自然光照条件下福寿螺分布规律
2021-06-25陈映僮陈海英丁思敏黄红英
郭 靖,陈映僮,陈 烁,陈海英,丁思敏,黄红英,曾 坚
(韶关学院英东生物与农业学院,广东 韶关 512005)
福寿螺(Pomacea canaliculata),又名大瓶螺、苹果螺,两栖淡水贝类软体动物,原产于南美洲亚马逊河流域。1979年福寿螺作为食用经济动物从阿根廷引入中国台湾省,随即于1981年引入广东省中山市养殖,并在20世纪80年代推广到全国十多个省份,由于其口味不佳、加工成本高等原因,大量福寿螺遭养殖户丢弃,进入到自然水生生境[1,2]。福寿螺适应能力强、繁殖力高,很快在野外建立自然种群,入侵40年来,严重危害中国南方水生作物生产和生态系统功能[1]。福寿螺是广州管圆线虫(Angiostron⁃gyluscantonensis)的重要中间宿主,该寄生虫可能导致人患上嗜酸性脑膜炎[3]。2003年国家环境保护总局和中国科学院[4]将福寿螺列入中国首批16种外来入侵物种名单。
国内外寻求多样化手段控制福寿螺的危害,化学防治往往是农民的首选,稻田撒施杀螺剂的控螺效果较好,且成本较低,但其对水生生物毒性大,破坏水生生态系统正常的物质循环和能量流动[5],且长期使用会导致福寿螺产生耐药性[6]。生物防治因其可持续性强、潜在生态效益和经济效益好等优点而成为研究的热点[7,8],但目前实际推广应用的物种非常有限,主要为常见的数种经济动物,包括鸭子、鲤鱼(Cyprinus carpio)、中华鳖(Pelodiscus sinensis)等[5,9,10],且在应用上存在投入成本高、管理难、风险大等问题。物理防治方法简单、易行、无污染,但往往耗费人力较多,常作为辅助措施使用[8]。寻求高效生态的控制手段,仍是当前福寿螺相关研究的主要工作之一。而探明福寿螺的生活习性,对其精准化、高效化防治具有重要的现实意义。
福寿螺一般生活在水中,只有在产卵时爬出水面,具有遇旱休眠、遇水复苏的特性[11]。前人据此采用水稻旱直播、水旱轮作等种植方式减少福寿螺对水稻的危害[12,13]。Wada等[12]研究发现水稻隔代改种大豆能够在一定程度上减少福寿螺对水稻的危害。国内亦有研究证实稻田冬种马铃薯能有效减少来年早稻田福寿螺的发生量[14]。前人亦根据福寿螺的食性探究对福寿螺有趋避作用的植物资源[15],通过诱集或驱赶福寿螺,以减少福寿螺对水稻等水生作物的危害[16]。储少媛等[17]通过插竿位置、间距、高度、形状4个因素探究福寿螺的产卵习性,确定了不同插竿方式对螺卵的诱集效率,节省人力。此外,在水稻生长前期,福寿螺往往将卵产在田埂上,此时期消除螺卵最为高效易行[14]。
有关福寿螺在水中的生活习性研究较为少见,光仅作为一种复杂的生态因子,光照强度、光谱成分及光周期性变化直接或间接地对福寿螺的摄食、生长和存活等产生影响[18-20],但至今鲜见自然光照条件下福寿螺分布规律的研究报道。本研究选在晴天白天,调查了不同时间福寿螺在光照区和遮阴区的分布情况,揭示福寿螺在自然光照条件下的分布规律,为进一步探索福寿螺的生态适应性和精准防控提供基础数据和科学依据。
1 材料与方法
1.1 材料
试验用螺于2019年9月23日和25日采集于韶关市浈江区黄屋村稻田(24°78′N,113°66′E),置于实验室楼顶,以塑料水族箱(72 cm×56 cm×38.5 cm)进行养殖,水深约20 cm,以新鲜生菜为食物每天投喂。驯养数天后,挑选表观正常、活力较强的福寿螺个体用于试验。
1.2 研究方法
2019年9月27 日,选用9个塑料水族箱(55 cm×45 cm×35 cm),灌入自来水,分成5、10、20 cm 3种不同深度水层,各水层均设3次重复。次日,各箱引入10只成年福寿螺(壳高25~40 mm)和10只幼龄个体(壳高14~25 mm)。每天9:30、12:00、14:30测定水体温度,将温度计置于光照区域水下约5 cm处,精确到0.5℃;用直尺测量塑料箱底部光照区域面积,遮阴区面积通过塑料箱底面积减去光照区面积获得。同时,对福寿螺的分布进行观察,确定光照区和遮阴区的成、幼螺数量,计算光照区和遮阴区的成、幼螺密度。如遇阴天,则不进行观察。试验持续到10月6日,期间共观测7 d,10月2—3日未观测,10月6日9:30时处于阴天状态,未进行观测。
试验期间每天傍晚投喂少量新鲜生菜,确保当晚能吃完。每2 d(傍晚)换一次水,换水量约为原有水量的3/4,保证水体较为清澈。除喂食及换水,箱表面一直覆盖纱网,以防止螺逃逸。若在观测或换水时,发现死螺,及时捞出,试验期间共死亡5只螺。为避免死螺对不同处理间福寿螺密度的影响,依据光照区和遮阴区的成螺(或幼螺)比例,进行等比例数据转换,使得各箱中成螺和幼螺的数量在数据分析时均为10只。
1.3 数据分析
运用Microsoft Excel软件处理数据,采用SPSS 20.0统计软件进行分析,对同一水深同一时间光照区和遮阴区的福寿螺(未区分成螺和幼螺)分布密度进行正态性检验,如服从正态分布,则进行t检验,否则进行U检验;对同一水深同日不同时间福寿螺(未区分成螺和幼螺)分布密度差(遮阴区螺密度-光照区螺密度)进行正态性检验,如服从正态分布,则进行Duncan’s多重比较,否则进行Kruskal-Wallis非参数检验;对不同水深、不同时间的水温进行重复测量方差分析,并对同一因素不同水平进行Duncan’s多重比较,如水温不服从正态分布,则进行Kruskal-Wallis非参数检验,并用Nemenyi检验进行两两比较;对不同大小福寿螺在是否有光照、不同水深、不同时间的分布密度进行重复测量方差分析。整理绘图采用Origin 8.0软件进行。
2 结果与分析
2.1 福寿螺所处环境温度变化
由图1可知,每天9:30—14:30水温均表现出不断升高的趋势,在观测的7 d内,9:30水温基本保持在25~30℃,除5 d时,5 cm水深水族箱的水温显著高于10 cm和20 cm水深,其余各天9:30不同水深间水温差异不明显。12:00不同水深处理的水温均升高,同时三者水温差异增大,调查的7 d内,12:00时5 cm水深处理下水温均显著高于20 cm水深处理(P<0.05),两者温差此时最大,10 cm水深处理下的水温居于5 cm和20 cm水深中间。14:30不同水深处理的水温均继续升高,水温均在35℃以上,甚至个别时候水温高达40℃,部分时候出现10 cm水深处理下水温最高的情况,但三者水温差异减小。
重复测量方差分析(因9:30无光照,10月6日数据剔除)发现,不同日、同日不同时间、不同水深均对水温有极显著的影响(P<0.01),通过两两比较发现,就整体而言,不同水深水温从高到低依次是5、10、20 cm,两两之间达到显著差异(P<0.05);不同时间水温从高到低依次是14:30、12:00、9:30,同样地,两两之间差异达到显著水平(P<0.05)。
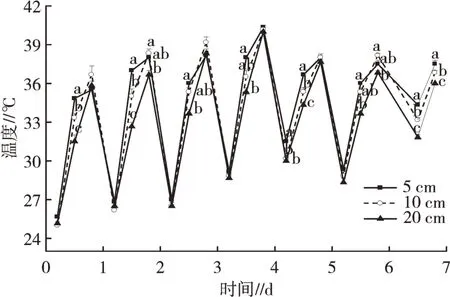
图1 福寿螺所处的环境温度
2.2 自然光照对福寿螺分布节律的影响
由图2可知,调查的各天9:30福寿螺多表现出在遮阴区和光照区密度相近,少数时候两区域间达到显著差异,总体来看,规律不明显。随着时间的推移,遮阴区福寿螺密度逐渐增加,光照区福寿螺密度逐渐降低,在调查的各天中午(12:00),不管水深为5、10 cm或是20 cm,福寿螺在光照区域的密度均显著小于遮阴区域。各天12:00—14:30,遮阴区福寿螺密度基本上呈现出不同程度的下降,光照区福寿螺密度变化不一,但均低于遮阴区,近50%调查时间点两者间差异达到显著水平,主要体现在水深5 cm和10 cm的环境条件下。
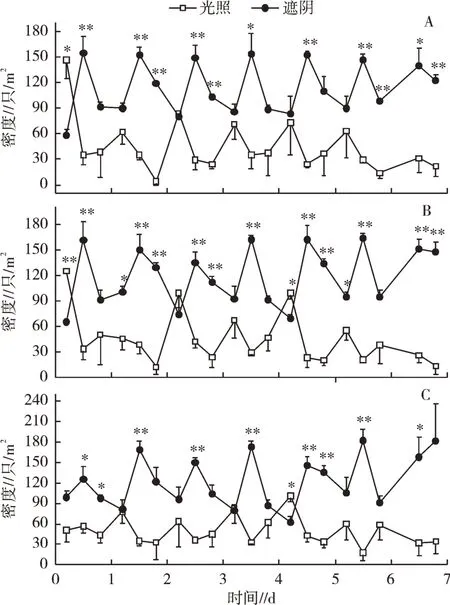
图2 自然光照条件下福寿螺分布情况
由图3可见,在水深5 cm条件下,调查的前3 d发现9:30福寿螺分布密度差(遮阴区螺密度-光照区螺密度)显著低于12:00和14:30的分布密度差,而整个调查过程中,12:00和14:30福寿螺分布密度差差异均未达到显著水平。而在10 cm和20 cm水深条件下,在调查的4 d和6 d,12:00福寿螺分布密度差显著高于14:30的分布密度差,其余各天两时间点间差异不显著。除此之外,在调查的前6 d,水深10 cm和20 cm情况下,12:00福寿螺分布密度差多显著高于9:30的分布密度差,而9:30和14:30两时间点之间的福寿螺分布密度差多未表现出明显差异。
由于重复测量数据不满足球形假设检验,单因素方差分析结果表明,处理时间对福寿螺的分布没有显著影响。重复测量方差分析结果(表1)显示,光照和同日内不同时间点及其两者交互作用对福寿螺的分布均存在极显著的影响,其余因素对福寿螺的分布影响均未达到显著水平。

图3 同日不同时间点的福寿螺分布密度差
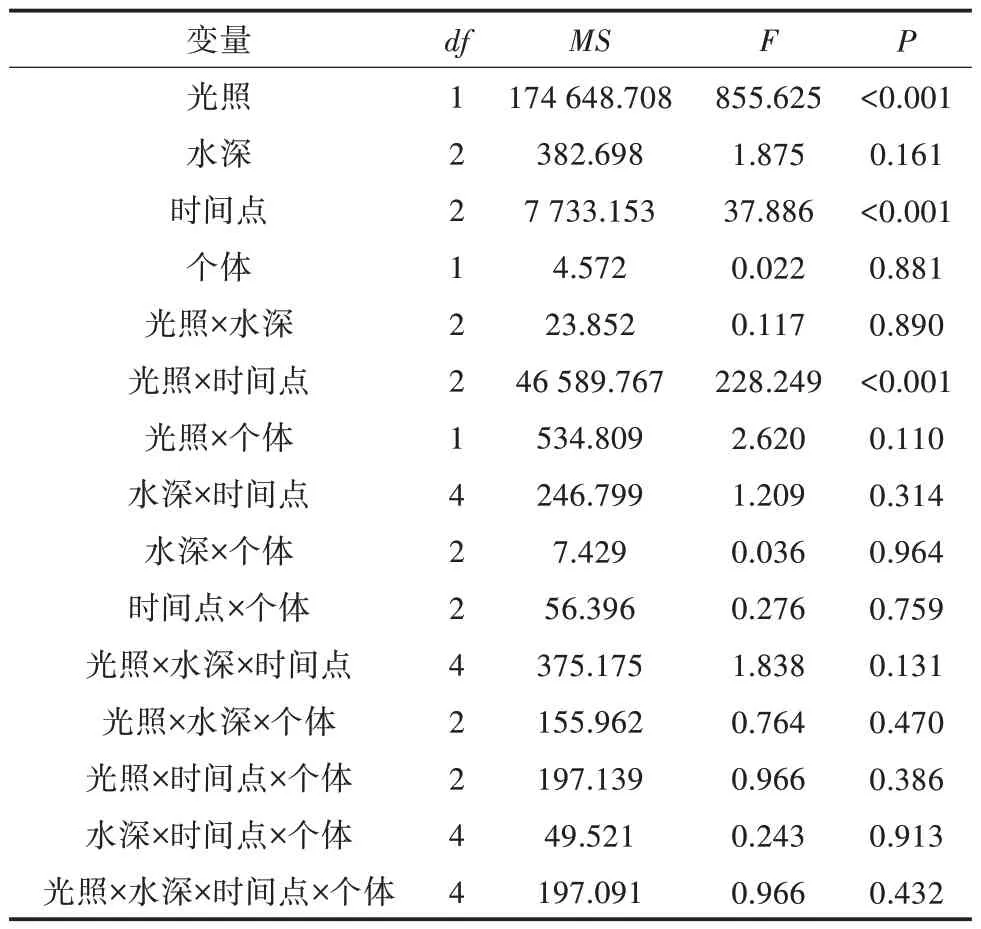
表1 福寿螺分布的重复测量方差分析
3 小结与讨论
光作为重要的生态环境因子,通过光质、光强和光周期等方面对福寿螺产生直接或间接的影响。本研究发现自然光照会影响福寿螺在水中的分布,福寿螺在光照区和遮阴区的分布密度呈现出日变化规律。福寿螺在弱光条件下取食量相对较大,22:00至次日6:00摄食旺盛,活动能力更强,白天取食较少,尤其是12:00—15:00几乎不取食,此时福寿螺会聚集在水中植物叶片下方,避免强光直射[18]。本研究亦发现在阳光直射下,福寿螺亦往遮阴处移动,原因可能包括但不限于以下3个方面:一是与水温有关,本研究中12:00和14:30水温均在30℃以上,甚至高达40℃,而水温超过30℃时,福寿螺的存活率、摄食率、生长率均随着温度的升高而降低[21,22],爬至遮阴处有利于福寿螺躲避高温带来的不适;二是与光照强度有关,本研究中14:30水温最高,但12:00遮阴区和光照区螺密度之差往往更大,这可能与福寿螺对光照反应较敏感有关[20],晴天正午时光照度可能超过80 000 lx[23],有研究发现福寿螺在光照度超过30 000 lx时即会逃避[18];三是福寿螺的天敌多为昼行性天敌[8],爬至隐蔽处或聚集在叶片下方,减少其暴露而被天敌捕食的机会。
除了光照强度以外,光质和光周期亦会对福寿螺造成一定影响。光质对福寿螺有趋避效应,且趋避效果随光质和螺壳颜色变化而变化。红色光对黄壳和黑壳螺均有明显的驱避效果,而黄色光对黄壳螺有诱集作用,绿光和蓝光对黑壳螺的诱集效果较好[19]。已有研究发现,黄光有利于水稻生长初期株高增长,红蓝组合光有利于培育水稻壮苗[24],故可考虑在稻田构建红蓝光这种“诱集”与“趋避”相结合的推拉系统,同步实现控制螺害和促进水稻幼苗生长,这还有待于进一步研究。不同光质亦能不同程度地影响福寿螺假死持续时间、生长速率和趋食性[25]。此外,光周期对福寿螺的存活、生长发育和繁殖均能造成一定影响,连续长时间(30 d)光照和黑暗条件下,福寿螺存活会受到一定抑制[18]。朱丽霞等[20]研究发现在全光照条件(3 000 lx)下,福寿螺摄食较多,长时间处于活动状态,而光照会干扰福寿螺的产卵行为,导致产卵较少,进而使其体重增长较大;福寿螺在完全无光条件下,几乎不摄食,繁殖能力受抑制,同样不适于福寿螺种群发展;每日12 h光照会使得福寿螺产卵量和孵化率均较高,且孵化历期最短,由此可见,合适的日夜变换有利于福寿螺的种群发展。
本研究发现,福寿螺会表现出躲避强光的行为,水温可能是重要影响因素,下一步拟在冬春季节开展类似研究工作,以揭示不同季节强光对福寿螺分布节律的影响。值得注意的是,福寿螺繁殖盛期往往处于夏秋季节,高温条件下福寿螺的避光行为有利于福寿螺的生态防控,如农民可重点捞取遮阴区的福寿螺,或在遮阴区投放废弃瓜果蔬菜,或人为创造遮阴区等来提高诱集效率(12:00—14:30效果较好)。研究发现成螺和幼螺在自然光照条件下的分布节律未表现出明显差异,有利于对不同年龄期的福寿螺进行控制。水深会影响水温,但就本研究而言,不同水深带来的水温差异不足以显著改变光照对福寿螺分布的影响。此外,国内外尚未见报道光照在夜间条件下对福寿螺的趋避效果,且不清楚趋避效果是否会受光照强度和光质的影响,该方面的探究可进一步揭示光照条件下福寿螺分布节律,有助于建立福寿螺的光防治体系。福寿螺的生态防控需多手段相结合,才能将螺害降到最低。
