人类与猕猴大脑的同源脑区研究综述
2021-03-19王千山柴静文李斌强李海芳
王千山,王 玥,柴静文,李斌强,李海芳
(太原理工大学 信息与计算机学院,太原 030024)
理解大脑结构、发掘大脑工作模式,对解构人类思维、意识、认知等方面具有重大意义。探索大脑的结构和功能是21世纪最具挑战性的前沿学科[1]。2013年美国和欧盟分别提出BRAIN和HBP大脑研究计划,日本于2014年启动了“脑智计划”,我国在《国家中长期科学和技术发展规划纲要(2006-2020)》将“脑科学与认知”列入基础研究8个科学前沿问题之一。大脑研究计划的重要目的是揭示大脑的工作原理和疾病发生机制,带动相关学科及其上下游产业的共同发展。
人体为对象的各类实验受到严格的限制。国际通行的《纽伦堡法典》《赫尔辛基宣言》以及我国《涉及人的生物医学研究伦理审査办法(试行)》中都对人体实验设置了各类规范和限制。最新颁布的《民法典》中对人体实验提出明确的法律规定。在开展人体实验的过程中,不仅要关注伦理问题,实验对象的生命安全更应放在首位。因此侵入式的创伤性实验的开展,要经过反复论证,并在动物模型上试验通过后再评估能否进行人体实验。而且考虑到示踪药物对于人体的影响,诸如CT、PET等非侵入式的实验也不能在受试者上反复进行。此外,民众普遍对辐射较为敏感,因此即便是对人体影响最小的磁共振技术,受试者往往也不愿配合进行多次实验。人体实验难以开展的情况很大程度上限制了脑科学的发展。
动物大脑研究是研究人类大脑的重要途径,借助动物大脑可以研究人类大脑的工作机制和脑疾病的发展规律。猕猴在中国具有得天独厚的资源优势,随着体细胞克隆猴和转基因猕猴模型的相继突破,猕猴作为焦点模式实验动物的地位进一步强化。虽然非人灵长类动物具有和人类相似的大脑组织结构和保守特征,但在生物演化过程中仍出现了形态、连接、功能等方面的差异[2]。因此将动物实验的结果推广到人类还需要进一步研判。
同源脑区是指具有相同演化先祖、拥有相近或相同功能的脑区,能够为跨物种研究提供相对可靠的推测依据,为大脑组织方式研究提供独特解读依据[2]。现代医学研究得到的人类和猕猴已知脑图谱对于大脑区域的划分多达数百个,但尚不存在某两个图谱能够将人脑和猕猴脑区完整对应。且目前人类与猕猴的大脑同源脑区研究中,缺少对已证明的同源脑区的全面总结和综合研究记录。对同源脑区进行梳理总结对制作精确的人类和猕猴脑区映射关系图谱、探究大脑工作机制和脑疾病的发展规律具有重大意义。
本文收集整理2000年以来涉及人类和猕猴同源脑区选取的相关研究资料。寻找大脑分区数目明确、适用于计算机分析、有望为人类和猕猴大脑同源区研究提供较好帮助的脑图谱或脑模板;选取被明确证实的同源脑区或在跨物种脑区功能研究中使用的脑区,并根据其所涉及的具体功能进行归类总结。目的是回顾同源脑区的研究过程,总结已有的同源脑区和其研究进展,为新的同源脑区的研究找到理论支撑,为大脑高级认知功能研究奠定理论基础,以期推动人脑工作机制和脑疾病发展规律的相关研究。
1 两物种脑图谱匹配
人类脑图谱发展至今数量众多,代表性的是以MNI152标准空间作为脑模板[3],先后诞生的诸如哈佛-牛津图谱[4]、AAL模板[5]、Brainnetome人脑连接组图谱[6]以及Julich脑图谱[7]。相较于人类脑图谱,猕猴脑图谱数量较少。伴随影像学的发展,非人灵长类动物的脑影像数据的不断积累,基于MRI的非人灵长类动物的脑模板、脑图谱陆续被提出。特别是如PRIME-DE此类项目,提供了大量的多平台多中心的猕猴非人灵长类动物脑影像数据集[8]。
由于人类和猕猴大脑间已探知的同源组织数量和精细度严重不足,因此跨物种的大脑研究仍停留在低层次脑区的表征和比较层面,这样就限制了脑区同源性研究的发展。目前,一些研究团队已开始此方面的初步探索,如华盛顿大学医学院Van Essen实验室基于形态标记约束的表面配准过程实现两个物种在形态学上的体素拓扑映射[9],牛津大学纳菲尔德脑功能磁共振成像中心尝试提供一些连接模式上存在最佳对应关系的位点[10-12]。由此可见,构建能够逼近人类和猕猴真实同源关系的大脑体素级映射关系的脑图谱有着十分重要的价值。
猕猴脑图谱研究中,主要以FOX和LANCASTER[13]、PAXINOS et al[14]、SALEEM和LOGOTHETIS[15]提出的猴脑图谱标记作为标准,为后续脑图谱开发提供重要的分区依据。此后,2009年DUBACH et al[16]绘制了NeuroMaps三维猕猴脑图谱,2011年FREY et al[17]采用人脑MNI空间构建方法,将Paxinos绘制的猕猴图谱标记配准到MNI空间从而提出猴脑MNI脑模板。基于MNI猴脑模板ROHLFING et al[18]在2012年制作了INIA19脑模板,并将NeuroMaps猕猴图谱手工绘制其上。同年,VAN ESSEN et al[9]基于表面图集,在早期D99脑模板上绘制了F99的皮质表面图谱。2015年,为更好地使用连接度量分析,Duke大学的KOLSTER et al[19]使用弥散磁共振数据,结合Paxinos猕猴图谱和MNI标准空间,绘制基于弥散数据的CIVM MRI/DTI atlas猕猴脑图谱。DONAHUE et al[20]于2016年同期发布基于弥散数据的Yerkes19猕猴脑图谱。2017年,REVELEY et al[21]在早期D99脑模板的基础上,结合前人的工作,绘制了猕猴3D脑图谱。2018年,SEIDLITZ et al[22]结合此前工作,将LOGOTHETIS绘制的图谱中248个脑区配准其上发布了NMT脑图谱。随后JUNG et al[23]在其NMT的基础上引入恒河猴皮层层次图集(CHARM),制作了NMTv2脑图谱。本文检索大量脑图谱并研究其图谱属性,从而给出两类人类和猕猴大脑图谱的最佳匹配结果,两物种大脑图谱的匹配流程如图1所示。
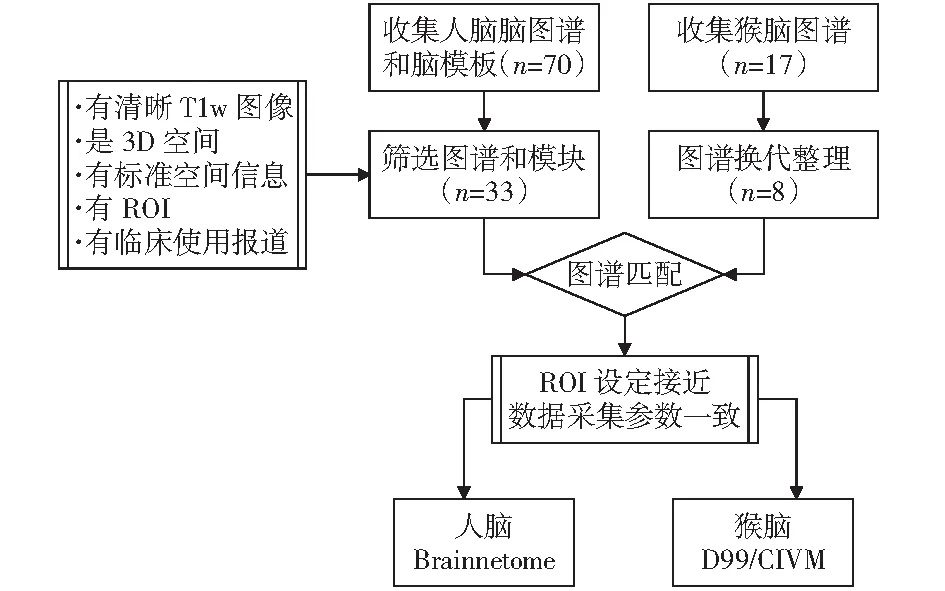
图1 选取匹配图谱
1.1 大脑皮层组织图谱
基于形态学标记约束的配准方法是构建人类和猕猴大脑对应关系的重要途径。该方法的准确性依赖于大脑皮层组织图谱的精细程度和配准软件的适应能力。在实际比对后发现,人类脑图谱在FSL、FreeSurfer、SPM等软件中,使用各自软件内置的脑图谱均可以产生较好的配准效果。在AFNI中使用D99脑模板或FreeSurfer中使用F99脑模板能够得到相对较好配准结果。通过对脑区位置和数量进行详细比对,发现人脑Brainnetome连接组图谱提供的246个区域、Julich-Brain Atlas标记的248个区域能够同2017年更新的猴脑D99脑模板的254个区域构成一定的对应关系。
1.2 弥散磁共振图谱
弥散张量成像(DTI)技术能够较好的观察大脑内部白质纤维走向。通过使用弥散磁共振技术已经围绕人脑中运动皮质区域建立起感觉运动区模板(SMAAT).围绕猴脑展开的图谱研究,目前还未有同人脑中SMAAT图谱相似的白质纤维束图谱。Brainnetome连接组图谱中提供了基于弥散态的图谱信息,其中的246个区域能够同Duke大学的CIVM MRI/DTI 图谱提供的241个区域在一定程度上形成对应关系。为猴脑计算制作全脑的连接组图谱,势必能够为构建人脑和猕猴大脑对应关系提供有力的支持。
2 已知的同源脑区
基于图论的复杂网络分析极大地推进了对于人类脑网络组织模式的认识。构建的网络中的节点数和网络复杂度,很大程度上依赖于脑图谱对于脑组织的区域划分结果[24]。特别在MARS et al[25]提出基于连接指纹图的方式进行跨物种比较后,同源脑区的选定变得更加重要。
虽然人类与非灵长类动物的大脑结构具有相似性,但在人类的进化过程中,出现了诸如大脑半球偏侧化、脑组织体积膨胀、功能转移等情况[26],导致跨物种比较中不能仅以大脑组织的宏观形态标记划分感兴趣区域,以脑区的同源性为根据,进行兴趣区域的筛选,有助于更精确地分析大脑结构和工作原理。
同源脑区的定义至今不够完善,研究者对同源组织进行过一些总结,如:华盛顿大学医学院Van Essen实验室总结利用55对大脑皮层同源脑区用于表面配准[27],牛津大学纳菲尔德脑功能磁共振成像中心预定义出22对同源脑区来比较人类和猕猴前额叶皮层位点的功能连接模式[10],随后的研究中MARS et al[28]仅通过预定义5对同源脑区研究了辅助运动区(SMA)、前辅助运动区(PreSMA)以及Area9、10等脑区的大脑同源性。中国电子科技大学生命科学与技术学院的WANG et al[29]预定义了17对同源脑区来比对人类和猕猴的PCun结构。这些研究都是带着特定目的粗略总结,详细的同源组织还需要专门系统总结。
同源区的研究主要通过电刺激效应分析、细胞构筑、解剖分析、连通性分析等四类方法开展[30-34]。如CASPERS et al[35-37]在2006-2013年间,先后受体细胞构筑、解剖分析、功能连接等方法,验证了人类和猴类大脑中顶叶部分中与认知相关的部分脑区之间的同源关系。JOHANSEN-BERG、MORECRAFT et al、SALLET et al先后使用解剖分析和细胞构筑的方法验证了人类和猴类大脑中辅助运动区(SMA)的同源关系[38-40]。MARS et al发表的基于结构连接的同源性研究方法在近年逐渐成为研究关注热点[28]。
本文根据人类大脑的一般工作过程,将同源脑区依照同感觉过程、行动过程、言语过程、认知过程四类处理过程的相关性进行分类。脑区同源关系确定过程如图2所示,通过查阅相关脑区的研究报道,筛选出至少包含3种同源区研究方法相关记载的脑区作为有效的同源脑区,为同源脑区研究提供相关的理论基础。
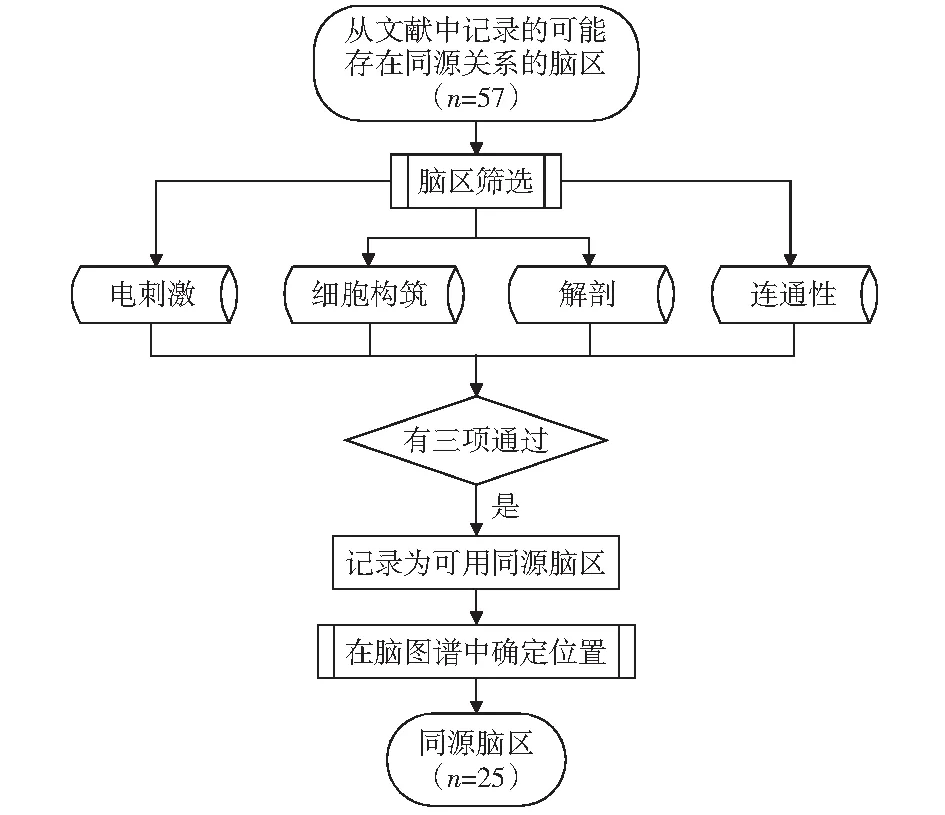
图2 同源脑区选取
2.1 感觉过程相关
目前脑区同源性研究主要集中在以视觉为代表的感觉过程相关脑区。灵长类动物通过视觉获取了大量的外部环境信息,这些信息经过加工整理,驱动人体产生各类具体反应。
1) 人类的外侧枕叶皮质(LOC)同猕猴LOC部分同源[31-34]。
2) 人类大脑中位于海马旁回附近的嗅周皮质区域(Perirhinal)同猕猴35、36脑区同源[41-43]。
3) 人类颞极(TP)皮质同猕猴TG区同源[32,43-45]。
4) 人类顶内沟腹侧(VIP)同猕猴VIP同源[45]。
5) 人类辅助运动前侧(preSMA)同猕猴的F6区域同源[25]。
2.2 动作过程相关
动作相关的同源脑区研究在众多脑区同源性研究中最早开始。动物都具有一定的行为动作能力,是区别于植物的主要特征。动作是包含表情变化和动作变化,以及由肢体动作变化导致的本体在空间上能够发生的位置变化。而动作的完成,主要是由中枢神经系统有规律的控制、协调各个肌肉群配合完成的。动作作为一种基本能力,在漫长的演化过程中得到了长期的保留,相关脑区的同源性较容易得到证实。
1) 人类和猕猴同位于前连合冠状面后面的内侧额上回的辅助运动区[38-40]。
2) 人类和猕猴的初级运动皮质区(M1)[46-47]。
3) 人类和猕猴的初级体感皮质区(S1)[38]。
2.3 言语过程相关
语言的复杂结构使得其表达范围比任何已知的动物交流系统都要广泛。语言包含视觉、听觉、触觉等多个方面,言语系统的表达虽然在不同生物间有很大差异,但依旧可以找到其表达过程的共有特征。
1) 人类额中回前部和额上沟的交界区(aMFG+SFS)与猕猴9/46区[39,48-50]。
2) 人类额上回下延至旁扣带回区域(sSFG+Cingulum)与猕猴8B/8Bm区[39,51-52]。
3) 人类额下回腹侧(vIFG)与猕猴44v区[53-56]。
2.4 认知过程相关
认知是人类获取并加工信息的过程。这一能力是人类与其他灵长类动物的主要差异。在该研究领域中,研究人员既希望找到更多的同源特性论证,以辅助药物、精神疾病研究,同时也急切的希望找到更多的差异区域,借此解开人类智慧的奥秘。因而此类区域集中了大量的研究工作。与认知相关的同源脑区有:
1) 人类顶下小叶前部和岛叶外侧区域(PFop)同猕猴大脑的PFop/7op区域[26,35-37]。
2) 人类顶下沟前部(aIPS)同猕猴大脑的顶内沟前部(AIP)[26,35-37].
3) 人类顶下沟后部(pIPS)同猕猴大脑的顶下沟后部(pIPS)[26,35-37].
4) 人类顶下小叶顶部(PG)同猕猴大脑的顶下小叶的PG/7a区[26,35-37]。
5) 人类扣带回(PC)后部同猕猴大脑扣带回后部(pPC)[38,57-59].
6) 人类扣带回皮质的后皮质区域(rsplC)同猕猴23ab区[38,57-59]。
7) 人类前扣带回膝部(pgACC)同猕猴pgACC区[10,60-61]。
除上述区域外,还有一些结构清晰、功能明确的脑区,在早期研究中得到明确同源论证结果。此类脑区有腹侧纹状体(ventrStr)、尾状核头部(Caud)、壳核(putamen)、海马体(hippoc)、杏仁核(amygd)、下丘脑(hypoth)、腹侧被盖区(VTA)[62-65].
本文梳理出的25对同源脑区在MNI标准空间中的位置如图3所示,人脑和猕猴同源脑区在图谱中名称的对应关系详见表1.
通过梳理文中涉及的同源脑区相关研究工作发现,人类Perirhinal皮质同猕猴35、36脑区的同源研究中,尚未使用纤维追踪判断连通性的方式论证其同源性;人类和猕猴的初级运动皮质区(M1)同源研究中,尽管人类已经构建了SMAAT图谱以描述人类大脑各运动单元的白质纤维连接情况,但两者的连通性分析仍未进行。人类额下回腹侧与猕猴44v区域同源研究关联到Broca区的研究分析,已在细胞构筑和连通性分析方面进行了详细论述,但功能连接分析没有进行。人类扣带回(PC)后部同猕猴大脑扣带回后部同源性研究情况也存在类似功能连接分析尚未论述的情况。
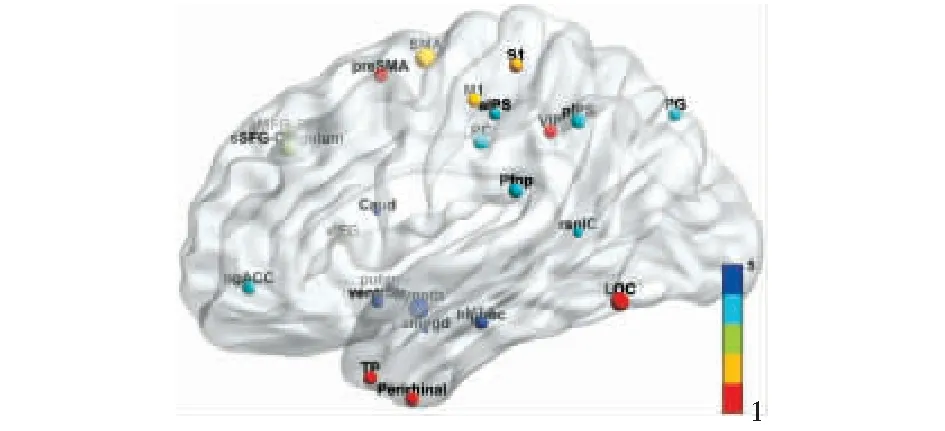
图3 同源脑区位置示意

表1 已知同源脑区的人猴对应关系表
目前仍有大量的脑区同源性有待证实。猴脑脑区在PAXINOS[14]和LOGOTHETIS et al[15]提出的猴脑图谱中均被手工标记了200个以上的大脑脑区。如认知控制和语言功能相关的45区、47/12区;社会认知和面部识别的vmPFC,14m区;对于奖惩敏感的FPm区域的研究;参与到空间注意、语言和高级运动加工的脑区如PF,PG,IPL,Broca等区域的同源性研究都有待更多的科学工作者揭示其中原理。同时,大量有望能够为药物靶点治疗提供更多可能的脑区的功能同源性分析仍亟待开展。
值得注意的是,人的动作、言语、认知过程均不是孤立发生的,各类的行为是在基于对感觉通路获得的大量信息进行有效处理后,经过复杂决策过程后进行的生理活动。特别是认知过程,要整合加工各类型信息,并在不同脑区间进行复杂的信息传递,而不是简单的在某个脑区内直接完成的。这就导致感觉通路上的各个脑区同大量其他活动相关的脑区都存在一定程度的联系。即使是对面孔的识别也存在由基本视觉区域到多个不同功能脑区的投射。因此在分析各种类型的脑区活动时,都应将感觉过程相关的同源脑区作为重要的信息输入参考引入到相关的计算和分析。
3 总结
目前国内开展的脑区同源性研究已得到很多成果,但在人类与非人灵长类动物方面还在发展阶段。多图谱的脑区识别方法能够为同源脑区研究提供更多信息,本文中提及的25对同源脑区在D99猕猴脑图谱中仅能有效的对应13个脑区,其他脑区在INIA19、CIVM图谱中予以寻找完整。
国内人类脑连接组图谱研究居国际领先地位,转基因猕猴模型制作也属前列,但仍缺乏高清的非人灵长类脑图谱。随着新型7T磁共振设备的推广[66],大量3T/7T磁共振数据势必大量产生,猕猴脑影像数的不断积累,可促使猕猴脑连接组图谱的产生,对非人灵长类图谱研究起重要推动作用。构建能够清晰反映人类和猕猴大脑体素级映射关系图谱能够有效推动大脑工作机制探究和脑疾病的研究进展。
