拟南芥LTP2参与乙烯信号转导调控的初步研究
2021-03-05张力琪李海鸥萧浪涛
张力琪,黄 珂,肖 帅,刘 清,李海鸥,萧浪涛
(湖南农业大学植物激素与生长发育湖南省重点实验室,长沙 410128)
脂质转运蛋白家族(lipid transfer proteins,LTPs)是一类富含半胱氨酸的小分子可溶性蛋白,分子量通常低于10 kD,具有4~5个α-螺旋,由8个半胱氨酸形成4个二硫键,一般形式为C-Xn-CXn-CC-Xn-CXC-Xn-C-Xn-C[1-2],其三级结构中有一个贯通的疏水腔,能可逆结合单磷脂分子和蜡质单体等多种疏水分子。脂质转运蛋白分布广泛且功能多样,如拟南芥(Arabidopsis thaliana)中的糖基磷脂酰肌醇锚定脂质转运蛋白(glycosylphosphatidylinositol-anchored lipid transfer protein,LTPG)能影响表皮蜡质积累[3-4];拟南芥中的AtLTP12参与调控花粉和种子发育[5-6];甘蓝型油菜(Brassica napusL.)中的BraLTP1过表达会导致植株异常的绿色着色、减少叶子的蜡质沉积,影响细胞分裂素合成基因IPT3高表达和花朵形态的变异[7];卷心菜(Brassica oleraceaL.)中的部分LTPs有冷冻保护素的功能,能提高卷心菜的抗寒能力[8];土豆(Solanum tuberosumL.)中的StnsLTP1过表达能显著提高其在逆境胁迫如高温、干旱与盐胁迫下的生存能力[9]。脂质转运蛋白家族功能多样,其中LTP2分子量为9.6 kD,定位于细胞壁与质膜。拟南芥ltp2突变体的黄化苗会出现蜡质层发育缺陷的表型[10];烟草(Nicotiana tabacum)中的LTP2参与细胞壁的伸长[2,11];大麦(Hordeum vulgare)中的LTP2在拟南芥中过表达能增强其对丁香假单胞菌(Pseudomonas syringae)的抗性[12]。
乙烯是一类重要的植物激素,各种植物组织在一定条件下都能产生乙烯。乙烯具有催熟果实、诱导蜡质合成、增强抗逆性等诸多功能。乙烯信号转导通路始于信号感受,即乙烯分子结合到乙烯受体蛋白1(ethylene receptor 1,ETR1)的N端,随后会解除受体与乙烯组成型三重反应蛋白1(constitutive triple-response 1,CTR1)的结合,使乙烯不敏感蛋白2(ethylene-insensitive protein 2,EIN2)去磷酸化,继而激活乙烯不敏感蛋白3(ethylene-insensitive protein 3,EIN3)与乙烯不敏感蛋白3类似蛋白(EIN3-like proteins,EILs),将乙烯信号传递到下游乙烯响应因子(ethylene response factor,ERF),从而调节下游基因表达,产生乙烯响应表型[13-14]。乙烯敏感性蛋白1(reversion-to-ethylene sensitivity 1,RTE1)蛋白合成受乙烯调控,通过占据ETR1的乙烯结合位点负调控下游的乙烯反应[15]。
使用乙烯或其前体物处理植物幼苗能诱导产生“三重反应”表型,可用于鉴定植物乙烯信号通路上的基因突变[16]。乙烯的“三重反应”是指外施乙烯或乙烯生物合成前体1-氨基环丙烷羧酸(1-aminocyclopropanecarboxylic acid,ACC)后,使植物产生顶部弯钩卷曲程度加大、下胚轴变粗变短、根伸长受到抑制的现象[17]。乙烯信号通路发生基因突变会改变其“三重反应”相关的表型,其突变体主要分为两类:一类是组成型乙烯反应突变体,通常是负调控因子突变激活了乙烯信号通路,如乙烯信号通路中的CTR1发生突变导致在无外源乙烯或ACC的条件下出现“三重反应”表型;另一类是乙烯不敏感突变体,通常是由于乙烯信号通路中的正调控因子突变,导致在外源乙烯处理下依然不发生“三重反应”[18-20]。
蜡质是植物表皮最外层的重要成分,其合成途径分为两步:质体内的C16和C18酰基链脂肪酸合成与质体外的C16或C18饱和脂肪酸延长及衍生物合成;质体外长链脂肪酸合成后,通过超长链脂肪酸经脱羰基途径和酰基还原途径,最终形成各种蜡质[21]。蜡质合成酶CER3(eceriferum 3)参与催化烷类的合成。蜡质合成酶CER6(eceriferum 6)是脂质合成中超长链脂肪酸合成的限速酶,其突变会导致拟南芥的表皮蜡质缺失[22]。研究表明,乙烯对植物表皮的蜡质合成具有重要影响,如作为乙烯响应因子的蜡质诱导蛋白1(wax inducer 1,WIN1)能调控蜡质合成并改变植物表皮渗透势[23]。ltp2突变体的黄化苗下胚轴细胞壁与蜡质层分离产生空隙[10]。虽然LTPs与乙烯均能单独影响蜡质层[10,23],但两者之间的关系尚未明确。
LTP1能通过与RTE1互作来调控乙烯信号通路,而ltp1突变体在ACC处理下会出现下胚轴比野生型更短的表型[24];ACC处理LTP1过表达株系,植株对乙烯弱敏感,过表达植株下胚轴长于野生型。虽然LTP2突变可影响拟南芥叶片表皮蜡质的形成与功能[10],但LTP2是否通过乙烯途径影响蜡质的合成目前还不清楚。为探究LTP2是否参与乙烯信号转导,本试验以拟南芥Col-0和脂质转运蛋白突变体ltp2为试验材料,分析ACC处理后两者的“三重反应”表型差异,并检测了LTP2基因、乙烯信号转导相关基因(RTE1、ETR1、CTR1、EIN2和EIN3)、乙烯响应因子基因(ERF1、ERF2和ERF9)及下游蜡质合成酶基因(CER3和CER6)的表达变化特性,初步结果表明,LTP2参与了乙烯信号转导途径。
1 材料与方法
1.1 材料
拟南芥Col-0与ltp2突变体。ltp2突变体购买于NASC(the EuropeanArabidopsisStock Centre,http://arabidopsis.info/),编号为SALK_026257。ltp2突变体属于T-DNA插入突变体,插入位点位于启动子区域,导致LTP2表达量大幅下调[10]。
1.2 主要试剂与仪器
ACC(S30902,1.00 g)购自源叶生物(http://www.shyuanye.com/goods-S30902.html);MS(Murashige and Skoog culture medium)培养基(M8521,100 L)购自北京索莱宝科技有限公司(http://solarbio.bioon.com.cn/);植物RNA小量提取试剂盒(R4151-02,50 Preps)购自美基生物科技有限公司(http://www.magentec.com.cn/);mRNA反转录试剂盒(AE311-03)购自北京全式金生物技术有限公司(http://www.transgen.com.cn/);组织破碎仪为QIANGEN TissueLyser II;RNA和DNA浓度测定使用多功能酶标仪(Tecan,SPARK);荧光定量PCR(Real-time PCR)仪为BIO-RADCFX96。
1.3 方法
1.3.1 含ACC的MS固体培养基的配制
称取ACC粉末0.05 g溶于1 L ddH2O配制成500μmol/L ACC母液,–20℃避光保存。使用时从母液中取1 mL溶液加入到100 mL的MS培养基中混匀,配制成含5μmol/L ACC的MS培养基,4℃避光保存,用于下一步试验。MS培养基含3.0%蔗糖、0.6%琼脂粉。
1.3.2 乙烯的“三重反应”验证
将Col-0与ltp2种子在75%乙醇中消毒15 min,无菌水清洗3次,悬浮于0.1%的琼脂中,于4℃低温条件下处理3 d。处理组将试验材料点种在含5μmol/L的ACC、3.0%蔗糖与0.6%琼脂的MS培养基的培养皿中;对照组培养皿中的培养基与处理组相同但不含ACC。将点种后的培养皿密封并置于22℃黑暗条件下,4 d后观察表型并计量数据。试验重复3次。
1.3.3 Real-time PCR分析
将样品收集至2 mL EP管中,加入钢珠,使用组织破碎仪研磨成粉末,过程中保持低温,防止样品解冻。使用微量RNA提取试剂盒提取样品RNA,根据RNA浓度调整反转录的RNA用量至500 ng。使用逆转录试剂盒将RNA逆转录为cDNA。所有样品根据OD260值(样品在260 nm波长处光的吸收值)将cDNA浓度调整一致,进行Real-time PCR。使用Real-time PCR检测LTP2以及乙烯信号转导通路基因(RTE1、ETR1、CTR1、EIN2和EIN3)、乙烯响应基因(ERF1、ERF2和ERF9)以及蜡质合成基因(CER3、CER6)的表达量。Real-time PCR反应体系:cDNA模板0.5μL、上游引物0.5μL、下游引物0.5 μL、SYBR Green Master 7.5 μL、ddH2O 6.0 μL,共15.0 μL。Real-time PCR反应程序:95℃预变性30 s,95℃变性10 s,60℃退火延伸30 s,共45个循环。试验重复3次。使用qPrimerDB-qPCR引物数据库网站(https://biodb.swu.edu.cn/qprimerdb/)设计引物(表1),引物由擎科生物科技有限公司合成。
1.4 数据处理
Real-time PCR数据结果使用荧光PCR仪中BIO-RAD CFX9 manager 3.1软件进行分析,使用Excel 2010进行显著性分析。
2 结果与分析
2.1 LTP2突变可弱化拟南芥黄化苗的乙烯的“三重反应”
ACC是乙烯合成的直接前体,在研究中可通过添加ACC的方式可控地调节乙烯反应,使研究结果更加准确。本研究中使用添加浓度为5μmol/L ACC的MS培养基对拟南芥野生型(wild type,WT)与ltp2种子进行处理。与对照组(control check,CK)表型进行比较,发现ACC处理后的ltp2出现乙烯弱敏感表型:顶部弯钩弯曲程度小,下胚轴与根长略长于野生型,对照组中ltp2长度与野生型无明显差异(图1、2)。试验结果表明LTP2表达量下调能影响植株对乙烯的敏感程度。
2.2 拟南芥LTP2基因表达受乙烯诱导
Ltp2突变体中LTP2的表达量大幅下降,而在ACC的处理下突变体出现了乙烯弱敏感表型。为了探究ltp2突变体产生乙烯弱敏感表型与乙烯信号通路基因之间可能存在的联系,我们对拟南芥野生型与ltp2突变体的相关基因进行了Real-time PCR检测,结果发现野生型的LTP2基因的表达量在ACC处理后上升了77.0%,而突变体中的LTP2表达量无明显变化(图3)。这一结果说明LTP2表达受到乙烯的调控。

表1 Real-time PCR 引物序列Tab. 1 Primers used for Real-time PCR analysis
2.3 LTP2突变影响乙烯信号通路基因的表达
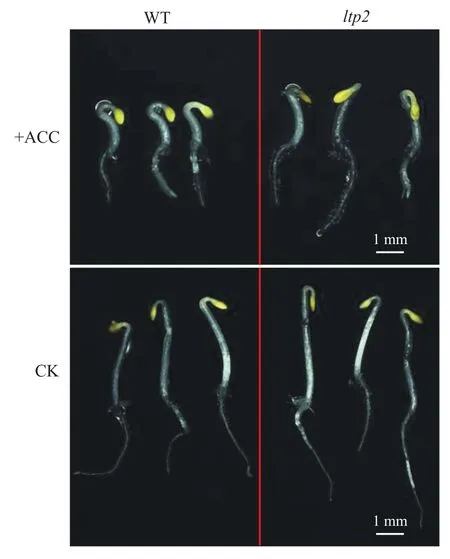
图1 ACC处理对拟南芥野生型与ltp2突变体表型的影响Fig. 1 Effect of ACC treatment on the Arabidopsis thaliana wild type and ltp2 mutant phenotype
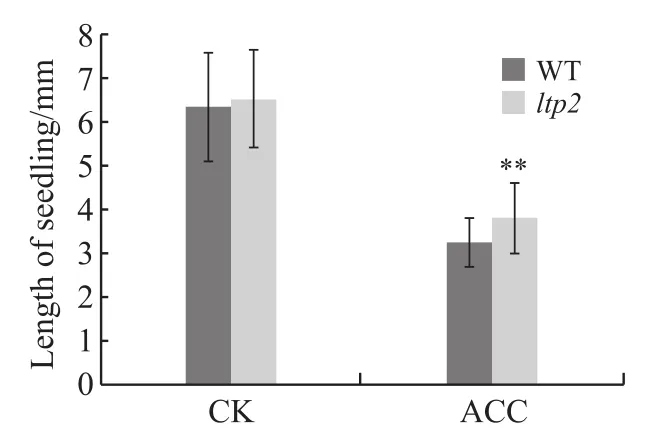
图2 ACC处理拟南芥野生型与ltp2突变体黄化幼苗长度对比Fig. 2 Comparison of relative length of the etiolated Arabidopsis thaliana wild type and ltp2 mutant with ACC treatment
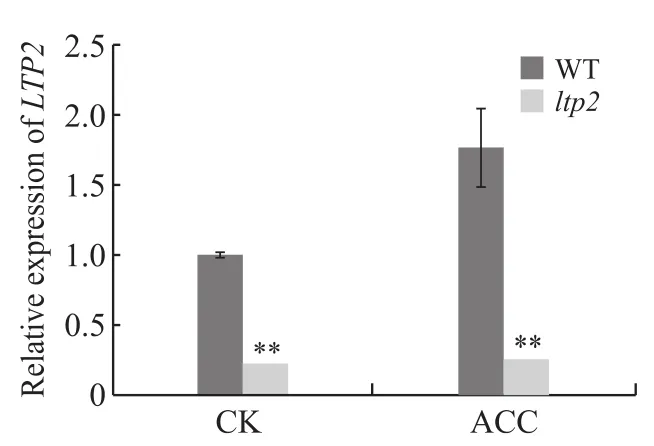
图3 拟南芥野生型与ltp2突变体的LTP2基因相对表达量Fig. 3 The relative expression of LTP2 gene in the Arabidopsis thaliana wild type and ltp2 mutant
LTP2突变会导致拟南芥材料对乙烯的敏感程度降低,从而弱化其乙烯“三重反应”表型(图1),而乙烯信号通路基因RTE1[15]、ETR1、CTR1[25]、EIN2[13]和EIN3[14]等产生突变也会引起“三重反应”表型的变化。因此,本试验使用ACC处理拟南芥野生型与ltp2材料,并分析两者乙烯信号通路相关基因表达的差异。在无ACC处理时,与野生型相比,突变体中ETR1与EIN2的表达量有显著升高,分别上升81.0%(图4b)与87.0%(图4d),而两种材料RTE1、CTR1和EIN3的表达量无明显差异(图4a、4c、4e);ACC处理组中,野生型RTE1表达量比突变体高96.0%(图4a),但野生型的CTR1的表达量比突变体低23.4%(图4c),ETR1、EIN2和EIN3表达量无明显差异(图4b、4d、4e)。根据表达量检测,LTP2可能通过影响RTE1与CTR1的表达来降低乙烯反应的敏感程度。
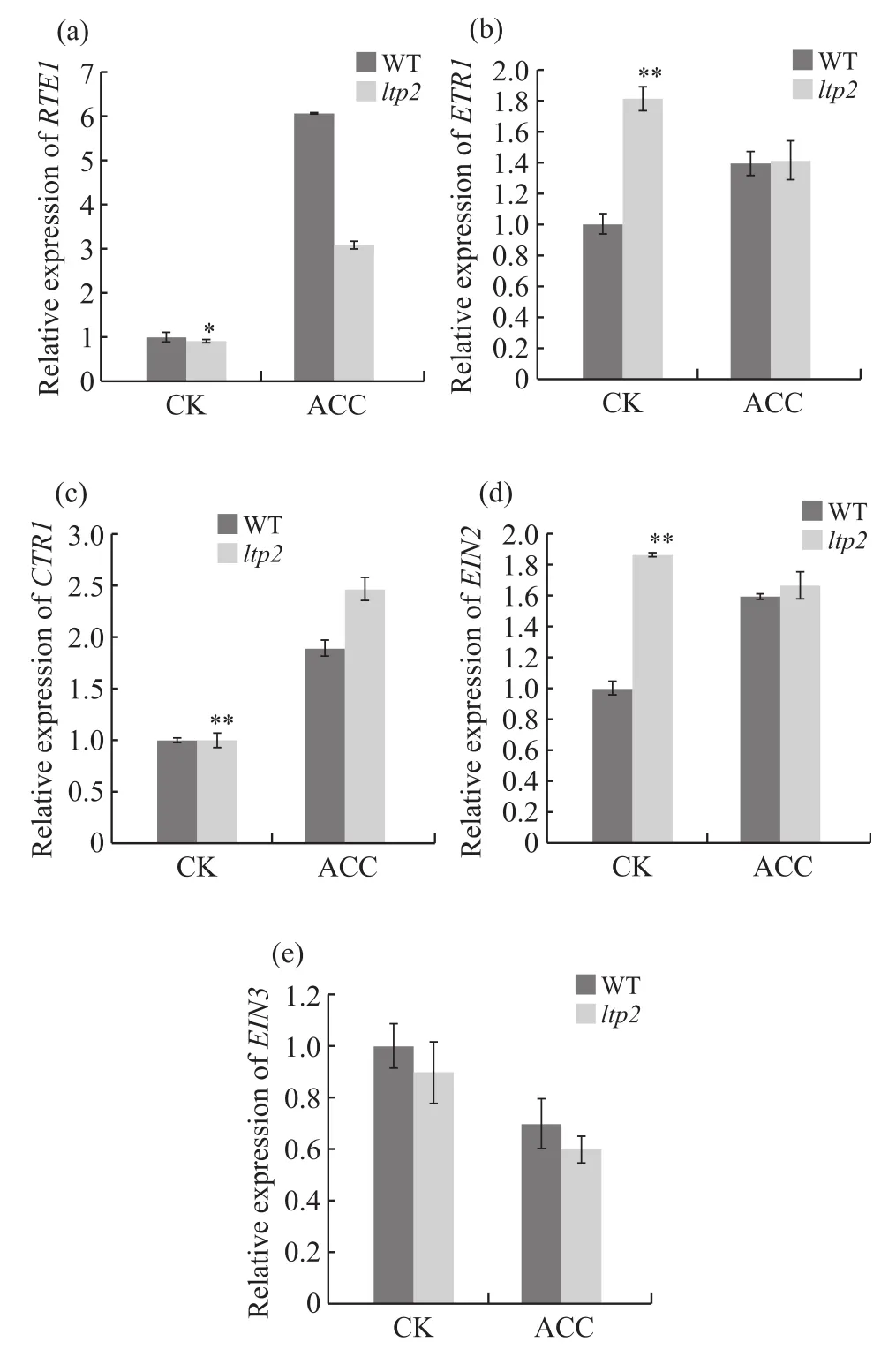
图4 乙烯信号通路基因相对表达量Fig. 4 The relative expression of ethylene signaling pathway genes
2.4 ltp2突变体中乙烯响应因子的表达量降低
乙烯响应因子ERF位于乙烯信号通路的下游,受乙烯信号调控并调控下游乙烯效应基因的表达,最终影响表型。ACC处理组中,野生型ERF1、ERF2和ERF9基因表达量比突变体分别高20.9%(图5a)、100.0%(图5b)、38.7%(图5c)。对照组中,野生型与突变体中的ERF1、ERF2和ERF9表达量无明显差异。从两者在处理组中的表达量差异可以看出,在ACC处理后突变体中以上3种ERF的表达量均低于野生型,表明突变体中乙烯信号要弱于野生型。
2.5 ltp2突变体的蜡质合成酶基因表达产生差别
乙烯参与调控植物表皮的蜡质合成[23],LTP2也参与蜡质层的形成[10]。对相关蜡质合成酶基因的表达量检测发现:野生型的蜡质合成酶基因CER3[26]的表达量在ACC处理后略有降低,而突变体中则升高了56.5%(图6a);野生型与突变体的CER6[27]的表达量在处理组与对照组中都呈下降趋势,但突变体中的表达量一直高于野生型(图6b)。突变体CER3的表达量在两组中的差别与野生型不同,与乙烯处理下CER3表达量的降低趋势相反[28-29],推测蜡质的合成受乙烯与LTP2的共同调控。
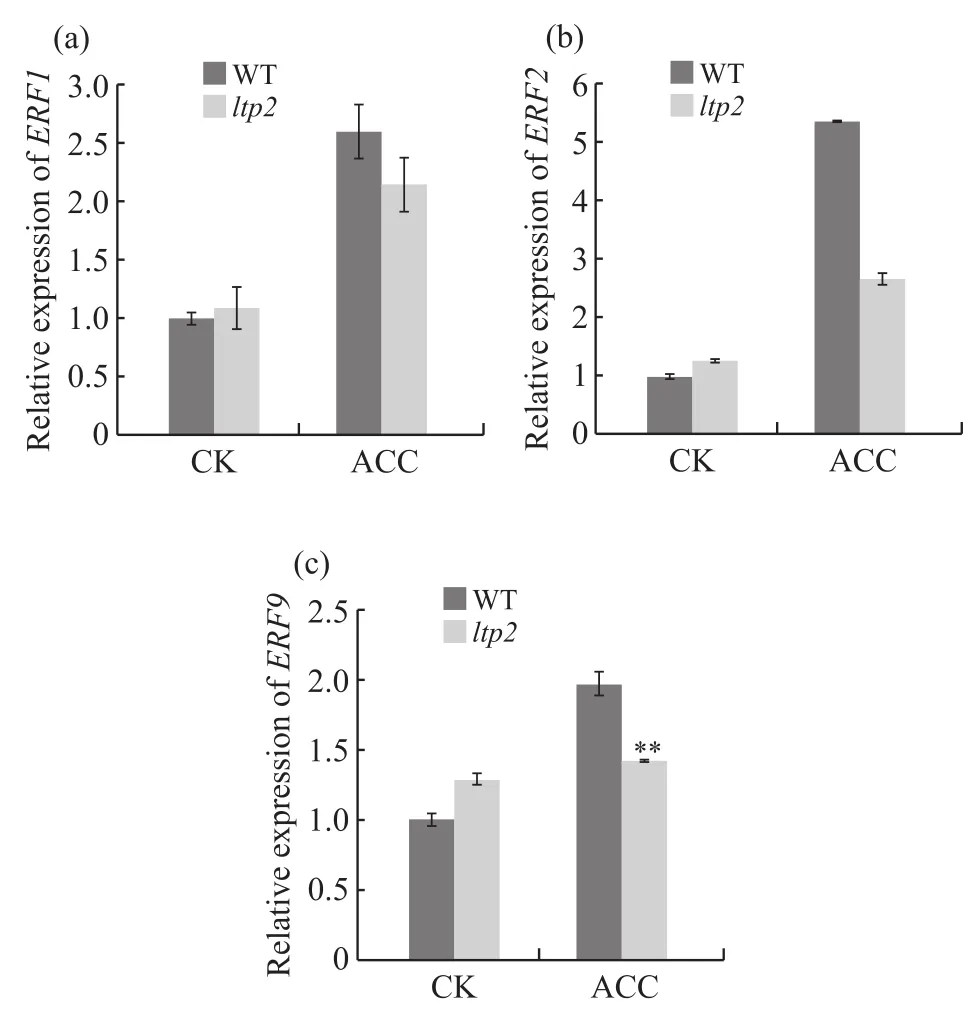
图5 乙烯响应因子基因相对表达量Fig. 5 The relative expression of ethylene response factor genes

图6 蜡质合成酶基因相对表达量Fig. 6 The relative expression of the waxy synthetase gene
3 讨论
使用适当浓度的外源乙烯或其前体处理拟南芥能出现典型的“三重反应”特征表型。利用这点,可以判断研究对象是否参与调控乙烯信号转导、调控属性与位点,还能研究乙烯与各类生理表型的关系。拟南芥中XBAT31基因的RNAi株系在乙烯处理下产生乙烯过敏感表型,而添加乙烯受体抑制剂AgNO3后,过敏感表型消失,表明XBAT31负调控乙烯信号转导且调控处于乙烯受体上游[30]。赵宇航等[31]通过对大量烟草乙烯不敏感突变体株系的低温处理,发现植物的乙烯不敏感性与低温抗性的提高有着密切的联系,其中的调控机制还需更深一步的研究。在本试验中,突变体中的LTP2表达量大幅下调,在ACC处理下表现为乙烯弱敏感,突变体整体下胚轴和根长略长于野生型;对照组中野生型与突变体植株长度无差异。试验结果表现出LTP2对乙烯信号转导的正调控,LTP2表达量的下调影响了乙烯信号的传递。
脂质转运蛋白功能广泛,拟南芥AtLTPd1的突变体atltpd1在接种丁香假单胞菌后,在其远叶中未能检测到相关病程的基因表达,证明AtLTPd1参与植物体内相关信号的转导[32]。在蜡质合成方面,有报导显示甘蓝型油菜在ACC处理后会导致蜡质中烷类含量与蜡质总量减少[29],其蜡质表面密度更高,覆盖面更光滑[33]。CER3是蜡质合成途径中一个重要的烷类合成酶,在某些LTP完全敲除突变体中烷类的含量减少[28,34]。本试验中野生型的CER3和CER6在ACC处理后表达量均呈下降趋势,符合其在乙烯处理后的基因表达改变趋势,突变体的表达量差异证明LTP2在乙烯诱导蜡质合成中有一定的作用。以上试验结果表明LTP2参与乙烯信号转导并影响下游响应因子的表达,其具体机制有待进一步深入研究。本文结果为深入研究脂质转运蛋白功能及其与植物激素的互作提供了新的参考依据。
