田间二化螟雄蛾对不同配比性信息素的嗅觉反应及性信息素识别相关基因表达水平差异
2021-07-07刘天伟陈运康许春梅杜永均
刘天伟, 陈运康, 许春梅, 郭 荣, 冯 波, 杜永均,*
(1.浙江大学农药与环境毒理研究所, 杭州 310058; 2.湖南攸县植保植检站, 湖南攸县 412300;3.犍为县农业农村局, 四川犍为 614400; 4.全国农业技术推广服务中心, 北京 100125; 5.温州医科大学健康与环境生态研究所, 浙江温州 325035)
昆虫在性成熟时释放的性信息素气味分子,通过气流扩散传播,被其同种异性昆虫的嗅觉感器感受产生神经生理反应,并整合其他感觉通道,最后使雄性昆虫产生朝向气味源的定向飞行行为反应,找到配偶而实现交配。在嗅觉感器内部,已经鉴定出许多基因家族在昆虫的性信息素嗅觉识别过程中发挥重要作用,包括性信息素结合蛋白(pheromone binding proteins, PBPs)(Zhuetal., 2016)、性信息素受体(pheromone receptors, PRs)(Linetal., 2015; Zhangetal., 2017)、细胞色素P450(cytochrome P450)(Fengetal., 2017)、普通气味结合蛋白(general odorant binding proteins, GOBPs)(Khuhroetal., 2017)、离子受体(ionotropic receptors, IRs)(Heetal., 2019)、感觉神经元膜蛋白(sensory neurone membrane proteins, SNMPs)(Sunetal., 2019)、化学感受蛋白(chemosensory proteins, CSPs)(Zengetal., 2018)和气味降解酶(odour degrading enzymes, ODEs)(Chertempsetal., 2012)。已经发现性信息素结合蛋白PBP1-4(Changetal., 2015; Dongetal., 2019)、普通气味结合蛋白GOBP1-2(Gongetal., 2009; Khuhroetal., 2017)和性信息素受体PR1-6(Changetal., 2015)参与了二化螟Chilosuppressalis性信息素的嗅觉识别。因此,这些基因的表达水平直接影响了神经输入信号,从而最终影响雄蛾的行为反应。
首先,性信息素的生物合成原料来自寄主植物,寄主植物是影响昆虫性信息素组成变异的一个重要因子(Landolt and Phillips, 1997)。草地贪夜蛾Spodopterafrugiperda玉米品系雌蛾几乎不与水稻品系雄蛾交配,而水稻品系雌蛾与两类品系的雄蛾均可交配,这是由于雌蛾性腺分泌的性信息素不同而造成的行为隔离(Caas-Hoyosetal., 2017)。玉米螟性信息素也受寄主植物的影响(徐翔等, 2018)。其次,昆虫性信息素的生物合成受一系列酶控制,环境中的多种生物和非生物因子影响其相应基因的表达水平,这些基因的表达水平的变异最后影响了雌成虫释放的性信息素组成和配比差异,尤其是微量组分的差异(Araietal., 1984; Linetal., 2018),从而形成性信息素的多型现象(Svenssonetal., 2002; Grootetal., 2014)。而与此相应的,其同种异性的雄蛾嗅觉系统也需要适应其变异并协同进化,由此形成了嗅觉的个体差异(Linetal., 2015; Zhangetal., 2017)。最早是在黑腹果蝇Drosophilamelanogaster中开展了自然种群的嗅觉行为的个体差异及其分子遗传机理研究(Rollmannetal., 2010; Wangetal., 2010)。在斜纹夜蛾Spodopteralitura的性信息素行为调控的雌性性信息释放的变异与雄蛾的嗅觉适应性的协同进化中,同样形成了嗅觉的个体差异,并由此形成了性信息素行为调控的地理种群差异(Linetal., 2015; Zhangetal., 2017; Linetal., 2018)。
二化螟C.suppressalis属鳞翅目螟蛾科,是亚洲、非洲北部和欧洲南部等热带及亚热带地区一种重要农业害虫(Yuetal., 2010)。长期以来,二化螟主要依靠化学农药进行防治,导致其抗药性水平快速上升,新农药的研发跟不上农药产生抗性的速度,一些地区出现了无药可用的地步,对水稻生产造成了巨大的损失(谷云勇等, 2018)。为了减少施用化学农药,基于性信息素的群集诱杀技术(苏建伟等, 2003; 左文等, 2008; Vacasetal., 2009; 杜永均等, 2013)和交配干扰技术(Ueno and Hayasaka, 1997; Alfaroetal., 2009; Chenetal., 2014)在二化螟的绿色防控中发挥了重要作用。二化螟雌蛾释放顺11-十六碳烯醛(Z11-16∶Ald)、顺9-十六碳烯醛(Z9-16∶Ald)、顺13-十八碳烯醛(Z13-18∶Ald)等组成的混合物(Tatsukietal., 1977; Mochidaetal., 1984; Chenetal., 2018)。最新研究发现,性信息素引诱的二化螟雄蛾具有更强的性信息素反应能力,同时性信息素引诱的大部分雄蛾为未交配状态(冯波等, 2017; 郭前爽等, 2019),这些研究结果从理论上解释了性信息素群集诱杀二化螟的防控有效性。由于二化螟是寡寄主习性,寄主植物包括水稻、茭白、玉米、小麦、油菜等植物(Houetal., 2010),这些作物在不同地区有不同的耕作方式和品种,二化螟的性信息素受寄主植物差异的影响显著(Cuong and Cohen, 2003; Uenoetal., 2006)。但是到目前为止还不清楚同种群二化螟的不同地理种群雄蛾是否对性信息素识别存在差异。
为了系统研究二化螟雄蛾嗅觉的地理种群差异及其机理,本研究利用田间试验分析不同配比性信息素对不同地理种群二化螟的诱虫量差异,利用风洞试验测定不同配比性信息素从同一种群引诱获得的雄蛾在室内对不同配比性信息素的嗅觉行为反应差异,利用RT-qPCR测定不同配比性信息素引诱同一种群二化螟雄蛾触角中与性信息素识别相关基因的表达差异。本研究结果首次证实了不同田间种群二化螟雄蛾对不同配比性信息素的反应差异,有助于阐述二化螟的嗅觉识别机理和制定有效的性信息素防控策略。
1 材料与方法
1.1 田间诱捕试验
2019年4-8月在浙江省杭州市萧山区水稻种植区(表1)选择长势基本均匀、管理基本一致的6个试验点,进行二化螟的田间诱捕试验,分别诱捕水稻田中越冬代和第2代二化螟雄蛾,每日7∶00-8∶00时将田间诱捕器内的雄虫取出,记录每个诱捕器的诱蛾量,带回浙江大学应用化学生态学实验室饲养,以备风洞试验以及RT-qPCR的试验。我们还在湖南省株洲市攸县、安徽省宣城市泾县、四川省眉山市东坡区、辽宁省辽阳市太子河区、吉林省长春市德惠市的水稻种植区进行了田间诱捕试验(表1),每日7∶00-8∶00时记录每个诱捕器的诱蛾量,并将田间诱捕器内的雄蛾清除。试验用二化螟性信息素PVC毛细管诱芯和新型飞蛾诱捕器都由宁波纽康生物技术有限公司提供。每枚诱芯中的信息素总含量为1.2 mg,Z11-16∶Ald和Z9-16∶Ald 的比例分别为540 μg∶540 μg, 864 μg∶216 μg, 945 μg∶135 μg, 980 μg∶98 μg, 1 003 μg∶77 μg, 1 016 μg∶64 μg和1 045 μg∶35 μg,Z13-18∶Ald的含量维持不变。每个诱捕器安装一枚诱芯,诱捕器高度约1 m,试验点按梯形放置,一行放置3个处理,另一行放置4个处理。处理之间间隔为30 m,两行间隔为30 m,两个试验点间隔为200 m。各试验点不同诱芯按不同排列顺序,以减少位置和风向的影响。

表1 本研究的田间诱捕试验地点
1.2 风洞试验
1.2.1风洞装置:长方体型风洞外壁由透明的有机玻璃和铝合金框架制成,长2 m,宽和高各1 m。风源为标准型净化工作台,风速为0.3 m/s。风源对侧装有排气扇可将带有化合物的气流排出室外。试验在红色光源中进行,光强度为1.0~1.5 lx。风洞温度为23~25℃、相对湿度为60%~70%。
1.2.2雄蛾行为测试:将田间取回的雄虫用10%的蜂蜜水喂养,饲养温度为25±2℃,相对湿度为75%±10%,光照条件为自然光照(光周期14L∶10D),于当晚22∶00时至次日凌晨进行风洞试验,每头雄蛾只使用一次。将诱芯固定于距风源50 cm、距顶部玻璃50 cm的鳄鱼钳上。在距下风端50 cm处置一高50 cm的释放支架,受试雄蛾置于其上。如雄蛾 5 min内无行为反应则换下,引入另一头雄蛾继续试验。试验中每个代次每种诱芯测试50~60头雄蛾,记录每头雄蛾的行为反应类型,包括静止、起飞、直飞、着陆于诱芯管上等。各种行为的比例=具有该行为的虫量/试验总虫量×100%。
1.3 RNA提取、cDNA合成及RT-qPCR分析
诱捕到雄蛾当晚21∶00时对田间样品取样,用于基因表达水平的测定。按照说明书用RNAiso Plus提取总RNA。提取的RNA溶于RNase-Free水中,经分光光度计测定浓度和OD值,然后按照说明书使用PrimeScriptTMRT Reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa)合成cDNA。cDNA的合成为两步法,第一步为10 μL体系,包括1 μL总RNA, 2 μL 5×gDNA Eraser Buffer, 1 μL gDNA Eraser和6 μL RNase-free水。第二步为20 μL体系,包括第一步的反应液10 μL, 1 μL PrimeScript RT Enzyme Mix I, 1 μL RT Primer Mix, 4 μL 5×PrimeScript Buffer 2(for Real Time)和4 μL RNase-free dH2O。所用的条件为42℃ 2 min, 37℃ 15 min和85℃ 5 s。所有样品均设置3个生物学重复。
参考转录组测序(Caoetal, 2014)的分析结果,设计引物的序列(表2),并选择Actin和TUB为内参基因,以不同诱芯诱捕雄虫触角的cDNA为模板,用RT-qPCR对不同配比诱捕到的二化螟雄蛾触角检测性信息素识别相关基因PR1-PR6,PBP1-PBP4和GOBP1-GOBP2的表达水平。RT-qPCR反应体系(20 μL): TB Green Premix Taq II 10 μL, 上下游引物(10 μmol/L)各1 μL, ROX Reference Dye 0.4 μL, DNA模板2 μL, 灭菌水5.6 μL。其中DNA模板为cDNA稀释4倍的溶液。RT-qPCR在实时定量PCR仪(Bio-Rad)上完成,每个样品重复6次。采用两步法qPCR扩增程序:预变性95 ℃ 3 min;95℃ 10 s, 55℃ 30 s, 39轮循环。所有的定量PCR数据用2-ΔΔCt方法进行分析。

表2 本研究所用引物
1.4 数据分析
数据分析采用软件SPSS 20.0。多组平均数间比较采用One-way ANOVA法,差异显著性分析采用LSD(least significant difference)法和Tamhane’s T2法;风洞试验数据采用交叉表法,差异显著性分析采用卡方检验(Chi-square test)法。差异显著性分析时P>0.05表示差异不显著,P<0.05则表示差异显著。
2 结果
2.1 不同配比性信息素对水稻二化螟的雄蛾诱蛾量占比比较
Z11-16∶Ald和Z9-16∶Ald不同配比性诱芯在全国6省进行诱捕试验,结果显示7种不同配比性诱芯在田间对水稻二化螟雄蛾均有引诱作用(图1)。在吉林省,1 045 μg∶35 μg和1 016 μg∶64 μg配比诱芯引诱的雄蛾数量显著多于540 μg∶540 μg, 980 μg∶98 μg和1 003 μg∶77 μg配比诱芯引诱的,再显著多于配比为864 μg∶216 μg和945 μg∶135 μg诱芯引诱的。在辽宁省,1 045 μg∶35 μg, 1 016 μg∶64 μg, 980 μg∶98 μg和945 μg∶135 μg配比诱芯引诱的雄蛾数量显著多于540 μg∶540 μg和864 μg∶216 μg配比诱芯引诱的。在安徽省,1 016 μg∶64 μg和980 μg∶98 μg配比诱芯引诱的雄蛾数量显著多于1 003 μg∶77 μg和864 μg∶216 μg配比诱芯引诱的,再显著多于540 μg∶540 μg配比诱芯引诱的。在四川省,1 003 μg∶77 μg, 980 μg∶98 μg和945 μg∶135 μg配比诱芯引诱的雄蛾数量显著多于1 045 μg∶35 μg配比诱芯引诱的,再显著多于540 μg∶540 μg配比诱芯引诱的。在湖南省,1 045 μg∶35 μg配比诱芯引诱的雄蛾数量显著多于其余配比诱芯引诱的,1 016 μg∶64 μg和980 μg∶98 μg配比诱芯引诱的雄蛾数量显著多于945 μg∶135 μg和864 μg∶216 μg配比诱芯引诱的,再显著多于540 μg∶540 μg配比诱芯引诱的。在浙江省,1 045 μg∶35 μg配比诱芯引诱的雄蛾数量显著多于其余配比诱芯引诱的,540 μg∶540 μg 配比诱芯引诱的雄蛾数量显著低于864 μg∶216 μg配比诱芯引诱的,再显著低于其他配比诱芯引诱的,980 μg∶98 μg配比诱芯引诱的雄蛾数量显著低于1 003 μg∶77 μg,1 016 μg∶64 μg和1 045 μg∶35 μg配比诱芯引诱的。上述6省二化螟性信息素诱芯Z11-16∶Ald和Z9-16∶Ald的最佳配比如下:吉林省为1 045 μg∶35 μg和1 016 μg∶64 μg,辽宁省为1 045 μg∶35 μg, 980 μg∶98 μg和945 μg∶135 μg,安徽省为1 016 μg∶64 μg和980 μg∶98 μg,四川省为1 003 μg∶77 μg, 980 μg∶98 μg和945μg∶135 μg,湖南省和浙江省为1 045 μg∶35 μg。同一配比诱芯在上述6省引诱蛾量明显不同,导致7种不同配比诱芯的诱蛾比例在吉林省、辽宁省、安徽省、四川省、湖南省和浙江省存在差异。结果表明,不仅诱蛾量最大的诱芯性信息素最佳配比在不同地区间存在差异,而且不同配比性信息素的诱蛾比例在不同地区间也存在差异(图1)。
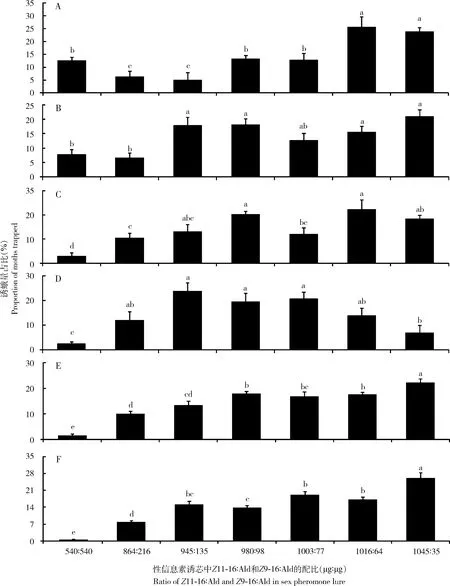
图1 中国6省水稻田间不同配比性信息素诱芯诱捕的二化螟雄蛾的诱蛾量占比
2.2 越冬代和二代水稻二化螟的风洞行为分析
利用田间性诱的越冬代水稻二化螟进行风洞行为测定时,发现二化螟雄蛾对各种诱芯的起飞率为99.3%~100%。Z11-16∶Ald和Z9-16∶Ald配比为980 μg∶98 μg诱芯诱捕的田间雄蛾在室内风洞试验中对980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是15.00%, 15.38%和10.53%,但是这些雄蛾均未着陆到诱芯上(图2: B)。864 μg∶216 μg配比诱芯诱捕的田间雄蛾在风洞中对980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是6.67%, 28.89%和4.65%,仅有4.4%的雄蛾到达864 μg∶216 μg配比诱芯,差异显著性分析表明864 μg∶216 μg配比诱芯诱捕雄蛾对864 μg∶216 μg配比诱芯的行为反应显著强于对980 μg∶98 μg和1 016 μg∶64 μg配比诱芯(图2: A)。1 016 μg∶64 μg配比诱芯诱捕的田间雄蛾对980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是6.67%, 8.89%和35.56%,有8.89%的雄蛾到达1 016 μg∶64 μg配比诱芯,差异显著性分析表明1 016 μg∶64 μg配比诱芯诱捕的雄蛾对1 016 μg∶64 μg配比诱芯的行为反应显著强于对980 μg∶98 μg和864 μg∶216 μg配比诱芯(图2:C)。
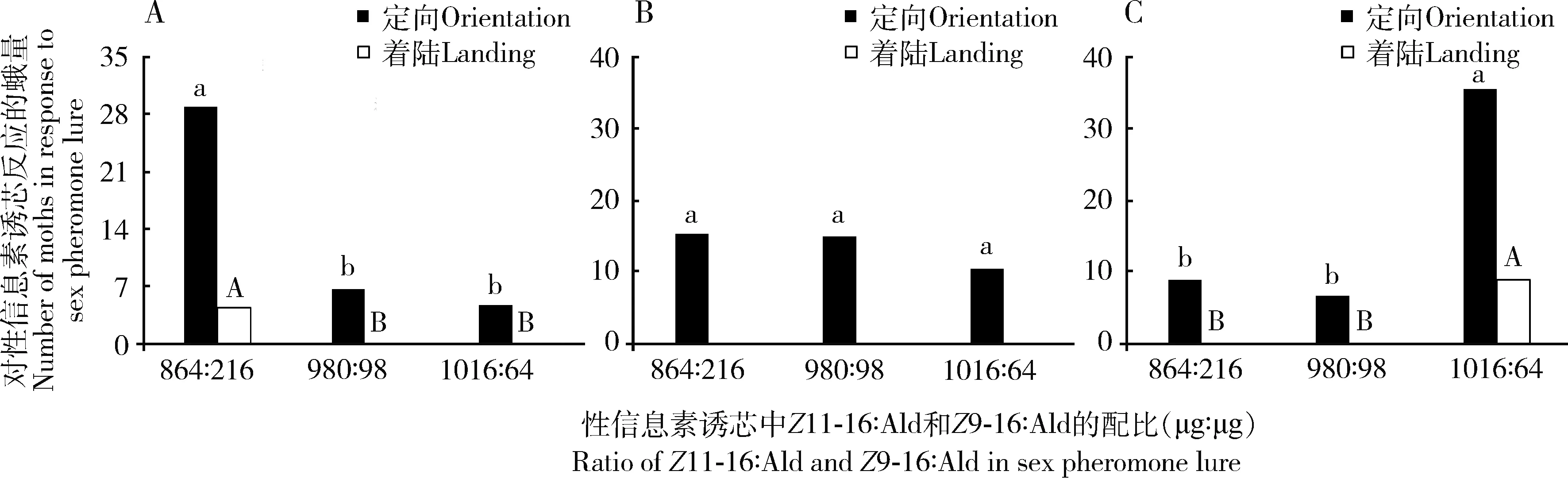
图2 田间性诱越冬代二化螟雄蛾对不同配比性信息素诱芯的风洞行为反应
第2代水稻二化螟进行风洞行为测定时,二化螟雄蛾对各种诱芯的起飞率为92.7%~97.0%。如图3,980 μg∶98 μg配比诱芯诱捕的田间雄蛾对诱芯980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是7.14%, 2.13%和12.82%,着陆到诱芯的比例分别为4.76%, 2.13%和10.26%,不同配比诱芯间差异不显著(P>0.05)(图3:B);864 μg∶216 μg配比诱芯诱捕的田间雄蛾对980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是2.04%, 17.54%和0,着陆到诱芯的比例分别为2.04%, 8.77%和0,864 μg∶216 μg配比诱芯诱捕雄蛾对864 μg∶216 μg配比诱芯的行为反应显著强于对980 μg∶98 μg和1 016 μg∶64 μg配比诱芯(图3: A);配比为1 016 μg∶64 μg诱芯诱捕的田间雄蛾对980 μg∶98 μg, 864 μg∶216 μg和1 016 μg∶64 μg配比诱芯定向飞行的比例分别是1.54%, 4.48%和18.92%,着陆到诱芯的比例分别为1.54%, 0和9.46%,1 016 μg∶64 μg配比诱芯诱捕雄蛾对1 016 μg∶64 μg配比诱芯的行为反应显著强于864 μg∶216 μg和980 μg∶98 μg配比诱芯(图3: C)。
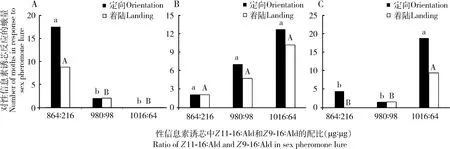
图3 田间性诱第2代二化螟雄蛾对不同配比性信息素诱芯的风洞行为反应
2.3 二化螟信息素受体和信息素气味结合蛋白的定量表达结果
采用RT-qPCR的方法分别比较了水稻种植区Z11-16∶Ald和Z9-16∶Ald不同配比诱芯诱捕的二化螟雄蛾触角中性信息素识别相关基因PBP1-PBP3,PR1-PR6和GOBP1-GOBP2的相对表达水平差异(图4)。发现GOBP1基因在各配比诱芯引诱雄蛾触角中的表达水平无显著差异(P>0.05)(图4: K)。GOBP2基因在864 μg∶216 μg配比诱芯引诱雄蛾触角中的表达水平最高,显著高于除配比1 016 μg∶64 μg外的其余配比诱芯(P<0.05)(图4: L)。PBP3,PBP4,PR5和PR6基因在864 μg∶216 μg, 945 μg∶135 μg和980 μg∶98 μg配比诱芯引诱雄蛾触角中的表达水平显著高于1 003 μg∶77 μg,1 016 μg∶64 μg和1 045 μg∶35 μg配比诱芯(P<0.05)(图4: C, D, I, J)。PR1基因在864 μg∶216 μg, 945 μg∶135 μg和980 μg∶98 μg配比诱芯引诱雄蛾触角中的表达水平显著高于1 003 μg∶77 μg和1 045 μg∶35 μg配比诱芯引诱的雄蛾触角中的(P<0.05)(图4: E)。PR4基因在864 μg∶216 μg, 945 μg∶135 μg和980 μg∶98 μg配比诱芯引诱雄蛾触角中的表达水平显著高于1 045 μg∶35 μg配比诱芯引诱的雄蛾触角中的(P<0.05)(图4: H)。PBP2基因在945 μg∶135 μg配比诱芯引诱雄蛾触角中的表达水平最高,显著高于除配比980 μg∶98 μg外的其余配比诱芯引诱的雄蛾触角中的(P<0.05),在980 μg∶98 μg配比诱芯引诱雄蛾触角中的表达水平显著高于1 016 μg∶64 μg配比诱芯引诱的雄蛾触角中的(P<0.05)(图4: B)。PR2基因在864 μg∶216 μg和980 μg∶98 μg配比诱芯引诱的雄蛾触角中的表达水平显著高于1 003 μg∶77 μg, 1 016 μg∶64 μg和1 045 μg∶35 μg配比诱芯引诱的雄蛾触角中的(P<0.05),在1 016 μg∶64 μg配比诱芯引诱的雄蛾触角中的表达水平显著低于其余配比诱芯引诱的雄蛾触角中的(P<0.05)(图4: F)。PR3基因在980 μg∶98 μg配比诱芯引诱的雄蛾触角中的表达水平显著高于其余配比诱芯引诱的雄蛾触角中的(P<0.05),在864 μg∶216 μg和945 μg∶135 μg配比诱芯引诱的雄蛾触角中的表达水平显著高于1 003 μg∶77 μg, 1 016 μg∶64 μg和1 045 μg∶35 μg配比诱芯引诱的雄蛾触角中的(P<0.05)(图4: G)。PBP1基因在980 μg∶98 μg配比诱芯引诱的雄蛾触角中的表达水平最高,显著高于除配比864 μg∶216 μg外的其余配比诱芯引诱的雄蛾触角中的(P<0.05);在1 016 μg∶64 μg配比诱芯引诱雄蛾触角中的表达水平最低,显著低于除配比945 μg∶135 μg外的其余配比诱芯引诱的雄蛾触角中的(P<0.05);在864 μg∶216 μg配比诱芯引诱的雄蛾触角中的表达水平显著高于1 003 μg∶77 μg配比诱芯引诱的雄蛾触角中的(P<0.05)(图:A)。

图4 性信息素识别相关基因在不同配比诱芯田间诱捕二化螟雄蛾触角中的相对表达量
相关性分析表明,GOBP1,GOBP2,PBP1,PBP2和PR3基因的表达水平与不同性信息素配比之间无显著的线性相关性(P≥0.064),其余性信息素识别基因的表达水平与不同性信息素配比之间均存在显著的线性相关性(r≤-0.847,P≤0.033),其中PR2和PR6的表达水平与不同性信息素配比之间存在极显著的线性相关性(r≤-0.935,P≤0.006)。
3 讨论
利用Z11-16∶Ald和Z9-16∶Ald不同配比的性信息素诱芯在同一田块对二化螟雄蛾进行诱捕比较试验,发现二化螟主要性信息素成分Z11-16∶Ald和Z9-16∶Ald的比例从540 μg∶540 μg至1 045 μg∶35 μg的各混合物均能引诱到雄蛾(图1),表明同一二化螟种群不同个体雄蛾对不同配比性信息素诱芯的嗅觉行为反应存在差异。进一步的风洞行为测定更证实了这种嗅觉差异。将不同配比性信息素诱芯引诱的田间二化螟雄蛾带回实验室进行风洞试验,发现田间864 μg∶216 μg配比性信息素诱芯引诱到的雄蛾对室内864 μg∶216 μg配比诱芯的反应显著高于对980 μg∶98 μg和1 016 μg∶64 μg配比诱芯的反应,田间1 016 μg∶64 μg配比性信息素诱芯引诱的雄蛾对室内1 016 μg∶64 μg配比诱芯的嗅觉反应显著高于对864 μg∶216 μg和980 μg∶98 μg配比诱芯(图2和3),再次证实864 μg∶216 μg和1 016 μg∶64 μg配比性信息素诱芯引诱到的雄蛾性信息素嗅觉反应存在差异,也证实同一二化螟种群中确实存在不同性信息素嗅觉反应的雄蛾。据此,我们可以将嗅觉反应相近的个体理解为一个嗅觉组,同一害虫种群由多个嗅觉组组成。在风洞试验中,部分864 μg∶216 μg配比性信息素引诱的雄蛾也向980 μg∶98 μg和1 016 μg∶64 μg配比性信息素诱芯飞行,部分1 016 μg∶64 μg配比性信息素引诱雄蛾也向980 μg∶98 μg和864 μg∶216 μg配比性信息素诱芯飞行,980 μg∶98 μg配比性信息素诱芯引诱的雄蛾向864 μg∶216 μg, 980 μg∶98 μg和1 016 μg∶64 μg配比性信息素诱芯飞行的比例无差异,表明这些个体的嗅觉谱可能比较广,它们可能可以识别较大范围的性信息素配比。
蛾类昆虫对性信息素的识别过程非常复杂,雌虫释放的性信息素首先通过雄性触角感器上的小孔进入感器中,脂溶性的性信息素通过与性信息素结合蛋白(PBP)形成聚合物的方式进入蛾类昆虫感器内的血淋巴中,进而到达神经细胞树突膜,和性信息素受体结合,刺激嗅觉神经,产生动作电位,将携带的化学信息转变为电信号,之后电信号通过神经突触传入蛾类昆虫脑部触角叶中进行整合,经过整合的电信号被传入蛾类昆虫大脑并产生相应的行为。该过程中,性信息素识别基因的表达水平会影响最终的昆虫行为。本研究的结果也证实了相关嗅觉基因在不同配比性信息素诱捕到的雄蛾触角中的表达量存在显著差异。斜纹夜蛾不同配比合成性信息素诱捕的雄蛾嗅觉受体基因表达水平也存在差异(Linetal., 2015; Zhangetal., 2017)。同时本研究发现,在目前研究的性信息素结合蛋白基因中,GOBP1,GOBP2,PBP1和PBP2的表达水平与性信息素配比之间无显著的线性相关性(P>0.321),而PBP3和PBP4的表达水平与性信息素配比之间均存在显著线性相关(P=0.033)。已有研究表明,PBP2不与二化螟性信息素组分结合,PBP1, GOBP1和GOBP2虽然结合二化螟性信息素组分,但是PBP1还结合其他昆虫性信息素组分(Changetal., 2015; Dongetal., 2019),GOBP1和GOBP2还结合植物气味(Gongetal., 2009; Khuhroetal., 2017),相反PBP3和PBP4与二化螟性信息素组分的结合能力明显强于其他昆虫性信息素组分(Changetal., 2015)。PBP3和PBP4可能才是二化螟触角中结合性信息素的关键蛋白。在目前研究的性信息素受体基因中,PR3基因的表达水平与不同性信息素配比之间无显著线性相关(P=0.064),其余性信息素受体基因的表达水平与不同性信息素配比之间均存在显著线性相关(P<0.023)(图4)。目前已经证实PR3对二化螟性信息素组分无反应,PR1, PR2, PR4和PR6均对二化螟性信息素组分有反应(Changetal., 2015)。有趣的是,有研究证实PR5对二化螟性信息素组分无反应(Changetal., 2015),但是本研究中PR5基因的表达水平与不同性信息素配比之间存在显著线性相关(P=0.023)(图4),这可能是由于二化螟性信息素中还有微量组分未被鉴定或者被测定。
本研究中,每个省份使用的Z11-16∶Ald 和Z9-16∶Ald系列配比中,均有一个配比的诱蛾量高于其余配比,表明利用单一配比性信息素进行群集诱杀时,必须选择诱蛾量最高的配比作为当地的最佳配比。例如在浙江省,1 045 μg∶35 μg配比性信息素诱芯的诱蛾量显著高于其余配比,表明浙江省的二化螟种群以识别1 045 μg∶35 μg配比诱芯的雄蛾数量最多。在利用单一配比性信息素对浙江二化螟进行群集诱杀时,我们需要选择配比为1 045 μg∶35 μg的性信息素诱芯,如果选择540 μg∶540 μg~1 045 μg∶35 μg间的任何一个配比,诱蛾量就明显降低(图1)。但是不同省份二化螟性信息素的最佳配比存在差异,在推广应用单一配比性诱剂进行群集诱杀前,需要对施用地害虫种群的性信息素配比进行筛选,找出针对该种群主要嗅觉组的性信息素配方,才能取得较好的防效。如果只是简单地将其他地方使用的配方搬来使用,可能会出现诱虫量低、防效差等情况。
从本研究结果,以及我们多年大量的、不同种植结构的田间试验示范结果来看,即使选择诱蛾量最高的最佳配比用于性诱防控,也只能诱捕田间雄蛾的一部分。田间剩下的雄蛾由于对最佳配比诱芯反应并不强烈,并不能及时被该诱芯所引诱。因此在田间群集诱杀时,单一最佳配比是不够的。此外,同一种群二化螟个体的性信息素嗅觉反应差异可能给性信息素群集诱杀防治二化螟带来挑战。当利用单一配比性信息素进行害虫群集诱杀时,虽然大量诱杀了对该配比反应强列的二化螟雄蛾,但是随着使用时间的增加,种群中对该配比反应强列雄蛾的数量急剧减少,而对该配比反应弱的雄蛾数量急剧增加,导致该配比性信息素诱虫量急剧减少、防效急剧降低,害虫种群对该配比性信息素快速表现出“抗性”。
为了避免害虫种群对单一配比性信息素快速表现出“抗性”,提高性信息素群集诱杀的防控效果,我们认为可以采用多配比诱芯组合的害虫诱杀新技术,即同时利用多种配比的诱芯组合对二化螟进行组合诱杀。多配比诱芯组合诱杀技术能够对同一害虫种群中不同性信息素嗅觉反应个体进行整体诱杀,不仅拓宽了二化螟性诱剂的引诱谱,增加了性诱防控效果,而且同时压低了害虫种群中不同嗅觉反应组的雄蛾数量,避免了单一配比诱杀导致对该配比无反应雄蛾数量急剧大量增加问题,避免和减缓了害虫种群对性信息素的嗅觉适应性。从2018年在浙江温岭所做的对第3代和第4代二化螟初步田间防治效果的比较试验中,3种不同配比诱芯组合的诱杀方法的防治效果比单一最佳配比诱芯要高出约20%(杜永均, 未发表资料)。另外,多配比组合诱杀技术由于同时使用了多种性信息素配比,可以有效解决性信息素地理种群差异这一应用难题,实现不同地区使用同一套性诱技术进行高效防控。
综上所述,二化螟雄蛾对性信息素的嗅觉行为反应存在地理种群差异,而这种差异不仅表现在诱蛾量最大的最佳配比上,也表现在种群嗅觉组的分布上。这方面的知识有助于性信息素田间应用技术的开发和应用效果提升。
致谢安徽省泾县植保站、吉林省德惠县植保站和辽宁太子河农技中心给予部分田间试验的支持,特此致谢!
