[28]研究发现施氮降低了冬小麦叶片SOD 和POD 活性及根系POD 活性,但增加根系SOD 和CAT 活性,当施氮量达到一定程度时叶片和根系SOD 活性均升高,中氮处理下冬小麦总体抗氧化能力增加,降低了养分胁迫的影响。本研究发现,随N、P、K 施入量的增加,杉木不同器官抗氧化酶活性呈先降低后增加趋势。杉木根系MDA 含量随氮、钾的增加呈先增加后减少,茎和叶则反之,说明氮、钾不足及过量对杉木根系细胞膜的影响不明显。
大量研究表明,微量元素与大量元素存在协同作用,Zn 是Cu/Zn-SOD 的结构组成部分,具有稳定SOD酶活性的作用,能促进植物对N、P、K的吸收利用,提高N、P、K肥料的利用率[31-32]。本研究发现,B和Zn 等微量元素施入后,随氮磷钾的增加,杉木不同器官的抗氧化酶活性变化与未施微肥处理基本一致,但能显著降低中氮、低钾和低磷处理杉木茎的MDA 含量,说明微肥能降低氮、磷不足及过量时对杉木叶部细胞膜损伤,从而促进杉木幼苗对N、P的吸收与利用,与前人的研究结果基本一致。
4 结论
不同配方施肥对杉木苗高和地径生长有显著促进作用,并能提高杉木抗氧化酶活性,在一定程度上降低丙二醛含量。微肥能调节杉木对土壤氮磷钾元素的需求量及比例,施微肥处理的促进效果比未施微肥处理更好。在不同施肥处理中,以N2P2K2处理对杉木生长的促进效果最好,是值得生产上推广的杉木良种育苗配方。
