盐沼潮滩生物动力地貌演变研究进展
2021-06-08林伟波张长宽
周 曾,陈 雷,林伟波,罗 锋,陈 雪,张长宽
(1. 河海大学水文水资源与水利工程科学国家重点实验室,江苏 南京 210098;2. 河海大学江苏省海岸海洋资源开发与环境安全重点实验室,江苏 南京 210098;3. 江苏省海涂研究中心,江苏 南京 210036)
潮滩是海岸带的重要组成部分,是海陆营力相互作用的前沿地带,在保护海岸湿地环境资源、维系海洋生物多样性、提高海岸防护能力、固存蓝碳等方面发挥着重要的作用。在国家海洋生态文明建设、蓝色海湾整治和海岸带保护修复行动等背景下,滨海潮滩湿地保护与修复日益成为各级政府和学术界的关注焦点。潮滩中上部多分布着耐盐植被,如盐沼、红树林等,构成潮滩湿地系统,在温带和亚热带地区主要以周期性被潮水淹没的草本植物盐沼分布最为广泛,其多发育于潟湖、海湾等半封闭的低能海岸或细颗粒泥沙供给丰富的河口三角洲、平原海岸等。盐沼潮滩湿地是生物生产力最丰富、蓝碳固存最高效的生态系统之一,世界范围盐沼植物优势种有米草属、盐角草属、盐草属和灯心草属等。中国沿海的盐沼分布十分广泛,常见于辽东半岛东部、渤海湾、江苏、上海、浙江、福建等地[1- 2],本地原生物种有芦苇、盐地碱蓬、藨草、海三棱藨草等,引入的外来物种主要有大米草、互花米草,其中互花米草至1979年引种以来迅速在中国沿海潮滩蔓延,已成为中国淤泥质海岸潮滩的主要物种之一,其引入的利弊分析一直是讨论的热点[1]。
从近年来发表在Nature或Science等顶级期刊上的国际引领性成果来看,河口海岸潮滩系统动力地貌学的研究已从传统的水- 沙- 地貌三者的相互作用逐渐过渡到考虑全球变化背景下的耦合机制[3- 5],具有多学科交叉、融合的特点,涉及包括海岸动力学、地貌学、沉积学、环境生态学、生物学等多个学科,对这一区域的研究已逐渐从传统的单一学科视角过渡到多学科的交叉和融合,形成了一些新的学科发展方向,其中“生物动力地貌学”(Biogeomorphology)自Viles于1988年提出以来得到了国际学术界的高度关注,近30 a来发展尤为迅速[6- 7],生物动力地貌学的研究将生物学、生态学的知识用于解释和分析地貌演化规律,极具创新性和挑战性。就河口海岸这一研究区域来说,潮滩生物动力地貌学在传统的水- 沙- 地貌关系基础上,进一步考虑海岸带动物(如贝类、鱼类、鸟类等)[8]、微生物(藻类、菌类等)[9]及耐盐植被(如盐沼、红树林等)[10]对水- 沙- 地貌的作用以及水- 沙- 地貌对生物的反作用。
作为潮滩湿地分布最广泛的生物类型之一,盐沼植被由于其生物特性引起的如弱流、消浪、固沙等生物- 物理效应(Biophysical effects)对于潮滩地貌演变有着极其重要的影响,而潮滩水动力、泥沙动力等因子对于植被的生长、消亡也起着至关重要的作用。此外,盐沼植被为大量野生动物、底栖动物以及微生物提供了宝贵的栖息地,构成了一个重要的湿地生态系统,各类共生生物之间也会相互影响和作用。尽管中国一些学者在20世纪八九十年代开始意识到盐沼植被与地貌之间的双向动力反馈机制,指出研究潮滩地貌演变与盐沼植被区域变化是不可割裂的[11- 14],但是限于过去认知水平和观测、计算能力,中国在本领域开展系统的科学研究时间还不长。在面对全球气候变化、海岸城市防洪以及海岸侵蚀的压力下,合理应用盐沼植被进行海岸生态防护或人工岸线生态化已成为很多发达国家的首选[15- 16],这也是中国“十四五”期间在海岸带保护修复领域的重大需求,因此,迫切需要加深对盐沼潮滩生物动力地貌演变机制的认知,突破其模拟预测技术,本文旨在综述盐沼潮滩动力地貌研究领域的研究进展,并探讨本学科方向未来有待进一步深入研究的科学问题。
1 盐沼潮滩地貌特征概述
盐沼潮滩中上部分布着耐盐植被,中下部一般为光滩,盐沼滩与光滩的交界处通常被称为盐沼前缘。陆侧成熟的盐沼通常是呈片状分布,且常伴随潮沟系统来实现物质输运,总体而言成熟的片状盐沼潮滩地貌变化缓慢,而其地貌演变通常以其前缘与光滩交界处的变化(如盐沼植被的海向推进与陆向蚀退)最为显著且相对易于测量,同时,由于植被、动力与地貌演变之间的反馈机制,盐沼前缘形态的变化通常可表征潮滩水动力与沉积环境特征,具有重要的科研与应用价值。
盐沼前缘总体呈现3种典型的地貌形态[17- 18],如图1所示。一是盐沼滩与光滩之间以盐沼簇团(即盐沼植物在潮滩上以簇团或斑块状的集中分布)的形式平缓过渡,受局部水动力、沉积物供给等因素的影响,盐沼簇团大小、形态各异,靠海侧簇团一般较小,靠岸侧簇团随着盐沼扩张可连成片[19- 22];二是盐沼滩与光滩之间表现为高度约0.5~2.0 m的前缘陡坎,这类陡坎形态一般被认为是侵蚀型潮滩的表征[23- 27],但也有学者发现前缘陡坎也可发育在淤涨型潮滩环境[28- 29];三是介于前两者之间的脊- 沟相间的地貌形态,常被认为是过渡性地貌单元[17,30],长期来看主要表现出侵蚀性特征,脊- 沟走向通常与岸线垂直并沿着海向呈现一般小于10°的坡度,靠海侧的脊- 沟上一般无盐沼植被分布。

图1 不同盐沼潮滩地貌前缘形态Fig.1 Tidal flat- marsh marginal morphologies
盐沼潮滩的地貌特征受控于很多因素,包括泥沙供给情况、海平面上升速率、区域水动力环境、生物种群竞争与共生行为等等。一般而言,盐沼簇团主要分布于波浪较弱、沉积物来源丰富的淤涨型潮滩,在中国江苏沿海潮滩与长江三角洲潮滩较为常见;盐沼前缘陡坎常见于波浪较强、沉积物供给较少的侵蚀型海岸,在英国东部海岸、荷兰瓦登海以及意大利威尼斯潟湖报道较多;脊- 沟相间的前缘地貌形态的研究目前文献报道相对较少,主要集中在英国的一些潮滩。这3类盐沼前缘形态会随着环境条件的改变发生系统状态转换,且一些地区的盐沼前缘可表现出周期性的侵蚀与扩张特征[17,29,31],对于潮滩湿地系统的影响巨大,但其系统状态转换机制尚不完全明确,例如,江苏盐城丹顶鹤自然保护区的潮滩盐沼前缘在短短5 a时间内从平缓型簇团前缘转化为侵蚀型陡坎[27]。
盐沼潮滩呈现上述地貌特征以及演化过程是盐沼植被与水- 沙- 地貌等多种动力因子相互作用的结果,近30 a来,学界在盐沼植被的弱流、消浪、截沙、固沙等生物作用以及潮滩动力过程反作用盐沼植被机制上取得了一系列成果,进而也推动了盐沼潮滩地貌演变数学模型的快速发展。
2 盐沼对潮滩水- 沙- 地貌过程的影响
盐沼植被因为拥有能改变生存环境使之更加适合自身发展的能力而被类比为“生态系统工程师”[32],通过其根、茎、叶来消减水动力、拦截泥沙、固定泥沙等生物- 物理效应,可以有效地促进淤积、提高床面高程,进而改善其生存环境。
2.1 盐沼植被对水动力的作用
潮滩常规水动力主要是潮流和波浪,现场观测和室内实验均表明盐沼有着显著的弱流与消浪作用。当潮流传播到盐沼区域时,盐沼植被就会对潮流产生阻挡,其阻挡水流的表现为平均流速降低、紊动产生、能量耗散以及垂向流速剖面的改变[33]。杨世伦等通过现场试验发现,植被带中流速在任何时刻均小于相邻的同高程光滩或人工刈割裸地区域,且相对应的植被缓流系数(盐沼区域流速与光滩流速之比)与植被覆盖率以及测量点距盐沼前缘距离呈正相关[13,34]。王爱军等[35]在泉州湾互花米草盐沼的观测同样表明,互花米草盐沼内的流速明显小于光滩地区,而且随着互花米草宽度、高度及植株直径等要素的增加,流速减小愈发显著。但当植被高度、宽度和直径增大到一定程度后,流速变化就基本不受以上要素的影响了。时钟等[11]在长江口南汇东滩进行现场水文泥沙观测取样发现,海岸盐沼植被的存在使近底层流速减弱,流速的减弱量值与水流在盐沼中流过的距离成正相关,同时与植物参数有密切关系,且盐沼区内底层流速在涨潮时刻大于落潮时刻,呈现不对称性。此外,盐沼冠层以上水流流速明显大于冠层以下流速,即植被缓流效应在潮流淹没植被冠层前后大不相同,且会改变潮流的垂向流速分布结构[33,36- 38]。
波浪在光滩上传播时,由于水深减小以及底摩阻增大,波浪开始变形甚至破碎,波能会逐渐耗散;而在盐沼区域内,波能衰减以波浪破碎为主要耗能方式,同时植物的地上部分与海水和水中沉积物直接接触,水体与盐沼植株茎叶发生强烈摩擦,植物茎叶会减缓流速,减少湍流,促进沉积物沉积,同时地下根系通过稳定土壤基质直接减缓侵蚀速率[11,39- 41]。Möller和Spencer[42]在英国Essex海岸的观测发现:Tillingham泥滩上,平均波高在147m内降低了20.57%,而在Tillingham盐沼内,平均波高在163m内降低了87.37%;在Bridgewick泥滩上,平均波高在102m内升高了20.57%,而在Bridgewick盐沼内,平均波高仅在10 m内就降低了43.81%,研究还发现大部分的波能衰减发生在盐沼前缘的几米处。史本伟等[39]在长江口崇明东滩观测也发现波高、波能密度在淤泥质潮间带盐沼上的衰减率比在光滩上高一个数量级,且其效能效果受到植被种类、高度、密度等因素影响。然而,值得注意的是,当水深淹没盐沼植被冠层顶部后,波浪衰减会明显减弱[2,13,43]。
2.2 盐沼植被对泥沙输运过程的影响
盐沼对泥沙输运的影响分为直接和间接影响两方面[1,11]。潮滩泥沙运动主要是由潮流、波浪、风暴潮[44]等水动力条件驱动引起,水动力的强弱决定了泥沙输运的多少。直接影响是指盐沼植株本身对于水体中悬沙有着显著的黏附作用,可以直接捕获水体中的泥沙颗粒。Mudd等[45]结合实验室研究和18a的现场植被特征数据记录发现盐沼植被的泥沙捕获量强烈依赖于盐沼内的水流流速,在大流速情况下(流速>0.4 m/s),捕获的泥沙颗粒占涨潮输运泥沙总量70%以上;在一般流速情况下(流速<0.1 m/s),这个比例小于10%。此外,泥沙捕获量还与植被特性、水体悬沙浓度和底床高程(决定植被淹水情况)相关。Li和Yang[46]在长江三角洲的实地测量发现,不同植被部位的泥沙捕获效率不同(茎叶结合部以及果实处的沉积物密度是茎、叶处的5~10倍);泥沙捕获量随植被生物量增大而增大;随着距离盐沼前缘或潮沟越远,捕获量越小。但Reef等[47]通过现场水槽试验也发现,一个潮周期内的泥沙捕获率与植被冠层形态无明显关系,且在一般动力条件下泥沙捕获率不受植被高度或生物量的影响。
此外,盐沼通过弱流、消浪等作用对泥沙输运也有重要的间接影响,盐沼植被通过消浪、弱流作用减弱了盐沼区内水流的挟沙能力且一定程度上抑制了底床悬沙质的再悬浮,同时也促进了泥沙沉降和滩面物质堆积。已有大量现场观测数据证实,盐沼植被滩内的水体悬沙浓度要明显低于没有植被的光滩区域[34,48- 49]。
2.3 盐沼植被对潮滩冲淤变化的影响
盐沼通过改变水动力环境以及悬沙输运,会对潮滩的堆积格局、地貌演化和稳定性产生影响。由于植被的缓流以及黏附作用,会使细颗粒悬移质沉降在盐沼区,从而使得盐沼区的滩面沉积物细化。陈一宁等[51]对比了江苏王港潮滩的盐蒿滩、大米草滩与互花米草滩的滩面表层泥沙粒径,发现互花米草滩的粒径最细,盐蒿滩最粗,而互花米草滩高程最低最为靠海,因此,互花米草促进了细颗粒物的堆积且改变了潮滩原有的泥沙堆积格局(岸向细化[50])。陈才俊[52]研究发现,大米草在江苏淤长型海岸可以增加2倍左右的淤积量,微淤和弱侵蚀海岸在种植大米草以后可以转为缓慢淤长,但大米草只能促进大米草带内的淤积。
潮滩环境中浅土层区并非完全密实,季节更替、潮水涨落、地下水位变化、盐沼植物的生长等都会导致土层的膨胀或收缩。秋冬季露滩时间较长,土层会因排水发生固结压缩,滩面高程降低,而春季大潮期间海水入侵滩面会使土体膨胀,从而导致滩面高程抬高。盐沼植被的地上部分通过改变水流结构,促进水体中悬浮颗粒的沉降,地下部分根系的生长与分解作用能增加地下土壤有机碳的储备,随着营养物的增加,盐沼植被的根系累积速率会发生改变,通过向地下膨胀从而有利于加快盐沼区高程的增加[53- 54]。表层泥沙的淤积同时也促进地下根系的生长,扩大根系主要吸水区域的土壤深度,促进土壤淤积层水分的消耗,加快土壤表层水与地表水之间的水分循环,改善土壤中的营养条件,有利于盐沼植物的生长和扩张[55- 56]。氧气在孔隙水中的扩散速率和浓度较低,植物生长与土壤通气性密切相关,植物的蒸腾作用能够加快地下水分的消耗,进而提高了植物根系周围的氧化条件[57- 58]。盐沼也会影响潮滩潮沟的演变,一般来说会增加其稳定性。以互花米草盐沼为例,由于互花米草植株高大、群落盖度高,滩面落潮水归潮深度变大,有更多的滩面水参与潮沟的塑造,因而互花米草盐沼潮沟密度大;互花米草根系发达,滩面比较“结实”,耐冲刷,使互花米草盐沼潮沟宽深比较小(一般小于8),同时也使潮沟弯道处的弧度较好,难以裁弯取直;互花米草对潮水有较强的阻挡作用,潮沟中潮水漫滩时,流速迅速降低,泥沙大量沉积,使互花米草盐沼潮沟形成明显的沿岸堤;在潮流由光滩进入互花米草盐沼时,互花米草的阻挡产生扰动,形成横轴环流,掏蚀盐沼边缘泥沙,在互花米草盐沼外侧形成明显的边缘冲沟式潮沟[59- 60]。
3 盐沼植被演替对动力过程的响应
盐沼潮滩是一个复杂的生态系统,盐沼植被作为其中的重要组成部分,在其影响水动力、泥沙输运及地貌演变的同时,盐沼本身的繁衍生长也受到环境因素的影响。河口海岸潮滩这种海陆交界的特殊环境和多种动力过程是影响盐沼植被空间分布与演替的主要因素(图2)。从世界多国的研究来看,盐沼植物群落的分布通常呈现一定的分带性特征,与淹水时间、盐度、营养供给、高程、水动力条件等环境梯度相关,这些通常被归类为物理因子,相关研究十分丰富;近年来,很多研究发现群落中动物、微生物、植物等生物因子的相互作用对盐沼植被群落的演替及稳定性具有重要的影响,成为研究的热点之一,研究通常认为物理环境因子与生物因子的相互作用塑造了盐沼群落分布特征、决定了其演替机制与过程[61]。此外,人类活动对盐沼植被分布及演替有着深刻的影响,主要体现在沿海水产养殖、海岸工程、生物入侵、海岸环境污染、气候变化等方面。以下从物理因子、生物因子以及人类活动等3个方面简要介绍盐沼植被演替方面的相关进展。

图2 盐沼潮滩特征与动力过程示意Fig.2 Characteristics and processes of tidal flats and saltmarshes
3.1 物理因子
传统景观生态学、海岸地貌学等研究领域在早期重点关注了物理因子的影响。在大尺度层面,气候和温度对盐沼植被的分布具有重要的影响,决定了盐沼生境的大致纬度;在具体河口海岸区域小尺度层面,包括潮流、波浪、风暴潮、泥沙供给等因子具有较强的影响。
潮流很大程度上影响盐沼的生长与扩张,潮汐淹水通过对土壤透气性和化学特征的改变而影响盐沼植物生长,尤其是处于生长阶段的植株,淹水频率过低会造成干旱高盐,而淹水频率过高则会造成缺氧,最后都会导致植物死亡,因此,盐沼植物生长需要一个适宜的高程条件[62- 64],此外,潮流也是输送盐沼种子使之在滩面扩散的主要原因。泥沙堆积也可促进盐沼的扩张,汪亚平等[12]根据琼港盐沼资料分析得出,小潮时,水流主要集中在潮沟中,只淹没盐沼边缘,此处频繁的潮流作用使盐沼在形成初期演化迅速;大潮时,水流可浸漫盐沼滩的大部分区域,水流溢出潮沟、漫过盐沼表面时产生流速突变,将泥沙带往盐沼面,由于延迟效应和盐沼植物的阻滞作用而堆积下来,进而进一步促进了盐沼的繁衍。波浪与风暴潮可对盐沼植被产生一定的侵蚀破坏,盐沼前缘陡坎侵蚀和崩塌的主要因素就是波浪作用,波浪对盐沼边缘作用力强弱取决于潮位情况,波浪作用力在波高一定时随着潮位上升而增大,增大到盐沼边缘滩面恰好被淹没时,之后波浪力呈现迅速减小的趋势[65]。
此外,海平面上升这种长时间尺度的动力过程也具有较大影响。如果泥沙供给引起的滩面高程提高速率小于海平面上升的速率,在造成潮滩面积减少的同时,也会引起盐沼的退化,形成光滩[66];如果泥沙供给充足,能够抵消甚至超越海平面上升带来的影响,会引起盐沼垂向沉积,促进盐沼海向扩张[12]。值得一提的是,潮滩地貌特征也对盐沼演替具有一定影响,潮沟是潮滩上常见的一种地貌单元,是盐沼区内外水体与营养物质交换的重要通道,潮沟的发育演变将影响盐沼区植被的生长与扩张。侯明行等[60]在盐城保护区通过实地调查与遥感影像相结合,分析发现潮沟的发育是影响米草向内陆碱蓬区扩张的因素之一,而米草陆向扩张的宽度和幅度取决于所在区域潮沟的发育程度;王青等[67]在黄河三角洲滨海盐沼的控制实验发现,盐地碱蓬的种子捕捉量、萌发率和幼苗存活率均与距潮沟的距离有关,随着远离潮沟区域,植物定植率降低。
3.2 生物因子
近年来,生物因子的影响也得到了学术界的重视,试图从生态系统内部物种间的相互促进、制约关系来揭示盐沼植物群落分布的生态学机制。不同种类植物的竞争与共生是盐沼植被演替的重要影响因素,以海三棱藨草为例,由于外来物种互花米草生长期长,植株高大,互花米草入侵海三棱藨草后,会形成较为密闭的空间环境,抑制海三棱藨草的生长[68]。在中国东南部沿海,互花米草盐沼和红树林间的竞争问题严重[69]。无论就纬度分布、潮间带的垂直分布还是沿河口上溯的分布而言,红树林和互花米草的生存环境都存在重叠,同时互花米草具有更广的盐度适应范围、更强的耐盐水能力和繁殖扩张能力,更具有竞争优势[70]。但有研究通过遥感图像分析得到,互花米草仅减缓了红树林向外扩张速率,并没有明显入侵到红树林之内,因此在一定条件下互花米草和红树林可以共存[69]。
盐沼湿地系统的演化是在水动力、沉积过程和生物过程之间相互作用和调整的基础上产生的[71- 72]。底栖生物的变化改变了水动力和沉积过程,影响了盐沼湿地系统内潮沟、盐盘、陡坎等地貌的发育[73],同时地貌的改变反作用于潮滩生物的分布。潮沟的形成始于潮滩表面潮汐通量的集中,产生的局部冲刷应力超过泥沙临界起动切应力。植物的斑块的存在、蟹类等底栖动物对植物斑块的扰动或地形的小扰动都会对潮流产生阻力,从而导致潮汐通量的集中;底栖生物在摄食、建造维持洞穴等过程中破坏了沉积物的原生结构,降低了土体的强度[74];洞穴- 地下水的交互作用也能促进潮沟的形成,退潮后被截留在洞穴内的地下水,由于蟹类侧向洞穴的存在,会加速从高水头向低水头流动。地下水常沿潮沟堤岸渗透,但如果存在贯穿洞穴,则会集中渗出,从而侵蚀松软的沉积物。同时,由于鱼类的捕食作用,洞穴开口会进一步增大,使得正在发育的潮沟不断加宽加深,进一步促进潮沟的形成和演化[75]。盐生植物多以圆形或椭圆形分布[76],植物的存在能够较好地缓解底栖动物的生存压力,底栖动物通过挖掘洞穴,改变沉积物结构的稳定性,加速了盐沼湿地的侵蚀,导致中心区域的植物死亡,盐生植物远离中心向外生长,底栖动物随之向外迁移,从而形成一个不断生长的圆环[77]。盐沼湿地内部陡坎的形成,始于早期盐沼湿地边缘植物- 沉积物- 水动力的反馈作用。植物的存在促进沉积物的累积,导致植被带边缘水动力作用加强,最终形成陡坎[78]。植物的存在可以稳定陡坎,蟹类等底栖动物的存在促进陡坎的崩塌。当生物量较高时,植被的稳定作用大于蟹类的解构作用,陡坎基本保持稳定;当植被生物量相对减少,蟹类对陡坎的破坏,促进陡坎上部植被的崩落前移。若水动力作用较弱,崩落的植被在陡坎前方定植,有益于陡坎的向海前进;反之,则导致陡坎的后退[73]。
其他生物的存在也会影响盐沼系统的盐化,但是不同的物种可能存在不同的效果。蜗牛等生物以盐生植物或其根系为食[79],抑制植被的生长,进而导致盐沼湿地的退化。但也存在贻贝、牡蛎、文蛤等双壳纲生物通过分泌胞外聚合物(EPS)粘结沉积物,其自身的聚集也可以有效地减缓水流,降低侵蚀风险,进而保护盐沼湿地[80]。
3.3 人类活动
潮滩湿地具有极大的土地资源开发潜力,可为沿海地区提供巨大的社会经济效益。围垦工程作为人类获取可用土地的重要手段之一,也成为了潮滩上规模最大的人类活动之一,例如中国的围填海工程自20世纪50年代以来就已颇具规模。但围垦工程在为沿海地区城市带来多种资源利益的同时,也对潮滩环境造成了巨大的影响[81]。
围垦会全部或部分隔绝工程区内外的物质交换,改变局部的海岸地貌,影响工程区附近潮滩的波浪、潮流等水动力条件,进而影响泥沙的沉积分布与冲淤变化趋势,最后导致工程区附近潮滩的剖面形态、潮沟等地貌发生变化,且这些影响在工程区附近最为强烈[82- 84]。此外,通过对这些环境因子的影响,围垦也会促进潮滩盐沼生物的演替,造成垦区内外不同的植被演化特征[85- 86]。由于海堤的影响,垦区内与外界海域的物质交换被全部或部分隔绝,原有盐沼植被的适宜生存条件(如土壤、淹水等)发生改变,导致其消亡或演替。孙超等[87]通过遥感影像提取分析近25 a江苏中部沿海的盐沼分布发现,滩涂围垦造成的盐沼消亡面积达569.47 km2,占盐沼消亡总量的85.4%,对弶港以北盐沼影响极大。相反,垦区外的盐沼植被则会受到一定的促进作用,这是由于海堤的存在会促进堤前沉积物堆积,再加上盐沼本身的促淤作用使得潮滩进一步淤高,为盐沼先锋植被的向海扩张提供了有利的生存条件,但前提是在围垦区外留有一定面积的原生盐沼。因此,若给予盐沼一定的生态恢复期,盐沼可以恢复[88]。朱冬和高抒[89]也发现互花米草扩展对于滩涂围垦具有不同的响应,若围垦时保留足够宽度的堤前米草,则围垦可以提高互花米草的年扩展率,其机理是米草种子产量和前缘淤积环境是影响互花米草扩展的主要因素,而围垦对种子产量和前缘淤积环境这两方面都有影响。
4 盐沼潮滩地貌演变模拟
盐沼潮滩系统的演变模拟一般包括物理模型实验和数学模拟技术。
物理模型实验可以提供直观的、感性的认识,但是物理模型需要考虑比尺效应、成本投入较高,目前关于潮滩地貌演变的物理模型实验重点关注潮滩系统中潮沟的发育与演化机制。模型实验结果表明,潮沟的发育经历了由快到慢的过程,初期由于退潮期间水流的作用,潮沟发育较快,之后随着潮沟系统逐渐发育成型并趋于平衡,发育减缓[90- 94]。Stefanon等[94]的澙湖型纳潮盆地型物模实验发现,相对平均海平面的变化直接影响潮滩系统的纳潮量,而纳潮量迅速且强烈地影响着潮沟的横截面积、潮沟系统结构及其排水密度。例如,纳潮量减小会导致潮沟宽度减小和退化。Vlaswinkel和Cantelli[90]的开敞式潮滩模型实验研究同样得到潮汐流量对宽度的影响比深度更强,结果还表明平衡态下的潮沟深度与宽度以及长度与宽度之间呈线性正相关。基于洪水和退潮主导洋流之间的比较,Geng等[93]的模型结果显示,涨潮主导有利于在潮滩上部形成小规模的潮汐分支,而持续时间较长的落潮主导会形成更弯曲和更宽的潮沟。此外,Iwasaki等[92]的物模实验结果还提到了植被与模型沙材料对潮沟系统稳定性的重要性。
目前的物理模型实验仍存在一些技术难题亟待突破。例如,物理模型均缺乏加沙系统,无法模拟来自外海的泥沙输入,因此,目前所有实验潮滩系统都处于侵蚀过程中,且在没有外界输沙的情况下达到实验平衡状态后,潮沟系统结构也没有明显变化。此外,在物理模型实验中,需要按照一定的比尺将真实的潮滩系统进行缩小,由于现实中潮滩区域较大,一般要求在垂直方向进行变形(使潮滩模型的坡度变陡),如此可以更完整地模拟潮沟系统在整个潮滩区域中的演变[90- 94]。但该种变形同时会使得重力对潮汐的作用增强,影响潮汐变形和不对称性。针对这个问题,Kleinhans等[95]设计了一种新型的缓慢摆动式潮滩平台来减弱这种影响,同时该装置还可以解决其他物理模型实验在涨潮期间以及达到平衡态后泥沙移动性减弱的问题。
不同于物理模型实验,数学模拟的成本较低,可控性、可设计性更高,因此该方面的研究较多,发展较快。盐沼潮滩演变动态模拟以及模型参数需要包含前述各种影响因子之间的相互作用和制约关系,目前这方面研究正处于蓬勃发展的阶段,但准确反映、数学描述各影响因子的作用及不同影响因子的耦合效果还处于起步阶段。当前,模拟盐沼在宏观尺度上演变的数学模型大体可以分为4类,包括统计模型、元胞自动机模型、基于动力过程的数学模型以及混合模型。
统计分析是学术界基于统计学最常用的预测性建模技术之一,有些过程无法用理论分析方法推导出其模型,但可通过试验测定数据,经过数理统计法求得各变量之间的函数关系,在滨海湿地植被群落演替研究中,应用较多的统计模型包括回归分析模型以及马尔可夫模型;元胞自动机是一种能够表现系统复杂行为的模拟方法,在多个学科应用广泛,适用于研究植物群落的时空动态过程,其特点是可利用局部小尺度观测的数据分析得到相应的规律(或规则),然后利用计算机模拟研究系统大尺度的动态演化特征[96];基于动力过程的数学模型试图通过耦合植被演替中涉及的环境、生态等各种动力过程来实现演变的预测,且不同的动力过程(如潮流、波浪、植物生长等)的数学描述通常是基于较明确的物理机制,这是此类模型的巨大优势。以上3类模型各有其特点及适用性,在解决实际问题时,有学者也尝试了融合不同类型的模型,开发混合型模型,达到取长补短的效果。近年来,基于动力过程的数学模型是研究的热点,也取得了系列成果,Fagherazzi等[97]进行了一定的总结。
图3给出了盐沼潮滩演变数值模拟典型的生物动力地貌数学模型框架以及通常要考虑的动力过程[98],模型主要包括水动力、泥沙运动、地貌改变和盐沼植被这4个主要模块,不同模块之间通过相互作用和耦合共同影响系统的演变。模型计算流程:给定某初始条件,依据水动力计算的相关方程式计算水位、流速、波高等物理量,将得到的水动力结果用于驱动泥沙输送的计算,得到地貌的改变,这其中也考虑与盐沼植被的互馈关系,通常是要建立盐沼植被生物特征参数(如生物量、植株密度、高度、直径等)与水动力、泥沙等参数的数学关系;进入下一个时间步长,在新的地貌条件下重新计算水动力、泥沙运动等,依次迭代。
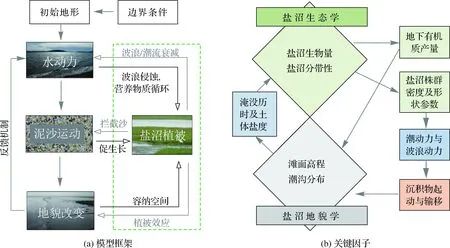
图3 盐沼生物动力地貌模型框架与关键因子[97- 98]Fig.3 Typical morphodynamic model framework of saltmarsh tidal flats and key factors
早期的盐沼生物动力地貌模型以一维剖面模型较为常见,van de Koppel等[99]通过耦合简化的水动力- 泥沙- 地貌演变方程与盐沼植被生长模型,模拟了盐沼前缘陡坎的生成与演化机制,指出盐沼前缘在自组织行为控制下的自我演化规律,即短期内自组织行为可改善盐沼系统的生境,而自组织的长期效应也可能会导致盐沼系统的消亡;在其模型的基础上,Mariotti和Fagherazzi[100]对于一些物理过程进行了更加详细的刻画,加入了具体的潮流、波浪、海平面上升、泥沙供给等动力过程;盐沼的模拟采用了现场观测生物量与高程数据进行拟合给定[101]。此外,盐沼与潮流、波浪、泥沙动力之间的相互作用也通过一些已有方程合理地进行了反映,模拟结果揭示了盐沼在海平面上升以及边界泥沙供给变化情况下的演化情况。Zhou等[98]在开源代码Delft3D中加入了风浪模块、盐沼模块,进一步分析了盐沼的弱流、消浪、截沙、固沙等生物物理效应对于潮滩多组分泥沙分选作用的影响。
由于一维模型无法反映潮沟等地貌单元,在实际应用中也报道了一些二维动力地貌模型的研究成果。一些学者开发的二维模型发现盐沼植被虽然在总体上促进淤积、减少侵蚀,但是在不同盐沼簇团之间由于水流的汇聚作用可形成较强的冲刷,有利于冲刷型潮沟的发育[102]。应用不同的盐沼植被动力地貌模型,Belliard等[103]发现泥沙供给的增大可促进潮沟的发育且最初的地貌形态对最终的地貌平衡态有较大的影响,进一步地,不同的盐沼类型之间存在促进与竞争机制,也会影响湿地地貌的演化[104]。此外,Schwarz等[20]发现植物生活史特征对于地貌塑造的巨大影响,快速定植的盐角草属植物容易形成片状分布的盐沼湿地,而慢速定植的米草属植被容易形成斑块状分布的盐沼湿地,因而,米草属植被主导的湿地更有利于潮沟的发育,这也决定了不同类型湿地的长期恢复力情况。
5 结论与展望
潮滩生物一方面通过弱流、消浪、截沙、固沙等生物物理效应影响着水动力、泥沙输运与地貌演变过程,另一方面也受到这些过程的反作用促进或抑制其自身的繁衍、生长,理解生物作用与动力地貌过程之间的双向反馈机制是认知盐沼潮滩生态系统演变的关键。早期研究较多的是盐沼、红树林等植物在海岸动力地貌演变中的贡献,近期研究也指出底栖动物和微生物也可产生一定的影响。近年来,生物动力地貌数学模型的研发方面也取得了较大进展,一定程度上揭示了盐沼潮滩的演变机制,在定性层面预测其未来演化趋势。
在全球变化的大背景下,围绕国家需求与国际前沿问题,针对盐沼潮滩的研究还有很多科学问题亟待解决、很多技术难题亟待突破,包括但不限于以下几方面:
(1) 盐沼潮滩高频率、高精度、多参数同步观测技术。野外观测数据在潮滩盐沼动力地貌研究中十分关键,无论对于科学问题发现、机理机制研究,还是模型验证,都是必需的基础资料。但是,盐沼潮滩自陆向海横跨植被区、易陷光滩区和淹没区,观测数据获取难度极大。目前主要的数据获取方式是人工跑滩、无人机观测与卫星遥感,这些方法对应了从小尺度到大尺度的资料获取,但同时观测精度也随之降低。如何更加高效地获得长期连续的观测资料,推动潮滩盐沼动力地貌模型向定量化方向发展,是亟待突破的难题。因此,应加强海- 陆- 空立体同步观测技术研究,综合运用卫星、无人机、水动力泥沙及生物观测设备,开展潮间带极浅水与潮下带淹水区水动力参数同步观测,发展滨海湿地全域高程监测技术,优化底床沉积物组分遥感反演方法,创新泥沙临界起动切应力观测方法,发展盐沼生物量遥感反演模型,实现高频、高时空分辨率数据获取,为滨海湿地演变模拟提供基础数据支撑。
(2) 盐沼潮滩生物过程与物理过程的相互作用机制。对盐沼潮滩生物作用的研究还处于起步阶段,生物作用的研究难点不仅在于其本身多种物种的不同特征表现所带来的差异(例如形态参数、生长模式、生物行为等),还具有季节性、地域性和种群竞争等特点。例如,盐生植物、动物和微生物的存在不仅影响了水流结构,还改变了泥沙的临界起动切应力。在现有的模型中该值多为固定值,未来的数值模拟需要进一步贴合实际情况,建立与时空分布、形态参数、生长模式、生物行为有关的控制方程,能够提供更为准确的地貌模型。反之,地貌的改变也影响着各类生物的分布与生长,两者相互耦合,能够更好地与现实吻合。同时结合实测数据来校准模型,提高其准确性和适用性,并利用数学模型进行机制探索和系统演变预测。
(3) 具有定量预测能力的盐沼潮滩生物动力地貌模型。针对陆地和海洋环境已建立了相对完备的数学模型,但对于盐沼潮滩这样的海陆过渡地带的复杂生态系统,还缺乏统一的数学模型,一方面是因为很多动力过程还缺乏数学描述(如复杂的生物行为、潮滩浅水边界层动力过程、岸壁坍塌过程等),另一方面是耦合这些不同时空尺度的动力过程到同一个模型难度很大。例如,目前运用到数学模型中的盐沼动力过程的表达十分粗糙,大多不考虑不同类型的盐沼植被(盐蒿、互花米草等)的地域差异性和季节差异性,这就使得模型结果的可信性存在质疑,只能停留在定性合理的层面,定量预测能力十分有限。有必要进行中长期连续测量及实验室分析,探究不同类型盐沼植被过程的时空差异,提出更加科学、详实的动力过程的数学表达,为研发具有预测能力的海陆耦合、环境- 生态动力过程耦合的动力学模型建立基础。
