桂林岩溶石山常绿落叶阔叶混交林不同叶习性植物功能性状梯度分析
2021-05-11蒋婷向珊珊方秀茹秦琦云施伟凤姜勇
蒋婷 向珊珊 方秀茹 秦琦云 施伟凤 姜勇

摘要以桂林巖溶石山常绿落叶阔叶混交林不同叶习性树种为研究对象,在55个样方88种木本植物的6种功能性状数据的基础上,采用性状梯度分析法研究不同叶习性共存物种的性状差异和生态策略选择。结果表明,常绿树种的叶绿素含量、叶厚度、叶干物质含量和木材密度高于落叶树种,而落叶树种的叶面积和比叶面积高于常绿树种。落叶树种的生态位宽度大于常绿树种的生态位宽度。说明常绿树种采取维持生长、延长叶寿命为主的保守策略,而落叶树种采取以提高资源利用效率为主的积极策略。常绿和落叶树种6个功能性状的α值变化范围都比β值的变化范围大,说明共存物种之间的相互作用比环境因素对物种性状的影响更大,且群落内的作用强于群落间的作用。常绿和落叶树种β值的相关性很高,而α值的相关性很低。说明多个植物功能性状在所处群落中表现出更高的整体差异,功能性状在群落间的相关性大于群落内的相关性。性状间的不相关或弱相关表明,不同性状在2种叶习性树种的生长发育中表现出独立的维度。研究不同叶习性物种功能性状在环境梯度上的差异,对了解常绿和落叶物种的共存机制及群落构建和生物多样性维持机制具有重要意义。
关键词常绿落叶阔叶混交林;功能性状;叶习性;性状梯度分析;桂林
中图分类号S718.5文献标识码A文章编号0517-6611(2021)08-0104-10
doi:10.3969/j.issn.0517-6611.2021.08.028
开放科学(资源服务)标识码(OSID):
GradientAnalysisofFunctionalTraitsofDifferentLeafHabitPlantsinEvergreenDeciduousBroad-leavedMixedForestinKarstHillsofGuilin,SouthwestChina
JIANGTing1,2,XIANGShan-shan1,2,FANGXiu-ru1,2etal
(1.KeyLaboratoryofWildAnimalandPlantEcologyGuangxiCollegesandUniversities,GuangxiNormalUniversity,Guilin,Guangxi541006;2.CollegeofLifeScience,GuangxiNormalUniversity,Guilin,Guangxi541006)
AbstractInthispaper,westudieddifferentleafhabittreespeciesintheevergreendeciduousbroad-leavedmixedforestofkarsthillsinGuilin,SouthwestChina.Basedonthedataof6functionaltraitsof88woodyplantsin55plots,weusedthetrait-gradientanalysistostudythecoexistenceofdifferentleafhabitspeciestraitdifferencesandecologicalstrategyselection.Theresultsshowedthat:thereweresignificantdifferencesinthefunctionaltraitsbetweenevergreentreespeciesanddeciduoustreespecies.Leafchlorophyllcontent(Chl),leafthickness(LT),leafdrymattercontent(LDMC)andwooddensity(WD)ofevergreentreespecieswerehigherthandeciduoustreespecies,whilelaminaarea(LA)andspecificleafarea(SLA)ofdeciduoustreespecieswerehigherthanevergreentreespecies.Thenichewidthofdeciduoustreespecieswaslargerthanevergreentreespecies.Itshowedthatevergreentreespeciesadoptedaconservativestrategytomaintaingrowthandextendleaflife,whiledeciduoustreespeciesadoptedapositivestrategytoimproveresourceutilizationefficiency.Therangeofthealphavalueofthesixfunctionaltraitsofevergreenanddeciduoustreespecieswaslargerthanthebetavalue,indicatingthattheinteractionbetweencoexistingspecieshadagreaterimpactonthespeciestraitsthanenvironmentalfactors,andtheroleofwithin-sitestrongerthantheroleofamong-site.Therewerehighcorrelationsbetweenthebetacomponentofevergreenanddeciduoustreespecies,whilelowcorrelationwasfoundbetweenevergreentreespeciesanddeciduoustreespeciesinthealphacomponent.Thissuggestedthatmultipleplantfunctionaltraitsshowedhigherdifferencesinthecommunitieswheretheywerelocated.Thecorrelationbetweenfunctionaltraitsamongcommunitieswasgreaterthanthatwithincommunities.No-correlationorweakcorrelationbetweentraitsindicatedthatdifferenttraitsshowedindependentdimensionsinthegrowthanddevelopmentoftwoleafhabittreespecies.Studyingthedifferencesinthefunctionaltraitsofdifferentleafhabitspeciesonenvironmentalgradientsisofgreatsignificanceforunderstandingthecoexistencemechanismofevergreenanddeciduousspecies,communityconstructionandbiodiversitymaintenancemechanisms.
为减小采样数据的偶然性,对样地中出现的每一株木本植物,选取其叶片和枝条样品各3份进行功能性状测定,然后取平均值作为该植株个体的测定结果。采用最新的全球植物功能性状标准化测量手册中的测定方法[22],利用叶面积仪(Yaxin-1241,Yaxin,China)扫描记录每片新鲜叶样品的叶面积(cm2);用精度为0.01mm的电子数显卡尺(SF2000,Guilin,China)测量叶片厚度,避开叶脉量取每片叶样品的叶尖、叶中、叶根部数据取三者平均值作为该叶片的叶厚度(mm);用便携式叶绿素含量测定仪(SPAD-502Plus,KonicaMinolta,Japan)避开叶脉测定每片叶样品的叶尖、叶中、叶根部数据取三者平均值作为该叶片的叶绿素含量(SPAD);去除新鲜叶片上的杂物后用精度为0.0001g的电子天平(BSM-220.4,Zhuojing,China)在无风环境下称量叶鲜重,然后将叶片放入已编号的信封中有序放置在80℃的烘箱中48h,叶片烘干至恒质量后取出进行称量并记录叶干重,叶面积与叶干重的比值即为比叶面积(cm2/g),叶干重与叶鲜重的比值就是叶干物质含量(g/g);为了避免取样过程对树木生长的负面影响,木材密度的测量采用直径1~2cm的树枝代替采集树干核心部分,将树枝削去树皮后立刻用电子天平称量并记录其鲜重,然后利用量筒排水法测量枝条体积,再将枝条放入已编号的信封中置于80℃的烘箱72h,枝条烘干至恒质量后取出进行称量并记录枝条干重,枝条干重与枝条体积的比值即为木材密度(g/cm3)。
1.4性状梯度分析法
根据Ackerly等[8]提出的性状梯度分析法,将植物群落(数量为P)内的物种(数量为S)按其物种多度(aij)进行加权计算得到该群落(j)的平均性状值(pj),然后根据群落平均性状值由小到大的顺序进行排序,构建环境影响下的性状梯度,再利用该性状梯度将物种平均性状值(ti)分解成β和α2个组分。其中,β组分(βi)为物种i在该性状梯度上所处的位置,体现物种i在不同群落间的变异,反映物种i对群落间环境变化的响应;而α组分(αi)是某一物种i平均性状值(ti)与其所处群落平均性状值(βi)的差值,体现了群落内某物种i与其共存物种的性状值差异,反映了对同一群落内种间交互作用等非生物环境适应策略的差异。性状梯度分析法主要计算公式:
j=Si=1aijtijSi=1aij(1)
i=Pj=1aijtijPj=1aij(2)
βi=Pj=1jaijPj=1aij(3)
αi=i-βi(4)
式中,pj为群落平均性状值,ti为物种平均性状值,aij为物种i在群落j中的多度或权重,tij为物种i在群落j中的性状值,P表示调查的群落总数,S表示调查出现的物种总数。该研究中P=55、S=88。性状梯度分析之前对不满足正态分布的数据(LA、SLA、Chl)进行lg转换。然后分别对各个功能性状进行Pearson相关分析,并用T检验对相关性大小进行分析。数据在Excel2010软件进行初步整理,进一步的数据分析计算与绘图在R软件3.6.2版本(RDevelopmentCoreTeam,2011)中完成。
将性状梯度分析法可视化有助于更直观地理解,以主要常绿树种檵木和主要落叶树种喜树的SLA为例来解释α组分与β组分的生态学意义。由图1可知,灰色空心圆点表示群落内的每个物种,黑色三角形和红色菱形分别代表檵木和喜树,横轴表示群落平均比叶面积(pj),纵轴表示物种平均比叶面积(ti),在某pj位置被矩形框起来的一列点代表共生在群落j的所有物种。虚线y=x的斜率为物种平均比叶面积(ti)与群落平均比叶面积(pj)的比值。檵木与喜树的绿色图形点对应的x坐标表示该物种在群落比叶面积梯度上的平均位置,即i物种比叶面积的βi,因为αi=ti-βi,绿色图形点偏离虚线y=x的距离表示i物种比叶面积的αi。虚线bi表示喜树的物种平均比叶面积与群落平均比叶面积的回归直线的斜率,体现种内物种平均比叶面积随群落平均比叶面
积的变化而变化。依据上述公式计算得出常绿树种檵木的β组
分相對较小(lgSLAβi=2.26),位于由SLA的群落加权平均值构成的性状梯度的下部,表明檵木常生长在低SLA的群落中,且檵木的SLA小于群落均值(lgSLAαi=-0.02),反映了檵木的SLA相对低于共存物种。而喜树的β组分相对较大(lgSLAβi=2.38),处于性状梯度的上半部,表明喜树多生长于高SLA的群落中,且喜树的SLA大于群落均值(lgSLAαi=0.04),反映了喜树的SLA相对高于共存物种。檵木和喜树比叶面积具有相似的生态位宽度。
2结果与分析
2.1常绿和落叶树种6种功能性状之间不同参数的比较
由表1可知,常绿树种和落叶树种的6种功能性状值均存在一定的差异,其中落叶树种的叶面积和比叶面积的物种平均性状值和样地平均性状值都高于常绿树种,而常绿树种的Chl、LT、LDMC和WD均值都高于落叶树种。常绿树种各功能性状的生态位宽度小于落叶树种的生态位宽度。常绿和落叶树种6种功能性状的α值变化范围均大于β值的变化范围,且6种功能性状值的α和β值变化范围均存在差异。落叶树种LA和WD的α值变化范围大于常绿树种,但常绿树种SLA、Chl、LT和LDMC的α值变化范围大于落叶树种。常绿树种除SLA外,LA、Chl、LT、LDMC和WD的β值变化范围均大于落叶树种。落叶树种LA和WD的α值小于0的多于常绿树种,但LT的α值小于0的少于常绿树种。与β值相比,不同叶习性树种功能性状α值的分布趋势更加相近(图2、表2、图3、表3)。
2.2常绿和落叶树种6种功能性状不同参数的相关性
由表4可知,常绿和落叶树种6种功能性状的生态位宽度与物种平均性状值、生态位宽度与α值均无显著相关性(P>0.05)。常绿树种6种功能性状的β值、落叶树种的LDMC和WD的β值均与物种平均性状值无显著相关性(P>0.05)。除常绿树种的LT与物种平均性状值存在极显著(P<0.01)的正相关外,常绿和落叶树种LA、SLA、Chl、LDMC和WD的α值与物种平均性状值均呈极显著(P<0.001)正相关性(r>0.64)。常绿树种LT的α和β值存在极显著负相关性(r=-0.835),常绿树种Chl的α和β值存在显著(P<0.05)负相关性(r=-0.306),落叶树种LA的α和β值存在显著正相关性(r=0.294)。落叶树种除LT的β值与物种平均性状值存在极显著(P<0.001)正相关性外,其LA、SLA、Chl的β值与物种平均性状值存在极显著(P<0.01)正相关性。落叶树种的性状β值与物种平均性状值的相关系数均高于常绿树种。
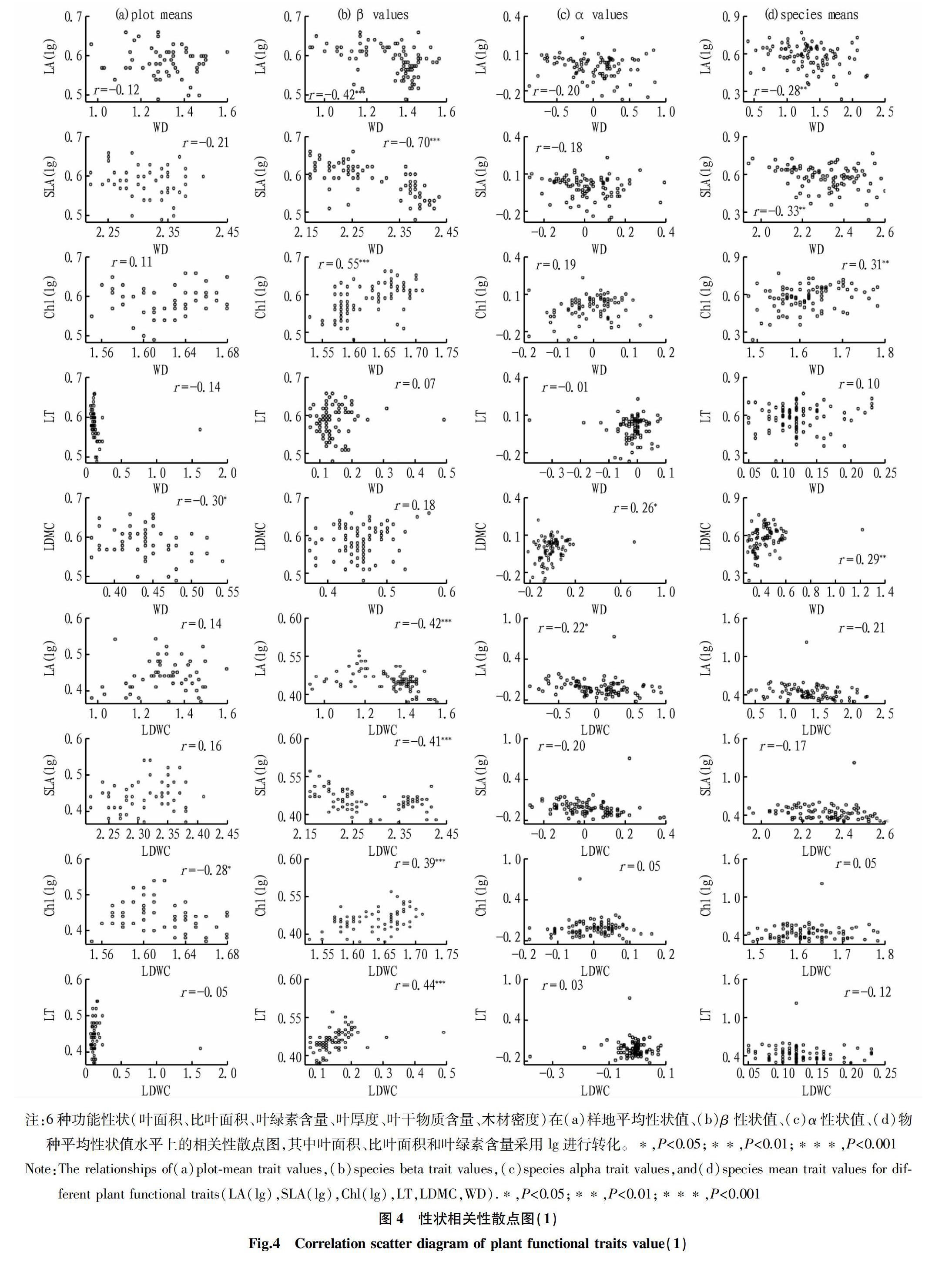
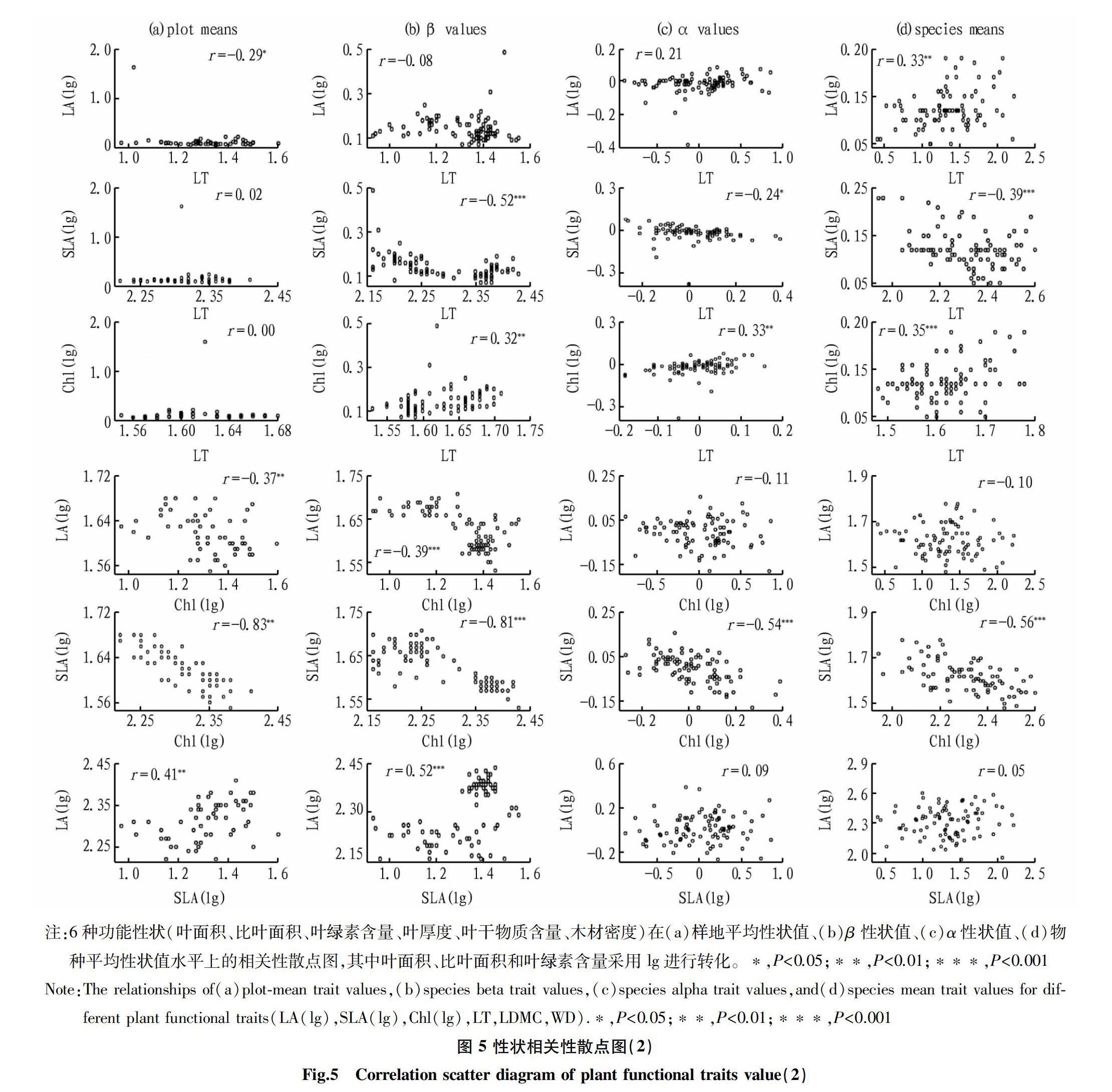
由图4、图5可知,不同植物功能性状的样地平均值、β值、α值和物种平均值多半表现出显著的相关性,也有小半呈弱相关或不相关,其中各个功能性状间β值的相关性更为明显。在各功能性状的样地平均性状值中,WD与LDMC显著负相关,LDMC与Chl显著负相关,LT与SLA显著负相关,Chl与LA、SLA均显著负相关,但SLA与LA显著正相关。除WD与LT、LDMC之间以及LT和LA之间β值不存在显著相关性外,其他各功能性状间的β值均呈显著相关性。对比β值和物种平均性状值,样地平均性状值和α值相关性较弱,其中WD与LDMC的α值和物种平均性状值呈显著正相关,而LT与SLA、Chl与SLA的α值和物种平均性状值呈显著负相关。
3讨论
3.1常绿和落叶树种的生存策略差异
不同叶习性(落叶和常绿)之间功能性状的变化可能直接驱动群落的功能多样性[23-25]。分析测得的9867个个体的性状数据可以发现,常绿和落叶树种的物种平均性状值存在显著差异。常绿树种的Chl、LT、LDMC和WD高于落叶树种,而落叶树种的LA和SLA高于常绿树种。常绿树种和落叶树种的不同生态策略为二者在桂林岩溶石山的常绿落叶阔叶混交林中稳定共存提供了机会。在胁迫性生境下,常绿树种会选择相对保守的生存策略,选择组织强健且资源周转慢的功能性状,较低的SLA和较高的LDMC支持较低的光合作用和蒸腾速率,较高的Chl、LT和WD,有利于全年的净光合作用和植物生长,抵御低温和干旱环境[26]。这种缓慢的投资回报策略有助于常绿树种在土壤贫瘠的生境中保持养分,这使它们具有竞争优势。与此相反,落叶树种年生长周期短,必须获得大量的养分才能在与常绿植物共存的环境中形成竞争优势,SLA高的物种往往营养浓度较高其光合作用和暗呼吸速率较快[27],落叶树种的高LA和SLA可以促进植物生长并获得更多的碳水化合物[28]。这反映出常绿树种对环境的主要适应策略以维持生长、延长叶寿命为主,而落叶树种的主要适应策略以提高资源利用效率为主[20-21]。该研究结果与刘微等[29]在研究不同生境中常绿和落叶树种的叶片氮素分配与光合能力的关系时得出的结果基本一致。
常绿和落叶树种的叶片、木材和生活型特征都受到群落中环境条件的影响,如光照、温度、土壤湿度和养分等[30]。物种性状的分异反映了它们对不同环境的适应。生态位宽度可以解释种群的环境适应性和资源利用能力[31]。生态位重叠较大的物种具有相似的生态特征或对生境因子的互补要求,而物种共存的基本机制之一就是物种的生态位分化,即物种之间所利用的生境资源存在差异[32]。桂林岩溶石山常绿落叶阔叶混交林常绿树种和落叶树种共存,说明该生境适宜两者共同生长。落叶树种的生态位宽度大于常绿树种的生态位宽度,表明常绿树种所利用的生境资源较为集中,而落叶树种能够利用的资源更为广泛其适应性较大。由于具有不同特征的物种往往有不同的资源需求,竞争往往比具有相似特征的物种弱,常绿和落叶树种生态策略的差异降低了群落内的竞争强度,因此具有不同功能特征的物种可以通过划分资源更好地共存[23]。
3.2常绿和落叶树种功能性状α和β值的大小反映了群落内分化和群落间重叠
环境筛选和生物竞争的不同作用促使植物功能性状发生变化,影响共存物种所采取的生态策略[24]。环境筛选使共存物种生态重叠出现相似性状,生物竞争使共存物种生态分化出现相异性状[6]。植物功能性状是生态策略变异的维度。α和β性状的维数显示了群落内和群落间变异的差异,并为植物群落组合提供了解释[12]。植物功能性状的β值代表物种某一群落平均性状值所构成的性状梯度上的位置,描述了群落间的功能性状差异,反映的是群落构建中的环境筛选作用,α值则代表某一物种偏离其所处群落平均性状值的大小,描述了群落中的功能性状差异,反映的是群落构建中的生物竞争作用。该研究中常绿和落叶树种6个功能性状的α值变化范围均比β值的变化范围大,这意味着在桂林岩溶石山常绿落叶阔叶混交林的构建过程中共存物种之间的相互作用比环境因素对物种性状的影响更大,且群落内的作用強于群落间的作用。原因在于,其一,环境筛选在环境梯度差异更显著的条件下(跨越较大的纬度梯度,生境的气候差异巨大;或跨越较大的经度梯度,海陆位置的水分影响;亦或跨越较大的垂直梯度,受到气温影响等)对物种的性状变异影响大[12]。该研究的调查区域相对较小,各群落可能有相似的环境条件,因此性状对环境的反应可能已经收敛,即β值的变化范围较小。其二,研究对象具有特殊性,桂林岩溶石山的亚热带常绿落叶阔叶混交林中常绿树种和落叶树种共存,由于性状差异它们采取不同的生态策略,所以种间和种内竞争可能非常激烈。但常绿树种的α值变化范围比落叶树种的α值变化范围更大,该研究结果与处于群落演替晚期的湖北星斗山亚热带常绿落叶阔叶混交林中落叶植物比常绿植物能够更好地利用资源存在差异[33]。这表明桂林岩溶石山的常绿树种比落叶树种拥有更好的适应恶劣生境下物种共存的策略。为了适应当地的生境条件,常绿和落叶物种都表现出性状特征趋同(重叠的β值),每个功能性状的α值具有不同程度的分化,这表明种内竞争导致性状分化[34],这是决定群落稳定性的一个重要因素[35]。如SLA的变化范围反映了常绿和落叶树种对光的不同反应策略,常绿树种通过降低其比叶面积(负α值)来提高其对光资源的利用效率。然而,大多数落叶植物在生长季节获得的光资源具有较高的比叶面积(正α值)。
3.3常绿和落叶树种的α和β性状相关性
植物生长过程中对生境的适应不是通过改变单一性状来实现的而是多种功能性状协调作用,所以植物功能性状之间总是存在某种关联[36-37]。常绿和落叶树种β组分的相关性很高,而α组分的相关性很低,表明多个植物功能性状在所处群落中表现出更高的整体差异,但在更大范围内对环境过滤的反应是收敛的,功能性状在群落间的相关性大于群落内的相关性。α组分的弱相关性表明每个性状都是独立分化轴,共存物种间的相互作用越大,物种α性状与群落平均值的偏差越大。根据有限相似理论,当群落功能性状发生变化时,物种可以获得更多的资源[38]。而β组分之间的强相关性可能与在相同生境中过滤条件下性状的收敛有关。群落内的环境异质性可能促进性状协调,因为它导致具有不同性状值的物种共存。相反,缺乏协调表明,性状代表不同的生态策略,是促进植物器官生态位分化的独立轴[8,39]。物种平均性状值与各性状α的相关性均达极显著水平,但β值与α值之间无显著相关性。局部生物过程如竞争增加了性状的差异性,并允许植物通过生态位分化的方式共存,功能特性之间的权衡揭示了主要的生态策略,这些策略可以提供对物种分布和生态系统过程的洞察[9-11]。
4结论
该研究以桂林岩溶石山常绿落叶阔叶混交林不同叶习性(常绿和落叶)树种为研究对象,探讨桂林岩溶石山常绿落叶阔叶混交林中,不同叶习性物种对环境变化的响应和群落内共存物种的适应策略是否存在差异。采用性状梯度分析法将物种的性状值分为α(群落内)和β(群落间)2个分量,定量分析不同叶习性物种的功能性状在群落内和群落间的性状变异规律,从而揭示研究样地内常绿与落叶共存物种的性状差异及其在环境梯度上的性状分化和生态策略选择。结果表明,
①常绿树种对环境的主要适应策略以维持生长、延长叶寿命为主,而落叶树种的主要适应策略以提高资源利用效率为主。但落叶树种的生态位宽度大于常绿树种的生态位宽度。常绿与落叶树种通过一定特性(α组分)的生态定位共存,这种定位减少了竞争,并允许它们获得更多的资源。
②环境过滤和生物竞争共同作用于桂林岩溶石山常绿落叶阔叶混交林的群落构建过程。环境过滤使局域群落内共存的物种产生相似功能性状,性状β组分存在收敛,而不同叶习性共存物种又通过种内变异产生不同的生态策略选择,性状α组分产生分化,从而减小群落内的生物竞争。
③功能性状α值的变化范围明显大于β值的变化范围,说明桂林岩溶石山常绿落叶阔叶混交林各样地内物种间的协调作用大于样地间不同环境因子对塑造物种性状的影响。常绿树种α值的变化比落叶树种α值的变化更显著,表明桂林岩溶石山的常绿树种较落叶树种拥有更好地适应恶劣生境下物种共存的策略。
④常綠和落叶树种β值的相关性很高,而α值的相关性很低。表明多个植物功能性状在所处群落中表现出更高的整体差异,但在更大范围内对环境过滤的反应是收敛的,功能性状在群落间的相关性大于群落内的相关性。性状间的不相关或弱相关表明,不同性状在2种叶习性树种的生长发育中表现出独立的维度。常绿和落叶物种平均性状值与α性状值的相关性非常显著。落叶树种的β和α性状值之间不存在显著相关性。
参考文献
[1]贺鹏程,叶清.基于植物功能性状的生态学研究进展:从个体水平到全球尺度[J].热带亚热带植物学报,2019,27(5):523-533.
[2]靳莎,闫淑君,黄柳菁,等.植物叶功能性状间的权衡研究进展[J].四川林业科技,2019,40(5):96-103.
[3]WEIHERE,FREUNDD,BUNTONT,etal.Advances,challengesandadevelopingsynthesisofecologicalcommunityassemblytheory[J].PhilosophicaltransactionsoftheroyalsocietyB:Biologicalsciences,2011,366(1576):2403-2413.
[4]CORNWELLWK,ACKERLYDD.CommunityassemblyandshiftsinplanttraitdistributionsacrossanenvironmentalgradientincoastalCalifornia[J].Ecologicalmonographs,2009,79(1):109-126.
[5]MOVI,VIOLLEC,KRAFTNJB,etal.ShiftsintraitmeansandvariancesinNorthAmericantreeassemblages:Speciesrichnesspatternsarelooselyrelatedtothefunctionalspace[J].Ecography,2015,38(7):649-658.
[6]MIDOLOG,DEFRENNEP,HLZELN,etal.Globalpatternsofintraspecificleaftraitresponsestoelevation[J].Globalchangebiology,2019,25(7):2485-2498.
[7]WRIGHTIJ,REICHPB,WESTOBYM,etal.Theworldwideleafeconomicsspectrum[J].Nature,2004,428(6985):821-827.
[8]ACKERLYDD,CORNWELLWK.Atrait-basedapproachtocommunityassembly:Partitioningofspeciestraitvaluesintowithin-andamong-communitycomponents[J].Ecologyletters,2007,10(2):135-145.
[9]梁士楚,刘润红,荣春艳,等.漓江河岸带植物功能性状变异与关联[J].植物生态学报,2019,43(1):16-26.
[10]李月娟,李娇凤,常斌,等.桂林岩溶石山青冈群落植物功能性状的尺度变化与关联[J].生态学报,2019,39(15):5555-5563.
[11]刘润红,梁士楚,黄冬柳,等.漓江河岸带木本植物功能性状跨尺度变异研究[J].生态学报,2019,39(21):8038-8047.
[12]KOOYMANR,CORNWELLW,WESTOBYM.PlantfunctionaltraitsinAustraliansubtropicalrainforest:Partitioningwithin-communityfromcross-landscapevariation[J].Journalofecology,2010,98(3):517-525.
[13]许驭丹,董世魁,李帅,等.植物群落构建的生态过滤机制研究进展[J].生态学报,2019,39(7):2267-2281.
[14]方晓峰.常绿阔叶林中常绿与落叶树种的多样性格局及其共存[D].上海:华东师范大学,2016.
[15]盘远方,李娇凤,黄昶吟,等.桂林岩溶石山不同坡向灌丛植物多样性与土壤环境因子的关系[J].广西植物,2019,39(8):1115-1125.
[16]盘远方,陈兴彬,姜勇,等.桂林岩溶石山灌丛植物叶功能性状和土壤因子对坡向的响应[J].生态学报,2018,38(5):1581-1589.
[17]胡刚.桂林岩溶石山青冈栎群落生态学研究[D].桂林:广西师范大学,2007.
[18]姜勇.海南岛热带森林植物功能性状及其多样性研究[D].北京:中国林业科学研究院,2015.
[19]ROSSATTODR,HOFFMANNWA,FRANCOAC.Differencesingrowthpatternsbetweenco-occurringforestandsavannatreesaffecttheforest-savannaboundary[J].Functionalecology,2009,23(4):689-698.
[20]路兴慧,丁易,臧润国,等.海南岛热带低地雨林老龄林木本植物幼苗的功能性状分析[J].植物生态学报,2011,35(12):1300-1309.
[21]OSUNKOYAOO,SHENGTK,MAHMUDNA,etal.Variationinwooddensity,woodwatercontent,stemgrowthandmortalityamongtwenty-seventreespeciesinatropicalrainforestonBorneoIsland[J].Australecology,2007,32(2):191-201.
[22]PREZ-HARGUINDEGUYN,DAZS,GARNIERE,etal.Newhandbookforstandardisedmeasurementofplantfunctionaltraitsworldwide[J].Australianjournalofbotany,2013,61(3):167-234.
[23]TOMLINSONKW,POORTERL,BONGERSF,etal.Relativegrowthratevariationofevergreenanddeciduoussavannatreespeciesisdrivenbydifferenttraits[J].Annalsofbotany,2014,114(2):315-324.
[24]马芳.东灵山暖温带落叶阔叶林木本植物动态与物种共存机制研究[D].北京:中央民族大学,2019.
[25]潘元琪.古田山常绿阔叶林木本植物物候对气候的响应研究[D].成都:成都理工大学,2019.
[26]谢玉彬,马遵平,杨庆松,等.基于地形因子的天童地区常绿树种和落叶树种共存机制研究[J].生物多样性,2012,20(2):159-167.
[27]MARKESTEIJNL,POORTERL,BONGERSF.Light-dependentleaftraitvariationin43tropicaldryforesttreespecies[J].Americanjournalofbotany,2007,94(4):515-525.
[28]POORTERL,KITAJIMAK.Carbohydratestorageandlightrequirementsoftropicalmoistanddryforesttreespecies[J].Ecology,2007,88(4):1000-1011.
[29]刘微,李德志,纪倩倩,等.两种生境常绿和落叶树种叶片氮素分配及与光合能力的关系[J].生态科学,2015,34(1):1-8.
[30]MOLESAT,WARTONDI,WARMANL,etal.Globalpatternsinplantheight[J].Journalofecology,2009,97(5):923-932.
[31]孙梅,田昆,张贇,等.植物叶片功能性状及其环境适应研究[J].植物科学学报,2017,35(6):940-949.
[32]CHESSONP.Mechanismsofmaintenanceofspeciesdiversity[J].Annualreviewofecologyandsystematics,2000,31(1):343-366.
[33]储诚进,王酉石,刘宇,等.物种共存理论研究进展[J].生物多样性,2017,25(4):345-354.
[34]YAOLJ,DINGY,YAOL,etal.Traitgradientanalysisforevergreenanddeciduousspeciesinasubtropicalforest[J].Forests,2020,11(4):1-17.
[35]黄永涛,姚兰,艾训儒,等.鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析[J].植物生态学报,2015,39(10):990-1002.
[36]于文英,高燕,逄玉娟,等.山东银莲花叶片形态结构对异质生境和海拔变化的响应[J].生态学报,2019,39(12):4413-4420.
[37]鐘巧连,刘立斌,许鑫,等.黔中喀斯特木本植物功能性状变异及其适应策略[J].植物生态学报,2018,42(5):562-572.
[38]唐青青.亚热带常绿落叶阔叶混交林的植物功能性状变异[D].北京:中国林业科学研究院,2016.
[39]FORNARADA,TILMAND.Plantfunctionalcompositioninfluencesratesofsoilcarbonandnitrogenaccumulation[J].Journalofecology,2008,96(2):314-322.
