基于Illumina Miseq分析黄精根腐病根际土壤真菌群落结构及多样性*
2021-04-12卢圣鄂任风鸣黄红燕
卢圣鄂,肖 波,任风鸣,卓 维,黄红燕
(重庆市药物种植研究所 重庆 408435)
黄精(Polygonatum sibiricum)是百合科黄精族黄精属(PolygonatumMill)多年生草本药食两用植物,《中国药典》2020年版收录的滇黄精(Polygonatum kingianumCol1. et Hemsl.)、黄 精(Polygonatum sibircumRed.)和 多 花 黄 精(Polygonatum cyrtonemaHua.),为其原生药[1]。黄精含有丰富的对人体有益的化学成分[2,3],传统医学认为,黄精具有补肾益精、滋阴润燥之功效[4,5]。现代医学研究发现,黄精还具有抗衰老、提高免疫力、调节造血能力、消炎、抗肿瘤和降血糖等多种功效[6,7]。黄精属植物正处于野生变家种阶段,但由于管理不当,种植年限延长,土壤微生物结构变化等因素导致根腐病等多种病害逐年加重,严重影响到黄精根状茎的产量和品质。
真菌和细菌会导致中药材根腐病的发生[8],植物的健康和营养情况与根际土壤的肥力和群落结构组成有重要的关系[9,10]。但是,药用植物根腐病还不能有效预防,通过对根际土壤真菌群落结构组成和多样性分析,对黄精根腐病的发生能有进一步的认识。吴依婷[11]等研究了采集自湖南洪江根腐病黄精土壤,分离菌出了尖孢镰刀菌(F.oxysporum),而该菌常引发作物根腐病害。同时分离出了对黄精根腐病菌株具有较高的抗菌活性的内生菌,经鉴定为枯草芽孢杆菌(Bacillus subtilis)。韩凤[12]等研究确定尖孢镰刀菌和腐皮镰刀菌是引起多花黄精根腐病的病原菌。但是针对一种病原菌简单的分离和回接,并不足以深入了解黄精根腐病致病菌,高通量测序技术能够有效迅速准确测定真菌群落结构。该项技术已经被用来揭示三七连作障碍[9],黄连[13]、西洋参[14]、苹果根腐病[15]与根际土壤微生物组成及多样性研究。因此本项目采用Novaseq-PE250 测序平台对4年生黄精根腐病根际土壤(GG组)、4年生健康黄精根际土壤(GJ组)和未种过黄精的土壤(GK 组)真菌ITS1 序列进行高通量测序,目的在于了解真菌群落与根腐病发生的关系,找到与根腐病发生相关的真菌类群,更有助于对黄精根腐病致病菌、致病机制进行深入研究。
1 材料与方法
1.1 根际土壤样品采集
土壤样品采集自四川省广安市邻水县坛同镇蜀耕农业开发有限公司黄精种植基地。海拔344 米,106°46'E,30°7'N,土壤类型为酸性紫色土,年降水量为1500 mm,年平均气温为16℃。样品分为3 组:①4年生的根腐病黄精根际土壤采集3组样品;②4年生的健康黄精根际土壤采集3 组样品;③距离采集点约50 m 未种过黄精的土壤作为空白采集3 组样品。黄精植株采集采用五点取样法,取黄精根茎周围3-5 mm 左右的土壤,待充分混匀后用无菌PET 树脂袋封装放于冰盒中带回实验室。
1.2 土壤微生物总DNA提取和PCR扩增
取土样提取土壤总DNA,称取0.5 g 土壤样品,使用Fast DNA TMSPIN Kit for Soil(MP, USA)提取土壤总DNA,依据检测DNA 样品的浓度后,将DNA 稀释到10 ng·µL-1,存于-80℃冰箱待用。使用ITS5F(5'-GGAAGTAAAAGTCGTAACAAGG-3')和ITS1R(5'-GCTGCGTTCTTCATCGATGC-3')为引物对ITS1 区进行扩增,扩增体系为20 µL(4 µL 5×Fastpfu Buffer,2µL 2.5 mM dNTP,0.8 µL 10 ng·µL-1引物ITS5F,0.8µL 10 ng·µL-1引物ITS1R,0.4 µL Fastpfu Polymerase,0.2 µL BSA,10 ng DNA)。扩增后取3 µL 利用2%琼脂糖凝胶电泳检测PCR产物。
1.3 文库制备与上机测序
采用TruSeq Nano DNA LT Library Prep Kit 制备测序文库,使用Novaseq-P E250 测序平台测序,由上海派森诺生物科技股份有限公司完成。

图1 患根腐病、健康根际土壤和空白土壤OUT数量关系
1.4 数据分析
数据采用SPSS 22.0(New York,美国)进行实验数据统计分析,差异显著性分析采用单因素方差分析(ANOVA)。α多样性分析,β多样性分析采用QIIME(Version 1.7.0)软件包进行。物种的α 多样性通过用Observed_species、Shannon 指数、Simpson 指数和Chao1指数4 个指标进行表征。在分类学层面,通过多种无监督和有监督的测序、聚类和建模方法,结合相应的统计检验方法,进一步衡量不同样本间物种丰富度组成的差异。用R 软件(version 2.15.3)分析和生成稀释度曲线、Venn 图、相对丰度图和50 个优势真菌属的热图。
2 结果与分析
2.1 测序数据和OTU聚类分析
本研究采用Novaseq-PE250 测序平台对黄精病株、健株和空白土样真菌进行高通量测序分析,通过对土壤中标准真菌ITS1 进行测序,去除低质量、Barcode 和引物序列后,9 个样品得到有效序列总数为1063335 条,其中健株土样有332391 条、病株土样有399724 条、空白组土样有331220 条。从韦恩图(图1)可以看出,黄精健株、病株和空白根际土壤真菌之间有共同的107 个OTUs,健康植株土样有1109 个特有的OTUs,空白土样有901 个特有的OTUs,而根腐病病株土壤仅有363个特有的OTUs。Venn图的变化说明,根腐病导致了黄精根际土壤真菌OTUs 数的减少。从稀释曲线(图2)可以看出,数据曲线延展平缓合理,本次测序数据量合理。
2.2 土壤真菌α-多样性分析
对黄精患根腐病株、健株根际和空白土壤真菌进行α-多样性分析表明(表1),种群多样性指数(Shannon)根腐病株最低,健株其次,空白组最高,根腐病株与健株和空白组差异显著;Simpson 指数根腐病株和健株相同,空白组最高;种群丰富度指数(Chao1和Observed_species)根腐病株最低,健株其次,空白组最高,且根腐病株与健株和空白组差异显著(P<0.05)。

表1 患根腐病、健康根际土壤和空白土壤真菌α-多样性
2.3 不同分类水平上相对丰度分析
患根腐病、健康根际和空白土壤门分类水平中的主要优势菌群均为子囊菌门、担子菌门、Zoopagomycota、毛霉门、壶菌门(图3)。患病土样中子囊菌门的相对丰度显著高于健康土样。健株、病株和空白根际土壤真菌组成差异较大,子囊菌门在根腐病根际土壤中为优势菌群,相对丰度要显著高于健康土样和空白土样,而毛霉门的相对丰度要显著低于健康土样和空白土样(P<0.05)。

图2 稀释曲线

图3 根腐病、健康根际土壤和空白土壤真菌门组成柱形图
在属分类水平上(图4),3组真菌群落结构组成及相对丰度差别较大,已报道的与黄精根腐病相关的病原菌镰刀菌属(Fusarium)存在于所有样品中,而有益菌木霉菌属(Trichoderma)在健康土样和空白土样的相对丰度显著高于患病土样。蓝状菌属(Talaromyces)在根腐病土壤中为优势菌属,相对丰度要显著高于健康土样和空白土样(P<0.05)。

图4 根腐病、健康根际土壤和空白土壤真菌门组成柱形图
2.4 根际土壤真菌聚类分析
基于样本距离矩阵做UPGMA 聚类分析,并将聚类结果与各样品在门水平上的物种相对丰度整合展示在中(图5),GG1、GG2 和GG3 这3 个病株样本聚在一起,GK1 和GK2 这2 个空白样本聚在一起,GJ1、GJ2、GJ3 和GK3 共3 个健康样本和1 个空白样本聚到一起,这种结果说明,黄精根腐病株与健株和空白样本的根际土壤真菌存在明显差异。
使用R 软件,将属水平的群落组成数据根据分类单元的丰度分布加以聚类,根据聚类结果对分类单元和样品分别排序,并通过热图呈现(图6)。丰度前50位的属聚类热图,能够更好的反映不同样本中真菌群落结构的差异。健康、空白、病株土壤微生物群落有较大差异,健株和空白微生物群落分布相对较为一致,群落结构更为多样,群落丰度更高。病株与健株和空白比较,且微生物群落结构较为单一,群落丰度亦较低。

图5 基于样本距离矩阵的UPGMA聚类树
从图6可以看出,患根腐病株土样中Exophiala(外瓶酶属)、Knufia、Eurvularia、Meyerozyma、Apiotrichum、Papiliotrema、Vanrija、Aspergillus( 曲 霉 属) 、Oidiodendorn(树粉孢属)、Scytalidium(柱顶孢霉属)、Talaromyces(篮状属)较健株土样和空白土样具有较高的物种相对丰度。
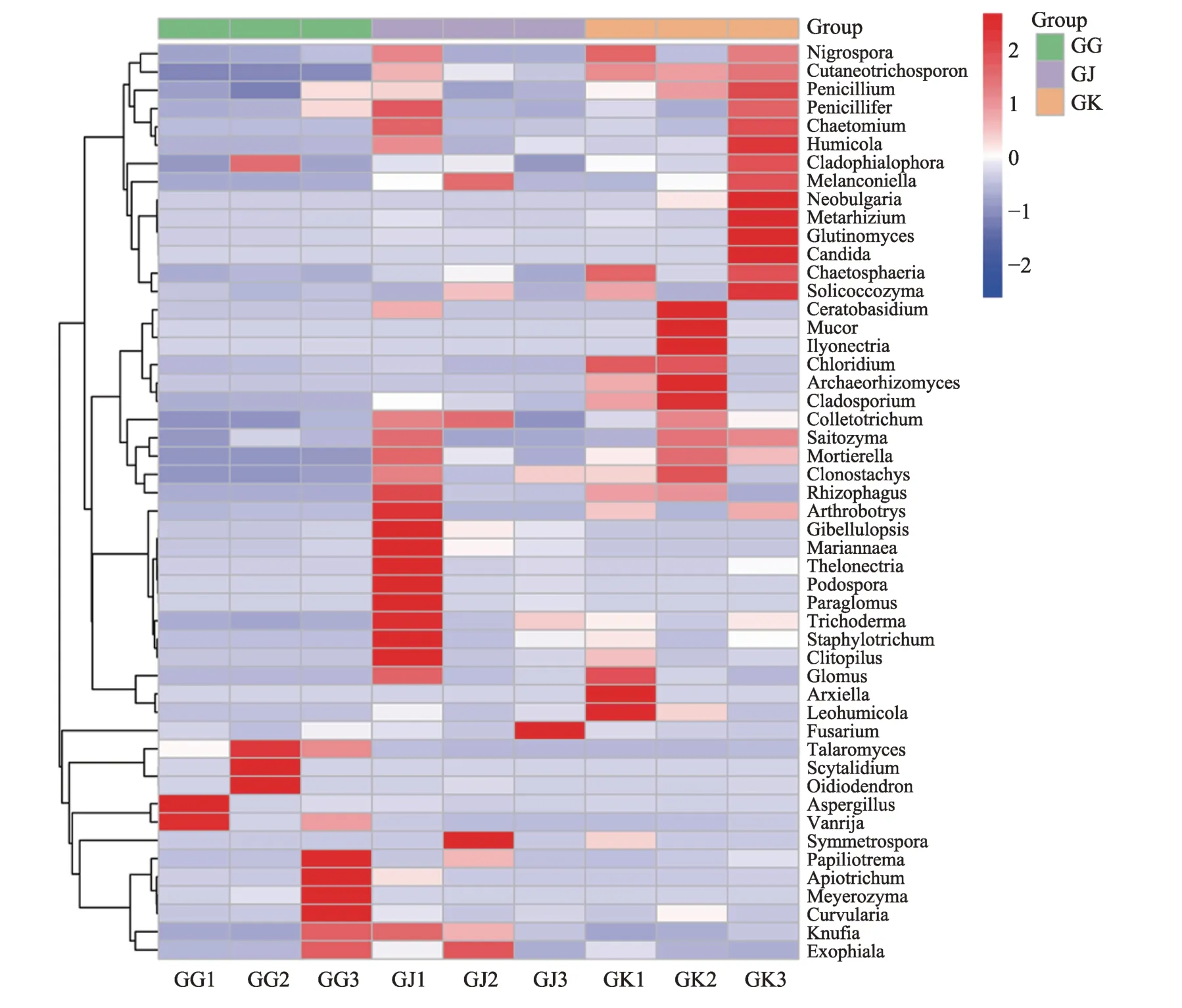
图6 土壤真菌丰度前50的菌属相对丰度Heatmap图
而健康根际土样和空白土样中Nigrospora(黑孢属)、Cutaneotrichosporon、Penicillium(青 霉 属)、Penicillifer、Chaetomium(毛壳菌属)、Humicola(腐质霉属)、Cladophialophora、Melanconiella(黑盘孢属)、Neobulgaria(螺菌属)、Metarhizium(绿僵菌属)、Glutinomyces、Candida(假丝酵母属)、Chaetosphaeria、Solicoccozyma、Ceratobasidium(角担菌属)、Mucor(毛霉属)、Trichoderma(木霉菌属)等相对丰度则高于病株土样。
3 讨论
土壤微生物群落结构能影响到土壤健康状况,其变化影响到土壤健康的变化[16,17]。植物根际土壤被病原菌感染后会导致微生物结构变化。真菌会分解植物的残体和影响植物疾病的发生[9]。Xu[18]等发现根的健康状况与真菌群落结构之间存在着明确的关系。
通过对中药材根腐病的研究证实,同种药材在各个产地和各个生长时间里的病原菌都不相同[8]。只对单一病原菌的分离培养和回接,不能完整认识导致黄精根腐病的病原菌,因此本项目采用高通量测序技术,目的在于了解认识黄精根腐病与根际土壤真菌群落结构的关系,更有助于对黄精根腐病致病菌、致病机制进行深入研究。
前人研究得出,植物土壤传播病害的发生与土壤微生物多样性的降低有很大的关系[19]。土壤微生物群落结构丰富会压制土壤病害的发生[20,21]。Wu[22]等研究发现,患根腐病的三七较健康三七的根际土壤微生物多样性较低。本研究中黄精根腐病株、健株和空白组黄精根际土壤真菌进行α-多样性分析表明,种群多样性指数(Shannon)根腐病株最低,健株其次,空白组最高,根腐病株与健株和空白组差异显著,Simpson 指数根腐病株和健株相同,空白组最高,种群丰富度指数(Chao1和Observed_species)根腐病株最低,健株其次,空白组最高,且根腐病株与健株和空白组差异显著。这与Wu[22]等的研究结果不一致,研究证实,三七健康植株根际土壤真菌丰度低于病株。余妙[14]研究显示西洋参根腐病组真菌群落多样性指数降低,真菌种类减少,达到最小。宋旭红[13]研究证实,黄连根腐病株根际土壤群落结构多样性指数高于健康植株,种群丰富度指数则稍高于病株土样。杨光柱[15]研究结果表明苹果患根腐病土样真菌多样性指数和丰富度都高于健康组,均匀度指数低于健康组。
本研究中患病及健康根际土壤门分类水平中的主要优势菌群均为子囊菌门、担子菌门、Zoopagomycota、毛霉门、壶菌门。患病土样中子囊菌门的相对丰度显著高于健康土样。与前人对黄连[23]、三七[22]、柑橘[24]等的研究结果相一致,余妙等人[14]研究西洋参根腐病也发现了相同的结果。杨光柱[15]研究结果表明苹果患根腐病土样健康和患病土样均观察到子囊菌门、担子菌门、壶菌门。对两组样本的优势菌群进行差异性分析,并且健康土样中子囊菌门的相对丰度显著高于患病土样。
中药材根腐病被认为是由镰刀菌属中的尖孢镰刀菌(F.oxysporum)和腐皮镰刀菌(F.solani)导致的[8],并且发现石柱黄连根腐病的一种致病菌是腐皮镰刀菌[25]。尖孢镰刀菌造成欧盟等国的观赏性植物枯萎、猝倒等,同时造成中药材根腐病的发生[26-30]。同时还发现苹果根腐病土壤中其相对丰度高于健康土壤。韩凤[12]等研究确定尖孢镰刀菌和腐皮镰刀菌是引起多花黄精根腐病的病原菌。吴依婷[11]等研究从黄精根腐病株中分离得到的菌株为尖孢镰刀菌(Fusarium oxysporum)。本研究在属分类水平上,3组真菌群落结构组成有较大差别,已报道的与黄精根腐病相关的病原菌镰刀菌属存在于所有样品中,而有益菌木霉菌属在健康土样和空白土样的相对丰度显著高于患病土样。篮状菌属在根腐病土壤中为优势菌属,相对丰度要显著高于健康土样和空白土样。
通过对根际土壤真菌群落结构的研究,也能找到拮抗微生物存在的线索[31]。在生物防治中木霉菌属因应用潜力广受瞩目[27],通过研究得出,木霉属真菌在健康组和空白组中高于病株。其中M.anisopliae是一种被当做微杀虫剂使用的昆虫病原真菌[32,33]。黄精及其他药用植物生态化种植的顺利开展需要利用好土壤中的优质拮抗真菌。
总之,黄精根际土壤真菌群落结构丰富度的降低和具有致病性的真菌相对丰度的增加有可能导致了黄精根际土壤微生物群落结构的改变和根腐病的发生。通过高通量测序分析,能够较好的去了解黄精根腐病组与健康组根际土壤真菌群落结构的差别,给黄精根腐病的解决提供新的思路。
