三江源区不同建植年限人工草地根系动态特征
2021-03-22李洁潘攀王长庭胡雷陈科宇杨文高
李洁 ,潘攀 ,王长庭 *,胡雷 ,陈科宇 ,杨文高
(1. 西南民族大学生命科学与技术学院,四川成都610041;2. 西南民族大学青藏高原研究院,四川成都610041)
三江源区地处青藏高原腹地,是青藏高原的重要自然缓冲区,素有“中华水塔”的美誉,在中国和东亚地区的水源保护、径流调节、生态安全和生物多样性维护等方面发挥着不可替代的作用[1-2]。近年来,由于气候变化、人口急剧增加、草场牲畜超载过牧、鼠虫危害等人为和自然因素的影响,天然草地的植被盖度和初级生产力急剧降低,生物多样性锐减,优质牧草的比例降低,草地严重退化,出现了大面积“黑土型”次生裸地[3-4]。退化草地的蔓延直接影响到三江源区草地畜牧业的发展,进而影响源区草地生态系统的健康状况和经济服务功能[5]。“黑土型”退化草地原生莎草科植被层遭到了严重破坏,自然繁殖更新能力极低,仅靠草地生态系统的自然修复难以恢复至最初状态[6]。如何实现三江源区“黑土型”退化草地的恢复与重建,是世界性的生态系统恢复难题之一[7]。
为了快速、有效地恢复退化草地,建植多年生人工草地是该地区草地治理的主要措施,建植人工草地不仅能够快速恢复植被,还能提供优质高产的牧草[8]。但是随着建植年限增加,由于放牧压力大,人工管理措施不当,使得人工草地植被与土壤质量降低,群落发生逆向演替[9]。为了高效维护和持久利用人工草地,许多学者在该地区对多年生人工草地进行大量研究,但是主要集中在不同建植年限人工草地植物群落(物种多样性、生物量和群落结构)的演变过程和土壤质量(土壤理化性质和微生物)的变化规律[10-12],对地下根系的研究较为缺乏,尤其是三江源区不同建植年限人工草地根系动态特征。然而根系在生态系统中占有较大比重,与地上部分有着密切的联系,根系动态特征在很大程度上决定着人工草地植物利用土壤资源的效率和群落恢复演替的程度[13]。研究不同演替阶段植物根系动态特征,能揭示群落演替的一般规律及不同演替阶段生态系统的功能[14]。
根系动态是生态系统能量和养分流动的重要来源[15]。根系的生长、死亡、分解构成了植物向土壤系统输入碳和养分的主要途径[16],根系寿命则可以在区域尺度上反映植物、土壤和气候的总体特征,其长短决定了养分和碳的消耗与循环速度[17]。但是由于根系生长动态的复杂性和寿命估计方法的局限性,限制了对根系生长、死亡和寿命的深入了解[18]。利用微根管技术能够原位观测和记录根系的生产、发展和死亡的整个过程。通过获取根系长度、侧根伸长、生长深度等进行不同土壤层次根系动态观察,了解根系生长过程。通过观察根系的颜色变化,了解根系的衰老、死亡和分解,从而估计根系的寿命[19-20]。因此,本研究在三江源区不同建植年限人工草地展开根系研究试验,利用微根管技术,原位监测根系动态特征对人工草地不同建植年限的响应过程,旨在探讨退化生态系统恢复重建过程中:1)不同建植年限人工草地根系生产量、死亡量和现存量等动态特征和空间分布格局的变化;2)根系特征与植物群落特征、土壤理化性质间的相互关系。以期更深入地了解三江源区不同建植年限人工草地群落演替和养分循环过程,为三江源区退化草地的恢复与重建提供基础资料。
1 材料与方法
1.1 研究区自然概况
研究区位于青海省果洛藏族自治州玛沁县(33°34′-34°30′N,99°54′-100°29′E),平均海拔4150 m。属于典型的高原大陆性气候,无四季之分,仅冷暖季之别,冷季漫长、干燥而寒冷,暖季短暂、湿润而凉爽。温度年差较小而日差较大,年均气温-1.7 ℃,最冷月平均气温-14.8 ℃(1 月),最热月平均气温 9.8 ℃(7 月),年均降水量486.9~666.5 mm,多集中在6-9 月,年蒸发量1119.07 mm。自然植被以高寒草甸为主,土壤为高寒草甸土,无绝对无霜期,植物生长期为120~140 d。由于气候变化及人为扰动等因素,天然草地出现严重退化现象,部分地区已成为“黑土滩”,优良牧草盖度不足10%[9]。
1.2 试验样地设计
选择青海省果洛藏族自治州玛沁县大武镇军牧场建植年限分别为 5(34°28′10″N,100°21.3′01″E,海拔3740 m)、6(34°28′09″N,100°21.3′03″E,海拔 3738 m)、9(34°27′41″N,100°12′35″E,海拔 3757 m)和13 年(34°21′44.8″N,100°29′58″E,海拔3949 m)的 4 个人工草地为试验样地,4 块人工草地均为垂穗披碱草(Elymus nu⁃tans)、冷地早熟禾(Poa crymophila)、中华羊茅(Festuca sinensis)和星星草(Puccinellia tenuiflora)混播地,比例为4∶1∶0.5∶0.5。在每个试验样地内随机选取一个面积为50 m×50 m 的样方,于2013 年8 月在每个样方内,随机选取4 个地上植被相对均匀的区域,在其下方安装一根聚碳酸酯材料微根管,微根管与地面夹角为30°,长100 cm,内径 5 cm,外径 6 cm[21]。
1.3 样品采集与指标测定
1.3.1 植物样品采集与测定 2015 年8 月中旬,在4 块样地内分别随机选择4 个50 cm×50 cm 的小样方,进行植物群落调查。记录小样方内植物物种数。植被调查结束后,将地上植被齐平地面刈割,装进信封,并做好标记,带回实验室65 ℃烘干至恒重,并称重。
1.3.2 土壤样品采集与测定 2015 年8 月中旬,在上述调查植物群落的样方内,用内径5 cm 的土钻分两个土层(0~10 cm 和10~20 cm)按“S”形钻取5 钻,分别混合为1 个土壤样品,并做好标记。样品风干后过0.15 mm 筛用于测定土壤有机碳(soil organic carbon,SOC)、全氮(total nitrogen,TN)、速效氮(available nitrogen,AN)、全磷(total phosphorus,TP)、速效磷(available phosphorus,AP)、全钾(total potassium,TK)、速效钾(available po⁃tassium,AK)含量和pH。采用重铬酸钾氧化滴定法测定SOC 含量,凯氏定氮法测定TN 含量,康惠法测定AN含量,钼锑抗比色法测定TP 含量,碳酸氢钠浸提-钼锑抗比色法测定AP 含量,原子吸收分光光度法测定TK 含量,四苯硼钠法测定AK 含量,电极法测定pH,用烘干法测定土壤含水率(soil moisture,SM)[22]。
1.3.3 根系图片的采集与分析 于2015 和2016 年植被生长季(5-9 月)对实验区的微根管进行连续取样,首先拆开微根管外包裹的塑料袋,打开顶端盖子,使微根管内部的湿气充分与外部气流交换,让管子内外的温差平衡,然后用探杆检查管底是否有积水,并擦拭管子内壁露水,最后确认取样的初始位置,将摄像机缓慢放入微根管内,连接电脑采用CI-600 根系扫描仪(CID Bio-Science,Camas,WA,USA)进行图像采集。每根微根管按垂直土层深度分为0~10 cm 和10~20 cm 土层分别采集图像数据,每次测定结束后,将露出地面的根管按第一次的方法包裹好,每个月采集一次图像,共取10 次图像,总共追踪标记到9884 条根系。
采用WinRHZIO Tron MF(CID Bio-Science,Camas,WA,USA)软件对CI-600 采集的图像进行分析处理,测定每张图像中根系的长度,通过图像中根系的颜色来区分活根和死根,将白色和褐色的根定义为活根,黑色和消失的根定义为死根[23]。每次图片处理都导入上次数据,识别上次的根系,同时记录新生根系,每条根有独立的根号,完整记录所有的根系从起始到死亡的长度变化。
根系寿命定义为自第一次在影像中出现时刻起到第一次被定义为死亡或消失的时间[15,24]。利用SPSS 22.0中的Kaplan-Meier 方法对根系存活率和根系平均寿命进行估测,并做出根系存活曲线[17]。
由根系图像分析得到根系的长度,对t时不存在而在t+1 时出现的所有新根根长求和,加上在t时原有根系上延长的长度和来估算根系生产量。利用t和t+1 间隔时死亡和消失掉的根长及在t+1 时出现的死根根长的总和估算根系死亡量。根系现存量则用每一时刻的活根长度来估算[25]。根系的生长量,死亡量,现存量均用单位体积根长密度表示,参照吴伊波等[26]的方法计算单位体积根长密度RLDV(mm·cm-3),利用如下公式计算:

式中:L(mm)为微根管中观察到的根长;A(cm2)为观测窗面积(图像大小);DOF(cm)为微根管到周围土壤的距离,DOF一般取 0.2~0.3 cm[27],由于草地群落的根系直径较小,计算中DOF取 0.2 cm[26]。
根系净生长速率RLDNGR(mm·cm-3·d-1)即单位时间内根系单位体积密度的净增长量,参照王孟本等[28]的方法,按以下方式计算:
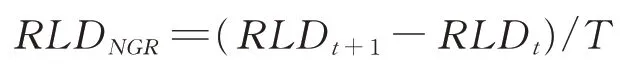
式中:RLDt+1和RLDt分别为第t+1 与t次观测中的单位体积根系现存量;T为相邻两次观测的间隔天数。RLDN⁃GR为正值说明根系的生产量大于死亡量(正生长),RLDNGR为0 表示根系的生产量等于死亡量(零生长),RLDNGR为负值表示根系的死亡量大于生产量(负生长)[28-29]。
1.4 数据统计与分析
采用SPSS 22.0 软件对数据进行统计分析,Origin 8.5 作图。利用Kaplan-Meier 方法计算根系存活率,并估计根系平均寿命。对根系特征(寿命、生产量、死亡量、现存量和净生长速率)、植物群落特征(地上生物量、丰富度)和土壤理化性质(SOC、TN、AN、TP、AP、TK、AK、SM 和 pH)进行单因素方差分析,用最小显著差数法(LSD)进行不同处理之间的显著性检验(P<0.05),不同土层(0~10 cm 和10~20 cm)之间的差异则采用独立样本t检验。采用Pearson 相关性分析植物群落特征和土壤理化性质与根系生产量、死亡量、现存量和寿命之间的相关性。利用AMOS 23.0 构建根系现存量、寿命与环境因子的结构方程模型。
2 结果与分析
2.1 植物群落特征变化
由图1 可知,随建植年限增加,植物地上生物量呈“N”字型变化,建植9 年人工草地地上生物量显著低于其他建植年限(P<0.05)。物种丰富度呈“V”字型变化,建植9~13 年显著增加(P<0.05)。

图1 不同建植年限人工草地植物群落特征变化Fig.1 Change of plant community characteristics in artificial grassland with different cultivation years
2.2 土壤理化性质变化
由表1 可知,除土壤pH 外,其余土壤理化指标均表现出0~10 cm 土层大于10~20 cm 土层。在0~10 cm 土层,随建植年限增加土壤pH 先增加后降低,逐渐趋于弱酸性;各建植年限间土壤TK 含量无显著差异,其余土壤指标均呈“N”字型变化,且各建植年限间差异显著(P<0.05)。在10~20 cm 土层,土壤pH 变化趋势与0~10 cm土层一致;随建植年限增加,土壤AN 和AK 含量显著增加;建植9 年人工草地土壤含水率、SOC 和TN 含量显著低于其他建植年限(P<0.05),土壤TP 和AP 含量显著低于建植13 年人工草地(P<0.05);建植13 年人工草地土壤TK 含量显著高于其他建植年限(P<0.05)。
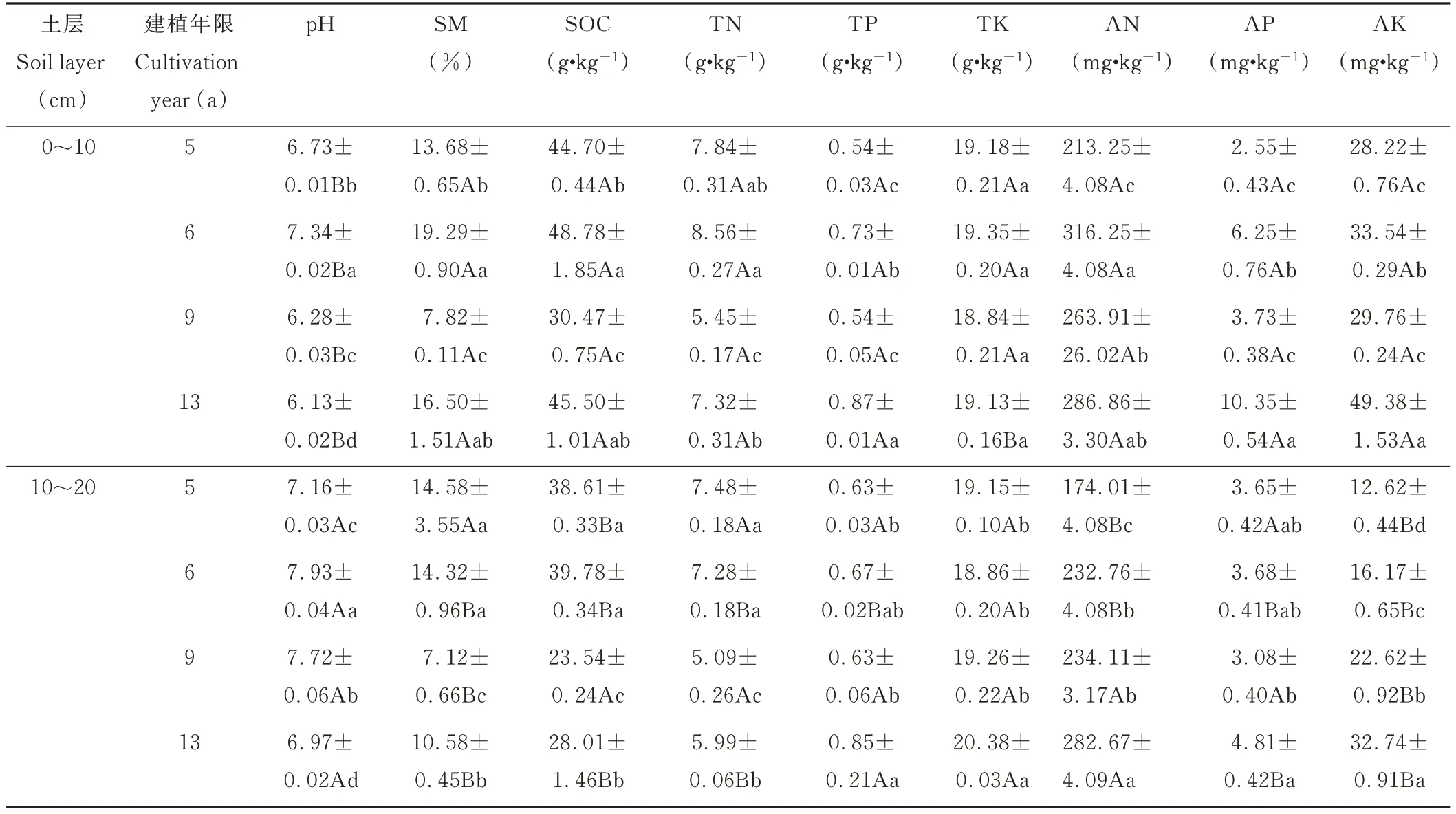
表1 不同建植年限人工草地土壤理化特征Table 1 Soil physical and chemical property in artificial grassland of different cultivation years
2.3 根系寿命变化
在0~10 cm 土层,4 个不同建植年限人工草地根系存活曲线相似,均表现为随存活时间的增加存活率降低,但建植13 年人工草地根系存活率最高,且根系平均寿命达到120 d,显著高于其他建植年限(P<0.05,图2A)。在10~20 cm 土层,根系存活曲线变化趋势与0~10 cm 土层一致,但建植6(161 d)和13 年(170 d)人工草地根系平均寿命显著高于其他建植年限(P<0.05,图2B)。

图2 不同建植年限人工草地根系存活率和平均寿命变化Fig.2 Changes in proportion of root surviving and average longevity in artificial grassland with different cultivation years
2.4 根系季节动态变化
根系生产量具有明显季节变化,在0~10 cm 土层,建植9 年人工草地2015 年持续上升,2016 年呈波动状态,7月达到生长高峰期,8 月达到生长低峰期;其余建植年限人工草地在两个生长季内均表现为先降低后升高,6 月为生长高峰期,8 月为生长低峰期(图3A)。在10~20 cm 土层,建植9 年人工草地根系生产量变化趋势与0~10 cm土层一致,其余建植年限人工草地在两个生长季内均持续下降,6 月为生长高峰期,9 月为生长低峰期(图3B)。

图3 不同建植年限人工草地根系季节动态变化Fig.3 Seasonal dynamics of the roots of artificial grassland with different cultivation years
根系死亡量也具有明显季节变化,在0~10 cm 土层,建植9 年人工草地在2016 年先降低后增加,在8 月达到死亡低峰期;其余建植年限人工草地在两个生长季均表现为先增高后降低,8 月达到死亡高峰期,此外,2016 年5月也是一个死亡高峰期(图3C)。在10~20 cm 土层,各建植年限人工草地在2015 年变化趋势与0~10 cm 土层一致(图3D)。
根系现存量在两个生长季均保持小幅度的先增加后降低的趋势,在7 月达到高峰期(图3 E,F)。0~10 cm 土层,建植13 年人工草地现存量在两个生长季均明显高于其他建植年限(图3E);10~20 cm 土层,建植5 年人工草地根系现存量在两个生长季均明显低于其他建植年限(图3F)。
不同建植年限人工草地根系净生长速率具有明显季节变化。建植5、6 和13 年的人工草地根系净生长速率均表现为先降低后增加,正负交替生长,在6 月达到正生长高峰,8 月达到负生长高峰,两个土层变化趋势相似(图4 A,B)。建植9 年人工草地0~10 cm 土层根系净生长速率先增加后降低,保持着较低水平的正生长,7 月达到正生长高峰,无负生长高峰(图4A);10~20 cm 土层,根系净生长速率2015 年与其他建植年限人工草地变化一致,2016 年先增加后降低,6 月达到负生长高峰,7 月达到正生长高峰(图4B)。

图4 不同建植年限人工草地根系净生长速率变化Fig.4 The change of net root growth rate of the artificial grassland with different cultivation years
2.5 根系累积生产量、死亡量和平均现存量的变化
图5 表示了两个生长季总根系累积生产量、累积死亡量和平均现存量的变化。随建植年限增加,两个土层根系累积生产量和死亡量均波动上升,建植13 年人工草地显著高于其他建植年限(P<0.05),且各建植年限人工草地0~10 cm 土层根系累积生产量和死亡量均显著高于10~20 cm 土层(P<0.05;图5A,B);根系平均现存量随建植年限增加持续上升,且建植5 和13 年人工草地0~10 cm 土层根系平均现存量均显著高于10~20 cm 土层,建植9 年人工草地0~10 cm 土层根系平均现存显著低于10~20 cm 土层(P<0.05),建植6 年人工草地两个土层间无显著差异(P>0.05;图5C)。
2.6 植物-土壤-根系的相关性分析
由表2 可知,地上生物量与根系生产量、死亡量、现存量呈显著负相关(P<0.05);丰富度与根系特征无显著相关性。0~10 cm 土层,AP 含量与各根系特征均呈显著正相关关系(P<0.05);AK 含量与根系生产量和寿命均呈显著正相关关系(P<0.05)。10~20 cm 土层,AK 含量与根系生产量和死亡量均呈极显著正相关关系(P<0.01);AN 含量与根系生产量、死亡量和现存量均呈极显著正相关关系(P<0.01);AP 含量与根系生产量和死亡量呈显著正相关关系(P<0.05,表3)。
利用结构方程模型对地上植被、土壤理化性质及根系特征进行适配和分析,最终建立了以下结构方程模型(图 6),该模型 CHI/DF=0.540,GFI=0.944,AGFI=0.843,RMSEA<0.001,P=0.928,说明该模型能够较好地适配研究数据以及路径分析体系。结果显示,结构方程模型(structural equation model,SEM)能解释根系寿命的60%,根系现存量的28%,有机碳的73%,速效养分的87%,地上生物量的24%,丰富度的5%。土层深度和建植年限直接影响根系寿命,且极显著正相关(P<0.01),其中土层为主要影响因子;土壤速效养分是影响根系现存量的主要因子,且呈极显著正相关(P<0.01);地上生物量直接影响根系现存量,且呈显著负相关(P<0.05);其余环境因子通过影响土壤速效养分或地上生物量间接影响根系现存量。

图5 不同建植年限人工草地根系累积生产量、累积死亡量和平均现存量变化Fig. 5 The changes of cumulative root production,cumulative root mortality and mean root standing crop of artificial grassland with different cultivation years

表2 根系特征与植物群落特征间的相关性Table 2 Correlation between root characteristics and plant community characteristics

表3 根系特征与土壤理化性质间的相关性Table 3 Correlation between root characteristics and soil physical and chemical property
3 讨论
3.1 不同建植年限人工草地植物群落特征变化

图6 根系寿命和现存量与环境因子结构方程模型Fig.6 The structural equation model of root longevity and standing crop and environmental factor
植物群落能够利用光合作用将无机碳转化为有机碳,为生态系统提供能量来源,在“黑土滩”建植人工草地,相当于引入新的植被,不仅能够提高生态系统生产力,还可以通过植物残体和枯枝落叶为土壤系统输入更多的有机物质,改善土壤质量[30-31]。本研究发现植物地上生物量呈“N”字型变化,建植5~6 年,土壤养分充足,物种间竞争较小,禾本科类植物植株较大,迅速占领较高的生态位,在整个群落中处于优势地位,因而地上生物量也较大[32];建植6~9 年土壤肥力下降,在养分匮乏时植物趋向于将资源分配给地下根系,以提高对有限土壤资源的吸收利用效率,且在资源胁迫下生长的植株比在有利条件下生长的植株个体更小[33-34];建植9 年后对资源需求少的莎草科植物逐渐取得优势地位,加之土壤养分增加,促进了地上生物量的增加。随建植年限增加,物种丰富度呈“V”字型变化,是由于人工草地的建植种在建植初期竞争能力较强,迅速占据优势地位,使得物种丰富度下降,但随着建植年限增加,原生物种会不断入侵并取得优势地位,人工草地逐渐趋于天然草地,物种丰富度显著增加。
3.2 不同建植年限人工草地土壤理化性质变化
土壤是草地生态系统营养物质的储存库,是孕育根系的主要载体[35],土壤养分、含水率和pH 是度量退化生态系统功能恢复与维持的关键指标,对植物群落的演替起着重要作用[36]。本研究发现,0~10 cm 土层土壤养分含量高于10~20 cm 土层,pH 低于10~20 cm 土层。这是由于高寒草地植物生长季短,不足以使根系向深层延伸,导致根系分布浅层化,根系死亡分解转化成有机质和其他养分,促进0~10 cm 土层养分累积[37-38],同时牲畜的踩踏使得土壤孔隙减少,土壤容重增加,阻碍了养分和水分下渗[39];此外,0~10 cm 土层植物枯落物等有机物质的累积会产生有机酸,导致土壤pH 下降[40]。建植6~9 年土壤养分含量明显下降,而在9 年后又显著增加,这是由于建植6~9 年无外源肥料的施入,加之毒杂草的侵入,土壤养分不足以弥补植物生长的消耗,造成土壤养分和含水率显著降低[41];建植9~10 年后,由于杂类草的大量入侵和繁衍,人工草地植物群落结构发生了较大改变,毒杂草比例增加,牧草品质和草地利用率大幅度下降,放牧干扰减小到了较低程度[42],同时克隆植物形成密集的匍匐茎网状结构,增加了植被覆盖度,减少了土壤水分的蒸发,枯枝落叶等向土壤输入的有机质也随之增加,从而促进了养分的累积和水分的保持[43],为一些原生莎草科植被的幼苗成功更新和补充提供良好的条件,与此同时也开始了人工草地植物群落的自然恢复演替过程。
3.3 不同建植年限人工草地根系特征变化
根系寿命是指根系从出生到死亡的这段时间,是决定植物碳通过根系更替进入土壤的关键,进而影响到植物以及整个生态系统的碳、养分和水分循环[44]。相关研究表明,根系寿命与土壤养分以及地上部分向根系供应碳含量呈正相关[45]。本研究中,建植5~6 年人工草地土壤养分含量呈上升趋势,根系吸收的养分增多,地上植被生长较旺盛,从而增加对根系的碳分配,根系寿命也随之增加。建植6~9 年人工草地处于二次退化阶段,土壤养分含量下降,使得根系吸收能力减弱,不足以满足地上植被生长,导致分配给根系的碳减少,根系衰老进而死亡,寿命降低,从而加速根系周转,增加根系对水分和营养物质的利用效率,同时可以将大量碳和养分带进土壤,促进微生物活动,改善土壤的理化条件[46]。
根系累积生产量、累积死亡量和平均现存量反映了植物根系吸收水分和养分的潜力,植物根系总长度越大,对养分和水分的利用率就越高[13]。本研究发现,随建植年限增加,根系累积生产量、死亡量均呈“N”字型变化,与养分变化趋势一致,这是因为土壤养分充足时,根系代谢活动较旺盛,从而促进根系的生长和死亡。此外,根系累积生产量、死亡量主要集中在0~10 cm 土层,这是由于随土层深度增加,土壤温度、含水率、养分和通气条件逐渐恶劣,植物为了适应不良环境采用减少根系生产和死亡的策略,从而降低自身养分消耗[21]。建植6~9 年根系平均现存量趋于深层化,是由于深根系的杂类草大量入侵,加之养分含量逐渐减小,不能够满足植物生长需要,植物为了更多地获取有效资源,采取增加深层根系的生长对策[47]。建植9 年后根系平均现存量又逐渐趋于浅层化,是因为6~9 年深层根系的增加,改变了土壤的通透性,为浅根系的莎草科植物根系生长提供了空间和资源上的生态位,从而开始了莎草科植物逐步入侵的恢复演替初期阶段。
根系对环境条件的变化反应十分敏感,常随季节呈现规律性变化[48]。根系季节动态特征直接反映了植物生长状况、土壤理化性质和营养物质等特征的变化[49]。在生长初期(6 月),根系生长量达到高峰期,这是由于植物生长旺季还没有到来,植物会优先将养分分配给根系生长,储存足够的养分,以满足植物生长旺季对养分的需求[50]。建植9 年人工草地净生长速率正生长高峰期迟于其他建植年限,这是由于根系生长的季节动态还与土壤水分的季节动态规律一致[51],该地区雨季出现在6-9 月,而建植9 年人工草地土壤水分含量较低,因此达到根系最适生长水分所需时间较长。根系现存量在7 月达到峰值,主要是由于该阶段土壤养分有效性高、温湿条件较好,根系处于正生长且死亡量较小[52-53]。8 月是高寒草甸植被生长的最佳季节,更多的养分分配到地上植被,而分配到地下根系生长的绝对量减少,增加了根系死亡量,达到死亡高峰期。
3.4 不同建植年限人工草地植物-土壤-根系的相关性
根系是连接土壤和植物的动态界面,为适应植物群落特征和土壤理化性质变化,根系特征会相应地改变,进而反馈于群落演替,影响人工草地恢复进程[14]。本研究中,土层深度是影响根系寿命的主要因子,这是由于0~10 cm 土层温度和含水率较高,根系代谢活力较强,根系周转较快,从而缩短了寿命[54]。地上生物量直接影响根系现存量且呈显著负相关,这是由于建植5~13 年人工草地还没有完全恢复到自然状态,为了适应高寒环境,随着建植年限增加植物根系会迅速积累[32],而演替过程中由于优势种的更替,人工草地建植种与原生物种间竞争激烈,地上生物量有略微下降的趋势。其余环境因子均通过速效养分间接影响根系现存量,且养分资源越丰富根系现存量越大,这是由于在自然生态系统中,植物要面对各种环境胁迫,为了保证最佳的养分吸收效率,并储存足够的养分应对不良的环境条件,在土壤中养分充足时,植物会采取增加根系现存量的生长策略[13]。相关研究表明,根系现存量越大其根系活力和竞争能力也就越强,在养分吸收方面就越具有优势,抵抗不良环境的能力也就越强[55],因此建植人工草地能够增强植物群落对不良环境的适应能力,有利于退化草地的恢复。
4 结论
1)建植5~6 年,人工管理措施合理,人工草地建植种处于优势地位,土壤养分含量较高,根系累积生产量和死亡量显著增加,根系代谢活力旺盛,抵抗不良环境的能力也较强。
2)建植6~9 年,人工草地出现了退化现象,土壤养分和水分资源可利用率低,植物减少对地上生物量的分配,增加10~20 cm 根系,以增加对水分和养分的利用效率,并降低根系寿命,加速根系周转来补充土壤养分含量,为新根生长提供养分。
3)建植9 年后,地上生物量、丰富度和土壤养分增加,根系分布趋于浅层化,且根系现存量和寿命均显著增加,以储存更多的营养物质,促进人工草地逐步向正向演替进行。
4)土层深度是影响根系寿命的主要因子,根系现存量与土壤理化性质、植物群落特征之间的反馈关系促进了人工草地生态系统结构、功能的恢复和完善。
