Genetic Interaction of Hd1 with Ghd7, DTH8 and Hd2 Largely Determines Eco-Geographical Adaption of Rice Varieties in Southern China
2021-03-20ZhangZhenhuaZhuYujunWangShilinFanYeyangZhuangJieyun
Zhang Zhenhua, Zhu Yujun, Wang Shilin, Fan Yeyang, Zhuang Jieyun
Letter
Genetic Interaction ofwith,andLargely Determines Eco-Geographical Adaption of Rice Varieties in Southern China
Zhang Zhenhua, Zhu Yujun, Wang Shilin, Fan Yeyang, Zhuang Jieyun
()
Although cultivated rice originated from the tropical region, a long process of domestication and human selection has enabled cultivated rice to grow in a wide range of geographical regions. Diversification of photoperiodic flowering provides a foundation for this diverse adaptation. Intensive studies have focused on elucidating howvarieties lost photoperiodic sensitivity (PS) to expand their cultivation areas to high latitudes, where rice is grown in a short season when day-length is long (Fujino et al, 2013; Naranjo et al, 2014; Gomez-Ariza et al, 2015; Li et al, 2015, 2018; Goretti et al, 2017; Ye et al, 2018). By contrast, limited attention has been paid to the genetic architecture of heading date (HD) variation among varieties in middle and low latitudes. The southern China rice region, located in middle and low latitudes, occupies the most important rice cultivation region in China. This region is predominantly planted withvarieties, which presents a rich diversity of regional and seasonal adaptations. In the present study, improved varieties and landraces used in this region were analyzed for allelic variations of 12 cloned QTLs controlling HD, as well as for the genotypic effects of these genes on HD and PS. Our objective was to clarify predominant genetic factors influencing eco- geographical adaption of rice varieties in southern China by comparing improved varieties with landraces.
A total of 224varieties were tested, including 132 improved varieties and 92 landraces (Tables S1 and S2). They were grown under natural short-day (NSD) conditions in Lingshui (LS), Hainan, and under natural long-day (NLD) conditions in Hangzhou (HZ), Zhejiang, China. The rice varieties were genotyped using gene-specific markers of 12 flowering genes (Table S3). Nine flowering genes showed allelic variations (Fig. 1-A). Five of them,,,,and, had two allelic types. The other four,,,and, had multiple allelic types. A single genotype was detected for the remaining three flowering genes, including,and.
The nine genes having different allelic types were analyzed for their effects on HD in HZ, HD in LS, and PS between HZ and LS. Among the five genes having two allelic types, significant genotypic effects were detected at,and, but not atandby using one-way analysis of variance (ANOVA) (Table S4). For HD in HZ that was the primary target trait of this study,andshowed differences between the two collections (landrace and improved variety). At, the effect was only significant in landrace, explaining 11.0% of phenotypic variance. At, the effect was only significant in improved variety, explaining 11.1% of phenotypic variance.
For the four genes having multiple allelic types,,,and, the Duncan’s multiple test was firstly performed, and the results were applied to re-classify the varieties into two groups. At, different non-functional types of the same collection showed no significant difference on all the three traits, but they differed significantly from the functional counterpart in LS (Table S5). Thus, different non-functional types were combined as a single non-functional group (Group 2) and the functional type remained as Group 1. Like, results of Duncan’s multiple tests forandmatched the classification of non-functional and functional types according to previous reports (Table S5). Therefore, the two genes were also classified into functional group (Group 1) and non-functional group (Group 2). For, results of Duncan’s multiple tests did not match the original classification of functional and non-functional types. In both collections, phenotypic differences were more distinguishable between the two non-functional types than between non- functional-2 and the functional type (Fig. S1). Thus, the functional type and non-functional-2 were combined as the functional group (Group 1), and non-functional-1 remained as Group 2.
Based on the two-group’s classification, effects of,,andwere determined with one-way ANOVA. All the four genes showed important effects, among which differences between the two collections were observed (Table S4). At, the early-heading group of improved variety was the functional type in both locations, but that of landrace changed from the non-functional type in HZ to the functional type in LS. Phenotypic variances explained (2) in improved variety and landrace were 8.2% and 13.3% in HZ and 69.5% and 27.9% in LS, respectively. At, major effects on HD were detected in improved variety with2of 42.3% and 55.6% in HZ and LS, respectively. However, the effects were non-significant in landrace, which may be partly caused by the unbalanced distribution of 85 functional and 7 non-functional cultivars. Atand, flowering in HZ was significantly promoted in the functional group compared with the non-functional group in each collection, with2ranging from 23.4% to 46.7%. In LS, the effects ofandwere only significant in landrace and improved variety, respectively.

Fig. 1. Allelic variations of nine genes and genetic interactions amongand,andin improved variety (ImV) and landrace (Lr).
A, Allelic variation of nine genes. LE, Low expression; HE, High expression; D, Defective; F, Functional;LH, Late heading; EH, Early heading; N, Non-functional; W, Weak; S1, Strong 1; S2, Strong 2; N1, Non-functional-1; N2, Non-functional-2; N3, Non-functional-3.
B, Heading date (HD) in Hangzhou (HZ) of genotypic groups in rice classified based on combinations ofand each of,and. Data are presented in Mean ± SD. Different capital letters indicate significant differences (< 0.01) among mean values based on the Duncan’s multiple range test.
C, Accumulation of the interaction of functional,andwith. PS index, Photoperiodic sensitivity index; NNN, FNN, FFN and FFF indicate the presence of functional genotypes at none, one, two and three of the,andloci, respectively. Data are presented in Mean ± SD. Values above the bars indicate number of varieties.
It has been known that,andare three main regulators of(Nemoto et al, 2016; Du et al, 2017; Zhang et al, 2019). Following the two-group classification of functional and non-functional genotypes at the four loci, each rice collection used in the present study was classified into four genotypic groups for gene pairs-,-and-, respectively. Phenotypic differences on HD in HZ among the four genotypic groups of the same series were analyzed using the Duncan’s multiple test. The differences revealed a consistent pattern of digenic interaction across all the three gene pairs in the two collections (Fig. 1-B). The functionaldelayed flowering with functional,and, but promoted flowering with non-functional,and. It would be also important to determine the joint effects of the three digenic interactions on the function of.
Rice cultivars in each collection were further classified into different groups according to the functional type ofand the total number of genes showing functional genotypes at the,andloci (Fig. 1-C). In improved variety, only seven groups were available because no cultivar had non- functional genotypes at all the four loci. In landrace, all the eight groups were available. In each collection, the effects ofon HD in HZ and PS index were analyzed with changes of the total number of functional,and. In improved variety, functionaldelayed flowering in HZ by 53 d when,andall had functional genotypes, but this effect almost completely disappeared when only one or two of the three background genes had functional genotypes. Consistent results were observed on PS index despite of the presence of a residual effect when two background genes had functional genotypes. In landrace, similar but less obvious tendency was detected for the two traits.
Ecotypes of the improved varieties and landraces were defined by the primary cropping region and season, followed by classification of functional groups according to the functional type ofand the total number of functional,and(Table 1). In improved variety, the seven functional groups showed a clear differentiation on growing seasons. In the group having functionaland non-functional,and, all the 55 varieties were early-season varieties. In the groups having functionalwith two or three other functional genes, all the 15 varieties were late-season varieties. In the three other groups having one or two functional genes, the 45 varieties were all early- or middle-season varieties, except one late-season variety used in central China. The last group, which had non-functionaland functional,and, contained five, three and two early-, middle- and late-season varieties, respectively. In landrace variety, differentiation of the functional groups on growing seasons was also observed but less distinguishable than in improved variety. Landraces having functional genotypic at all the,andloci were found in 29 of 31 late-season cultivars, but only in 4 of 50 early/middle cultivars.
Double cropping of early- and late-season rice varieties in the southern China rice region plays a critical role in ensuring the food security in China. It has been recognized that early-season varieties used in a large scale of the commercial rice production do not show sensibility to photoperiod or only have weak sensitivity, although they generally carry functionalhaving a strong sensitivity to photoperiod (Lin et al, 2000; Xu et al, 2010). Of the 86 early-season improved varieties for which genotypes of,,andwere all clear, 62 carried functional(Table 1). However, all the 62 varieties were not included either in the only group showing strong PS in the two collections,+FFF (functionalwith functional genotypic at all the,andloci), or in the group having residual PS in improved variety,+FFN (functionalwith two functional genotypes at the,andloci). Obviously, loss of the strong photoperiodic sensitivity of functionalin early-season rice varieties can be ascribed to the absence of sufficient complementation from,and. In the meantime, presence of the functionalin most early-season rice varieties was beneficial due to its favorable association with early heading in the presence of non-functional,and. It was also found 55 of the 62 early-season improved varieties were included in the+NNN group (functionalwith non-functional genotype at the,andloci) which showed no PS, whereas only two landraces belonged to this group (Table 1). These results suggest that the combination of functionalwith non-functional,andhas undergone intensive artificial selection in modern rice breeding.
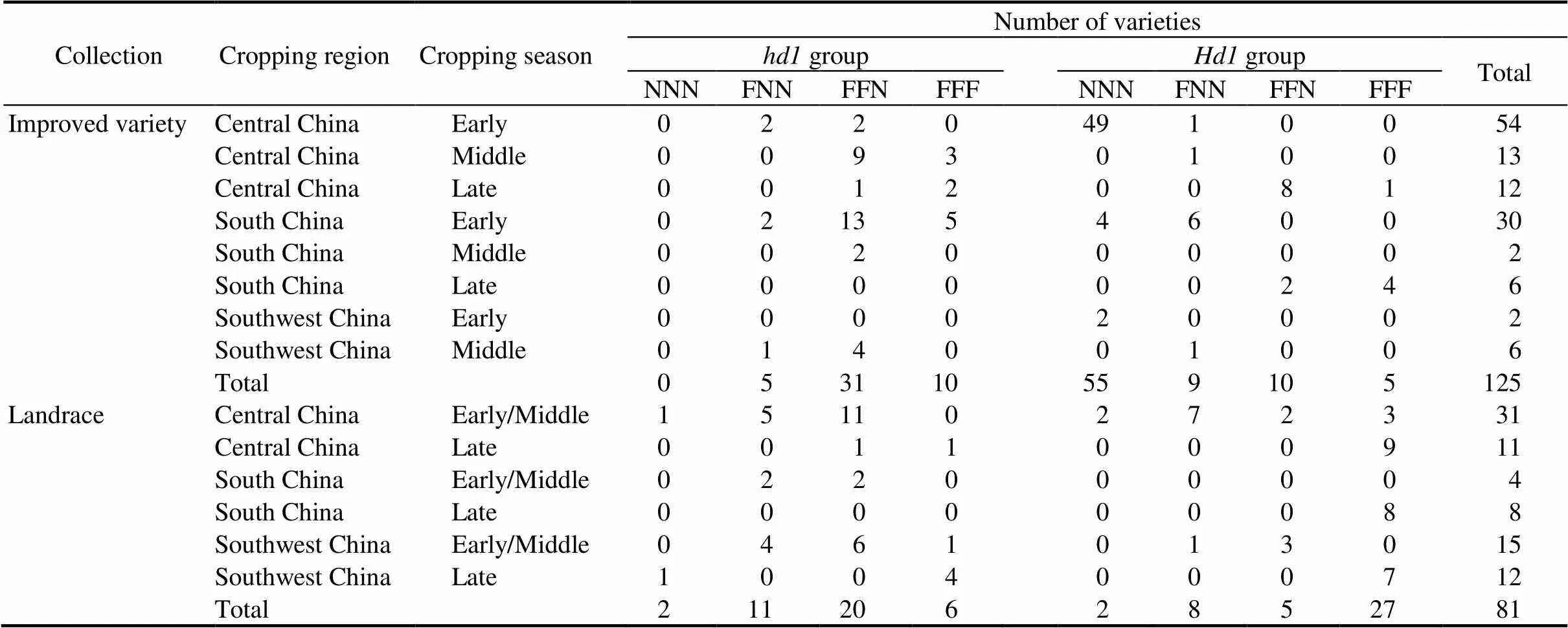
Table 1.Ecotypic distribution of different combinations of Hd1, Ghd7, DTH8 and Hd2 functional types.
, Functional;, Non-functional. NNN, FNN, FFN and FFF indicate the presence of functional genotypes at none, one, two and three of the,andloci, respectively.
Taking advantage of the climate conditions in winter and spring in Hainan Province of China, shuttle breeding from other regions of China is implemented in Hainan during the cold season (Zhou et al, 2014). The rice materials are grown and tested in Hainan under NSD conditions, but most of the varieties developed are to be used under NLD conditions. Such a difference may lead to inappropriate selection on the agronomical traits especially flowering time (Wang et al, 2001; Qin et al, 2016; Zhao et al, 2017). Based on the comparison between HD of the 132 improved varieties in HZ and LS, it appears that day-length difference between the two regions may be no longer a great problem. As shown in Fig. S2-A, discrepancy on HD between the two regions was only observed in five varieties in the+FFF group and several varieties in the+FFN group, and all of them are late-season varieties grown under NLD conditions in their original cultivating areas. On the other hand, discrepancy was observed for many more landraces (Fig. S2-B). These results indicated that intensive artificial selection in modern rice breeding not only shaped rice varieties to adapt to the eco-geographical conditions in southern China, but also brought about the adaptation to shuttle breeding between Hainan and other regions of China.
In conclusion, the effects ofconditioned by its interactions with,andlargely determined the eco-geographical adaptation of rice varieties in southern China. Intensive artificial selection in modern rice breeding has resulted in the accumulation of the genotypic combination of functionalwith non-functional,andin early-season varieties in southern China.
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation of China (Grant No. 31571637) and the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Science (CAAS-ASTIP-2013-CNRRI). We thank the National Medium Rice Genebank at the China National Rice Research Institute for providing rice varieties.
SUPPLEMENTAL DATA
The following materials are available in the online version of this article at http://www.sciencedirect.com/science/journal/ rice-science; http://www.ricescience.org.
File S1. Methods.
Fig. S1. Heading date and photoperiodic sensitivity index of three allelic types ofin rice.
Fig. S2. Comparison of heading date between Hangzhou and Lingshui.
Table S1. Improved varieties ofrice used in this study.
Table S2. Landraces ofrice used in this study.
Table S3. DNA markers used in this study.
Table S4. Heading date and photoperiodic sensitivity of multiple allelic types ofrice at,and.
Du A P, Tian W, Wei M H, Yan W, He H, Zhou D, Huang X, Li S G, Ouyang X H. 2017. The DTH8-Hd1 module mediates day-length- dependent regulation of rice flowering., 10(7): 948–961.
Fujino K, Yamanouchi U, Yano M. 2013. Roles of thegene controlling heading date for adaptation to the northern limits of rice cultivation., 126(3): 611–618.
Gomez-Ariza J, Galbiati F, Goretti D, Brambilla V, Shrestha R, Pappolla A, Courtois B, Fornara F. 2015. Loss of floral repressor function adapts rice to higher latitudes in Europe., 66(7): 2027–2039.
Goretti D, Martignago D, Landini M, Brambilla V, Gómez-Ariza J, Gnesutta N, Galbiati F, Collani S, Takagi H, Terauchi R, Mantovani R, Fornara F. 2017. Transcriptional and post-transcriptional mechanisms limit heading date 1 (Hd1) function to adapt rice to high latitudes., 13(7): e1006530.
Li X F, Liu H Z, Wang M Q, Liu H L, Tian X J, Zhou W J, Lu T X, Wang Z Y, Chu C C, Fang J, Bu Q Y. 2015. Combinations ofandgenes determine rice adaptability to Heilongjiang Province, northern limit of China., 57(8): 698–707.
Li X F, Sun Y Q, Tian X J, Ren Y K, Tang J Q, Wang Z Y, Cheng Y Q, Bu Q Y. 2018. Comprehensive identification of major flowering time genes and their combinations, which determined rice distribution in Northeast China., 84(3): 593–602.
Lin H X, Yamamoto T, Sasaki T, Yano M. 2000. Characterization and detection of epistatic interactions of 3 QTLs,,, and, controlling heading date in rice using nearly isogenic lines., 101(7): 1021–1028.
Naranjo L, Talon M, Domingo C. 2014. Diversity of floral regulatory genes ofrice cultivated at northern latitudes., 15: 101.
Nemoto Y, Nonoue Y, Yano M, Izawa T. 2016., aortholog in rice, functions as anrepressor through interaction with monocot-specific CCT-domain protein Ghd7., 86(3): 221–233.
Qin G, Ma Z F, Qing Y Y, Liu C, Zhang Y X, Huang D H. 2016. Effect of changes of ecological condition of plant breeding base of Hainan on yield related traits ofrice., 29(1): 1–5. (in Chinese with English abstract)
Wang J L, Li J Q, Zhang H, Xu W, Meng Q. 2001. The performance northernrice in Hainan and field managements., 32(2): 83–88. (in Chinese with English abstract)
Xu J F, Wei X J, Jiang L, Lu G W, Wang H J, Zhou Z L, Wan J M. 2010. Genetic analysis of heading date of some early seasonrice cultivars and hybrid rice parent in China., 24(3): 215–222. (in Chinese with English abstract)
Ye J, Niu X J, Yang Y L, Wang S, Xu Q, Yuan X P, Yu H Y, Wang Y P, Wang S, Feng Y, Wei X H. 2018. Divergent,, andalleles control heading date and yield potential ofrice in northeast China., 9: 35.
Zhang Z Y, Zhang B, Qi F X, Wu H, Li Z X, Xing Y Z. 2019.function conversion in regulating heading is dependent on gene combinations of,, andunder long-day conditions in rice., 39: 92.
Zhao F Y, Liu C Z, Ma B, Hu J F, Tan K F, Yu K C, Wang C. 2017. Main traits change of cold regionplanted in Hainan Province., 5: 31–33. (in Chinese with English abstract)
Zhou J, Luo J M, Xu X R, Zhang Y L, Ma S, Lu H Y. 2014. Strategic significance and development tend of Nanfan base construction., 38(3): 45–48. (in Chinese with English abstract)
17 March 2020;
5 August 2020
Copyright © 2021, China National Rice Research Institute. Hosting by Elsevier B V
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/)
Peer review under responsibility of China National Rice Research Institute
http://dx.doi.org/10.1016/j.rsci.2021.01.001
Zhuang Jieyun (zhuangjieyun@caas.cn)
杂志排行

Rice Science的其它文章
- Drought Tolerance in Rice: Focus on Recent Mechanisms and Approaches
- Genome Editing Strategies Towards Enhancement of Rice Disease Resistance
- RAVL1 Activates IDD3 to Negatively Regulate Rice Resistance to Sheath Blight Disease
- Osa-miR439 Negatively Regulates Rice Immunity Against Magnaporthe oryzae
- Exogenous Peroxidase Mitigates Cadmium Toxicity, Enhances Rhizobial Population and Lowers Root Knot Formation in Rice Seedlings
- Effects of Early- and Late-Sowing on Starch Accumulation and Associated Enzyme Activities During Grain Filling Stage in Rice