磺胺类抗生素对玉米幼苗生长、生理及代谢组的影响
2021-03-05贲蓓倍刘海学李娜杨仁杰
贲蓓倍 刘海学 李娜 杨仁杰




摘要:【目的】研究典型磺胺類抗生素(Sulfonamides,SAs)对玉米幼苗生长、生理及代谢组的影响,为进一步探究玉米对SAs的响应机制提供理论依据。【方法】以玉米品种郑单958和掖单13为试验材料,以不添加SAs为对照(0 mg/L),以终浓度为10、20、30、40和50 mg/L的磺胺甲基嘧啶(SM1)和磺胺二甲基嘧啶(SM2)为处理,测定分析对照及不同处理玉米幼苗的生长和生理指标,并通过气相色谱—质谱联用仪(GC-MS)对玉米幼苗代谢物的含量及种类进行检测。【结果】2种SAs处理后,玉米幼苗的根长、株高、鲜重、干重和叶绿素含量总体上表现出低促高抑的变化趋势。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性被激活,总体变化趋势为先上升后下降。经GC-MS检测出代谢物共70种,主要为有机酸、氨基酸、糖类及其衍生物和其他代谢物,将筛选到的12种差异代谢物富集到代谢通路上,结果映射得出7条显著的代谢路径:氨酰基-tRNA的生物合成,氰氨基酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,缬氨酸、亮氨酸和异亮氨酸的生物合成,乙醛酸和二羧酸的代谢,甘氨酸、丝氨酸和苏氨酸代谢,异喹啉生物碱的生物合成。【结论】SAs对玉米幼苗生长和生理指标的影响呈低促高抑的效果。高浓度SAs使玉米幼苗生长受抑,光合色素含量降低,遭受氧化损伤,具有不可忽视的生态毒性。氨基酸类代谢物为玉米幼苗响应SAs的关键代谢物。郑单958对SAs的耐性比掖单13强。
关键词: 磺胺类抗生素;玉米;生长;生理;代谢组
中图分类号: S513 文献标志码: A 文章编号:2095-1191(2021)11-2977-09
Effects of sulfonamides on the growth,physiology and metabolome of maize seedlings
BEN Bei-bei1, LIU Hai-xue1*, LI Na2, YANG Ren-jie1
(1Tianjin Agricultural University, Tianjin 300384, China; 2Fushun Forestry Development Service
Center, Fushun, Liaoning 113000, China)
Abstract:【Objective】In this study, we explored the effects of typical sulfonamide antibiotics(Sulfonamides, SAs) on the growth, physiology and metabolome of maize seedlings, in order to provide a theoretical basis for further explo-ring the response mechanism of maize to SAs. 【Method】 Two kinds of maize varieties (Zhengdan 958 and Yedan 13) were selected as the research materials with no added SAs as the control (0 mg/L), or with final concentrations of 10,20, 30, 40 and 50 mg/L sulfamerazine(SM1) or sulfamethazine (SM2) as the treatments. The growth and physiological indicators of maize seedlings were determined and the types and contents of metabolites of maize seedlings were detected by gas chromatography-mass spectrometry(GC-MS). 【Result】After SAs treatment,the root length,plant height,fresh weight dry weight and chlorophyll of maize seedlings showed a trend that low concentration promoted but high concentration inhibited growth. The activities of superoxide dismutase(SOD), peroxidase(POD) and catalase(CAT) enzymes were activated,and overall,the trend of change was firstly rising and then falling. A total of 70 metabolites were detected by GC-MS,mainly organic acids,amino acids,sugars and their derivatives and other metabolites.12 different metabolites were obtained,which were enriched on the metabolic pathways,and 7 significant metabolic pathways were mapped out:aminoacyl-tRNA biosynthesis;cyanoamino acid metabolism;alanine,aspartate and glutamate metabolism;valine,leucine and isoleucine biosynthesis;glyoxylate and dicarboxylate metabolism;glycine,serine and threonine metabolism;isoquinoline alkaloid biosynthesis. 【Conclusion】For maize seedlings treated with SAs,they show the effect of being promoted at low concentrations but inhibited at high concentrations. The high concentration of SAs inhibits the growth of maize seedlings,reduces the photosynthetic pigment content,suffers oxidative damage,and has a non-negligible ecotoxicity. Amino acid metabolites are the key metabolites of maize seedlings in response to SAs stress. The resistance of Zhengdan 958 against SAs is stronger than Yedan 13.
Key words: sulfonamids; maize; growth; physiology; metabolome
Foundation item: National Natural Science Foundation of China(41771357)
0 引言
【研究意义】近年来,抗生素污染逐渐受到关注。其中,磺胺类抗生素(Sulfonamids,SAs)是一类人工合成的有机物质,因其具有成本低、抗菌谱性广和稳定性强等优点而被广泛应用(陈姗等,2019)。SAs在使用后仅有小部分参与生物体代谢,大部分会以原药形式直接排放至多个环境介质,具有潜在的生态风险(Kumar et al.,2005)。玉米是我国典型粮食作物之一,具有极强的环境适应能力(赵久然等,2018)。在当前环境污染不断加剧的背景下,研究抗生素污染对玉米生长、生理及代谢变化的影响,对探究玉米响应逆境的机理具有重要意义,同时也可为抗逆玉米品种的选育提供新思路。【前人研究进展】抗生素长期暴露会改变土壤微生物群落的功能和结构多样性(Gutiérrez et al.,2010),诱发抗性菌及抗性基因的传播(张兰河等,2016;Grenni et al.,2019),进而产生新型污染。同时,抗生素能被植物吸收、转运并积累,影响植物的光合作用和正常生长。有研究显示,SAs可改变植物的根系地向性,对根系生长产生不利影响,影响水分吸收,具有很强的损害植物性能和生物量毒害作用(Michelini et al.,2012)。四环素类抗生素会降低淡水绿藻的细胞膜透性,造成严重的毒害作用进而抑制其生长(徐冬梅等,2013)。高浓度的土霉素污染对水稻苗期的生长及叶绿素含量具有显著抑制作用(徐秋桐等,2016)。随着SAs浓度的提高,苦草叶绿体的膜系统受到损伤,结构变形,严重的会造成叶绿体解体,进而影响光合作用(赵秀侠等,2017)。李亚宁等(2017)发现SAs在低浓度、高浓度染毒处理下能显著抑制油菜叶片叶绿素的合成。司雄元等(2018)研究表明,SAs在单一胁迫、复合胁迫下,随着处理浓度的升高,对小白菜的生长及叶绿素产生抑制作用,毒性作用表现为磺胺二甲基嘧啶强于磺胺。抗氧化酶是植物体内负责清除由环境影响产生的自由基的重要保护酶,其中超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是最主要的3种酶。肖明月等(2014)以小白菜为研究对象,对其进行6种常见抗生素处理,结果表明,6种抗生素均能显著诱导小白菜可溶性蛋白积累,对其抗氧化酶系统平衡造成破坏。严青云等(2016)发现环丙沙星对不同品种菜心的氧化损伤程度不同,高积累菜心品种对可溶性蛋白、POD和SOD活性的诱导作用强于低积累菜心品种,因此高积累菜心品种受到的氧化损伤较低积累菜心品种小。还有学者发现,随着抗生素浓度增加,小麦幼苗中的SOD活性显著增加,而CAT活性显著降低(Riaza et al.,2017)。经四环素与金霉素、四环素与磺胺嘧啶复合胁迫后,小白菜的SOD、POD和CAT活性均受到抑制,且受到的抑制作用强于单一抗生素处理(宫晓双等,2019)。【本研究切入点】代谢物是生物系统响应环境的体现者,通过确定和量化代谢物并对其进行信息提取与分析,以达到解释生化途径和响应机理的目的(Fiehn,2002),因此代谢组学是一个用于分析作物响应逆境的高效工具。但目前抗生素对植物影响的研究多集中在生长及生理影响方面(王朋等,2011;Ahmed et al.,2015),鲜有对代谢组影响的报道。【拟解决的关键问题】以2个玉米品种为研究材料,对其进行2种SAs处理,测定并分析生长、生理及代谢组学相关指标,研究典型SAs对玉米幼苗生长的影响,以期为进一步探究玉米对SAs的响应机制提供理论依据。
1 材料与方法
1. 1 试验材料
玉米品种为郑单958和掖单13;SAs为磺胺甲基嘧啶(Sulfamerazine,SM1)和磺胺二甲基嘧啶(Sulfamethazine,SM2),分别购于Aladdin和Macklin公司,纯度均为99%。
1. 2 试验方法
玉米幼苗培养采用水培法。玉米种子经75%酒精消毒15 min,经蒸馏水清洗5遍后置于培养盒中,于27 ℃黑暗催芽48 h,培养介质为去离子水。待种子露白后均匀选取15粒玉米种子置于27 ℃恒温培养箱中进行培养,光照12 h/黑暗12 h,光照强度4000 lx,湿度为60%,培养介质为Hoagland’s营养液。待培养至3叶1心期时,将培养介质换成SAs终浓度为10、20、30、40和50 mg/L的营养液,每处理重复3次,处理7 d。以不添加SAs的Hoagland’s营养液培养为对照(0 mg/L)。培养结束后收集玉米幼苗,洗净、吸干水分后进行生长、生理指标及代谢物测定。
1. 3 生长及生理指标测定
1. 3. 1 生长指标测定 在各处理中选取长势均匀的玉米幼苗6株,测定株高、根长和鲜重,经105 ℃杀青15 min,75 ℃烘至恒重,测定干重。
1. 3. 2 生理指标测定 生理指标参照苍晶和赵会杰(2013)的方法进行测定。叶绿素含量采用分光光度法测定,SOD活性采用氮蓝四唑光还原法测定,POD活性采用愈创木酚法测定,CAT活性采用分光光度法测定。
1. 4 代谢物提取及GC-MS条件
1. 4. 1 代谢物提取 将收集到的玉米幼苗全株经液氮研磨,混匀后称取150.0 mg于1 mL异丙醇-乙腈-水提取液(1∶1∶1,V/V/V)中4 ℃超声提取30 min,离心1 min(4 ℃,10000 r/min),吸取150 μL上清液氮吹后进行衍生(第一步:加入10 μL鹽酸甲氧胺吡啶,30 ℃衍生90 min;第二步:加入30 μL MSTFA+1% TMCS,37 ℃衍生30 min),最后加入1 mL(含20 μg十一烷内标物)正庚烷溶液,经0.22 μm滤膜过滤,滤液即为待测液。重复3次。
1. 4. 2 GC-MS条件 实验仪器为7890 A-5975 C型气相色谱—质谱联用仪(Aglient公司)。色谱条件:色谱柱为Aglient 19091S-433HP-5MS(30 m×250 μm×0.25 μm),载气为氦气(纯度99.999%),不分流进样。升温程序为常温升至60 ℃后维持1 min,以10 ℃/min升至270 ℃后运行35 min。质谱条件:电子碰撞内能70 eV,离子源温度230 ℃,四级杆温度150 ℃,溶剂延迟6 min,扫描速度2 scan/s,范围50~500 m/z,全扫描模式。
1. 5 统计分析
利用Excel 2019进行数据整理及计算,运用SPSS 26.0进行数据的差异性分析,Origin 2019制图。通过SIMCA-P 13.1对代谢组数据进行主成分分析(PCA)及偏最小二乘法判别分析(PLS-DA),利用Metabo Analyst平台进行代谢途径分析。
2 结果与分析
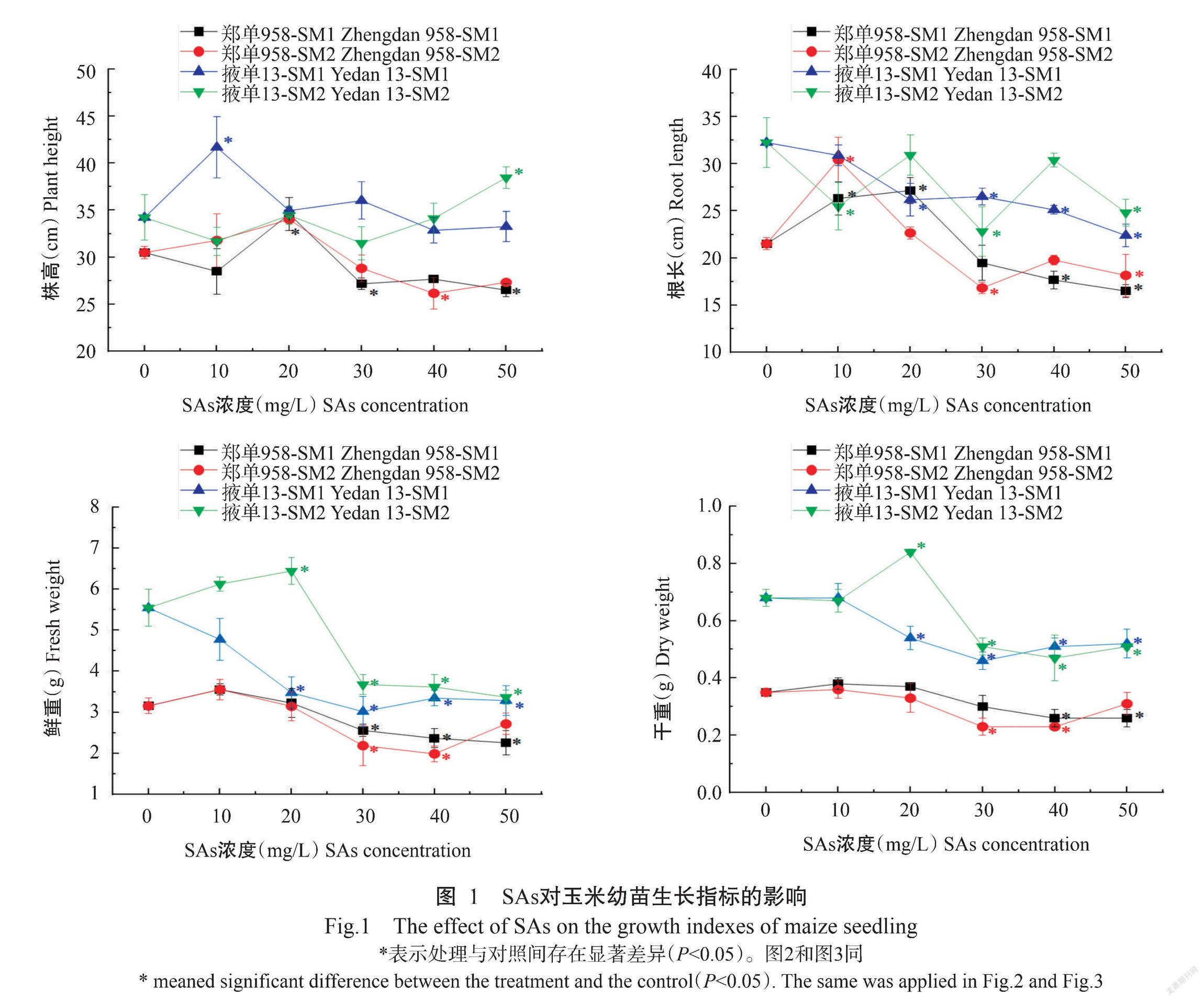
2. 1 SAs对玉米幼苗生长指标的影响
由图1可看出,SAs处理对玉米的生长指标均产生了一定程度的影响。在对株高的影响上,随着SM1、SM2浓度的升高,与对照相比,郑单958总体上表现出先被促进后被抑制的变化趋势, SM1和SM2的临界浓度[即指标受到显著(P<0.05,下同)抑制的最小处理浓度,下同]分别为30和40 mg/L;掖单13株高的变化趋势与郑单958相似,但在SAs处理范围内并未受到显著抑制(P>0.05,下同)。在对根长的影响上,随着SM1和SM2浓度的升高,与对照相比,郑单958表现出先被促进后被抑制的变化趋势,SM1和SM2的临界浓度分别为40和30 mg/L,掖单13则在SM1和SM2处理浓度范围内均被抑制,临界浓度分别为20和10 mg/L。在对鲜重和干重的影响上,随着SM1和SM2浓度的升高,郑单958鲜重和干重的变化趋势一致,与对照相比,均表现为先被促进后被抑制。郑单958在SM1处理下鲜重和干重的临界浓度分别为30和40 mg/L,在SM2处理下鲜重和干重的临界浓度均为30 mg/L。掖单13在SM1处理浓度范围内鲜重和干重均被抑制,其中在20~50 mg/L范围内抑制作用显著,鲜重和干重的临界浓度均为20 mg/L。在SM2处理下,掖单13的鲜重和干重表现为10 mg/L时被促进,20 mg/L时被显著促进,而30~50 mg/L时被显著抑制,临界浓度均为30 mg/L。
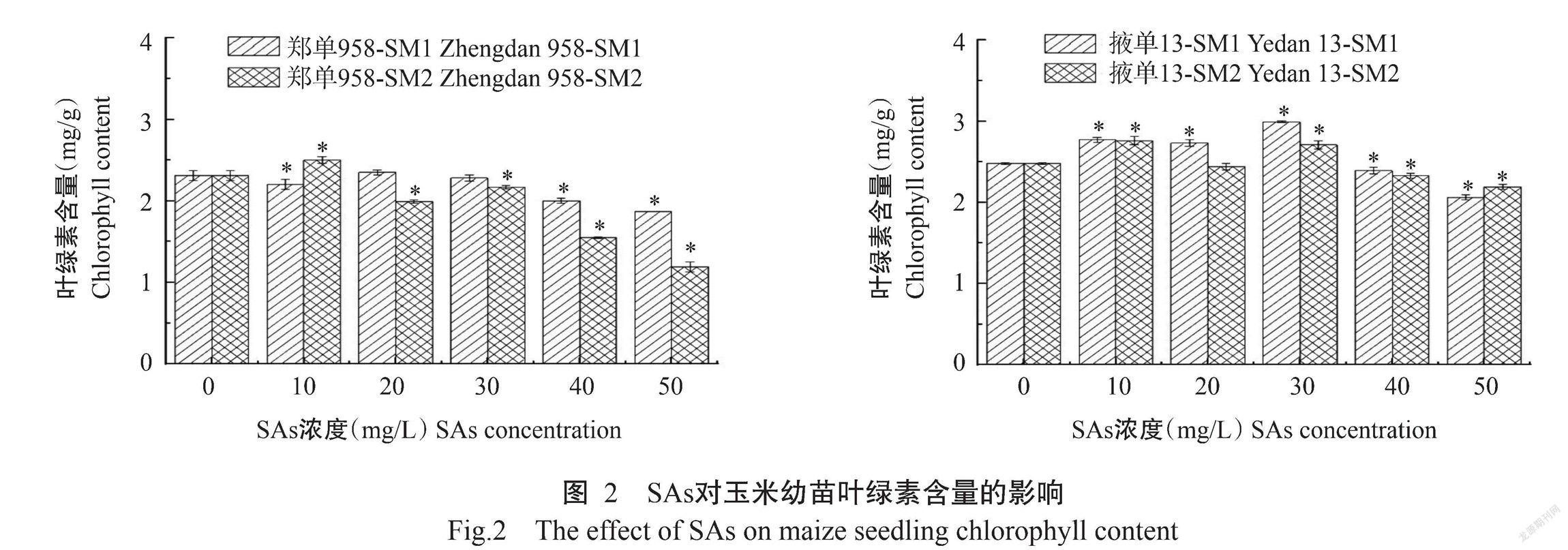
2. 2 SAs对玉米生理指标的影响
2. 2. 1 SAs对玉米幼苗叶绿素含量的影响 从图2可看出,郑单958经SAs处理后,与对照相比,在SM1处理下,叶绿素含量在10 mg/L时显著降低,在20~30 mg/L时有所提高但与对照差异不显著,在40~50 mg/L时显著降低;在SM2处理下,叶绿素含量整体呈先升高后降低的变化趋势,40~50 mg/L时表现出显著抑制作用。其中SM2处理下的叶绿素含量变幅(-48%~8%)大于SM1处理(-19%~2%),SM1处理的临界浓度为10 mg/L,SM2处理的临界浓度为20 mg/L。掖单13经SAs处理后,从变幅看,SM1处理下的叶绿素含量变幅(-17%~21%)大于SM2处理(-12%~12%),SM1和SM2处理的临界浓度均为40 mg/L。
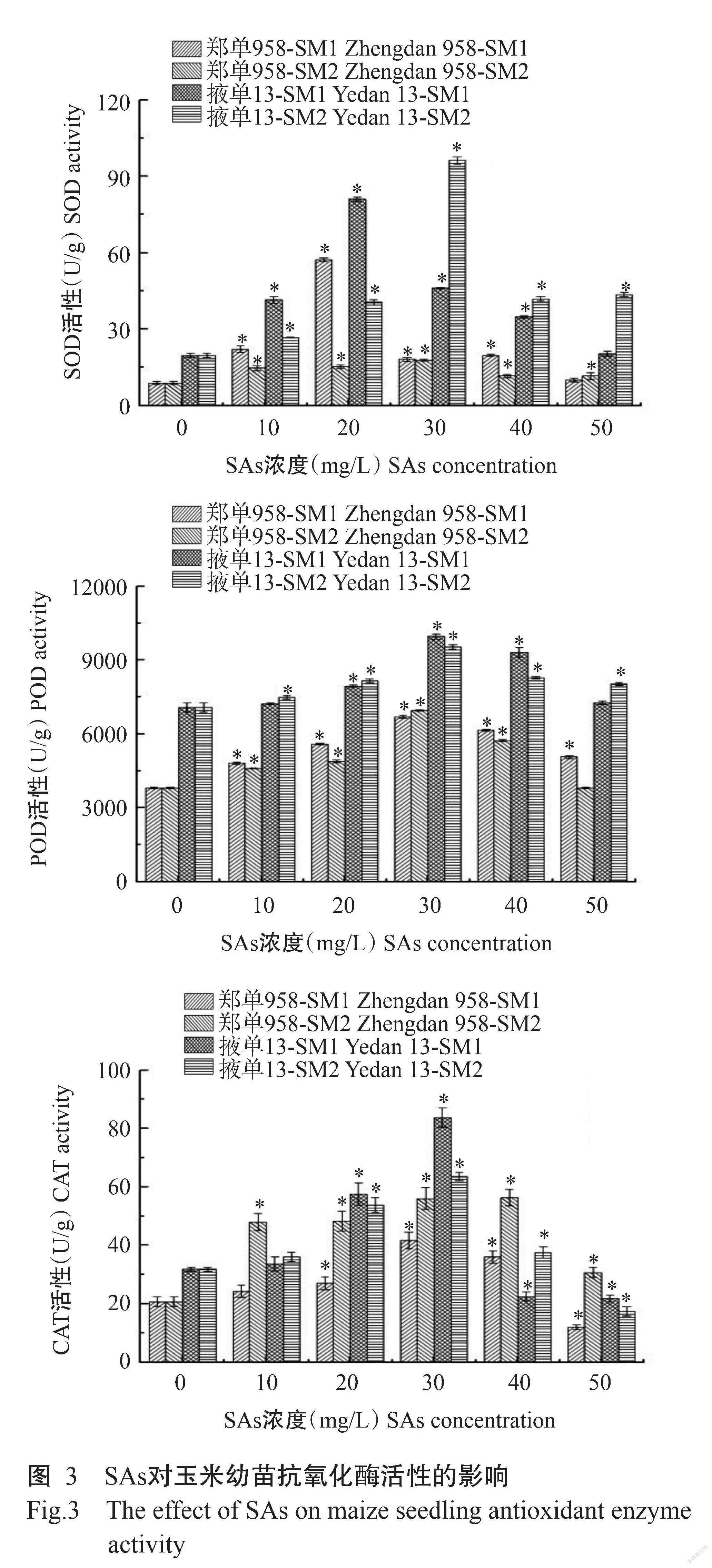
2. 2. 2 SAs对玉米幼苗抗氧化酶活性的影响 抗氧化酶是植物体内重要的保护酶体系,其变化反映植物抗氧化损伤的能力。从图3可看出,郑单958和掖单13经SAs处理后,SOD和POD活性均高于对照,整体表现为先上升后下降的变化趋势,表明酶的保护作用有限,在达到最大限度后酶活性会逐渐降低。其中,在SM1处理下,郑单958和掖单13的SOD活性均在浓度为20 mg/L时达最高值;在SM2处理下,2个品种的SOD活性均在浓度为30 mg/L时达最高值,SM1的最大促进作用浓度小于SM2,表明SM1对SOD活性的影响比SM2大。POD活性在SM1和SM2处理下均在浓度为30 mg/L时达最高值。在SM1处理下,郑单958和掖单13的CAT活性均表现出低浓度被促进高浓度被显著抑制的趋势,临界浓度分别为50和40 mg/L。在SM2处理下,郑单958的CAT活性均高于对照,浓度为40 mg/L时CAT活性最高;掖单13的CAT活性表现出低浓度被促进高浓度被显著抑制的趋势,临界浓度为50 mg/L。总体而言,SM1处理对玉米抗氧化酶活性的影响相对更大,郑单958在同等条件下其抗氧化酶活性受SAs的影响小于掖单13。
2. 3 SAs对玉米幼苗代谢物的影响
2. 3. 1 代谢物种类鉴定 采用GC-MS检测玉米幼苗代谢物,结果共鉴定出70种代谢物,包括有机酸27种、糖类及其衍生物18种、氨基酸14种和其他有机物11种,分别占总代谢物总数的38.6%、25.7%、20.0%和15.7%。
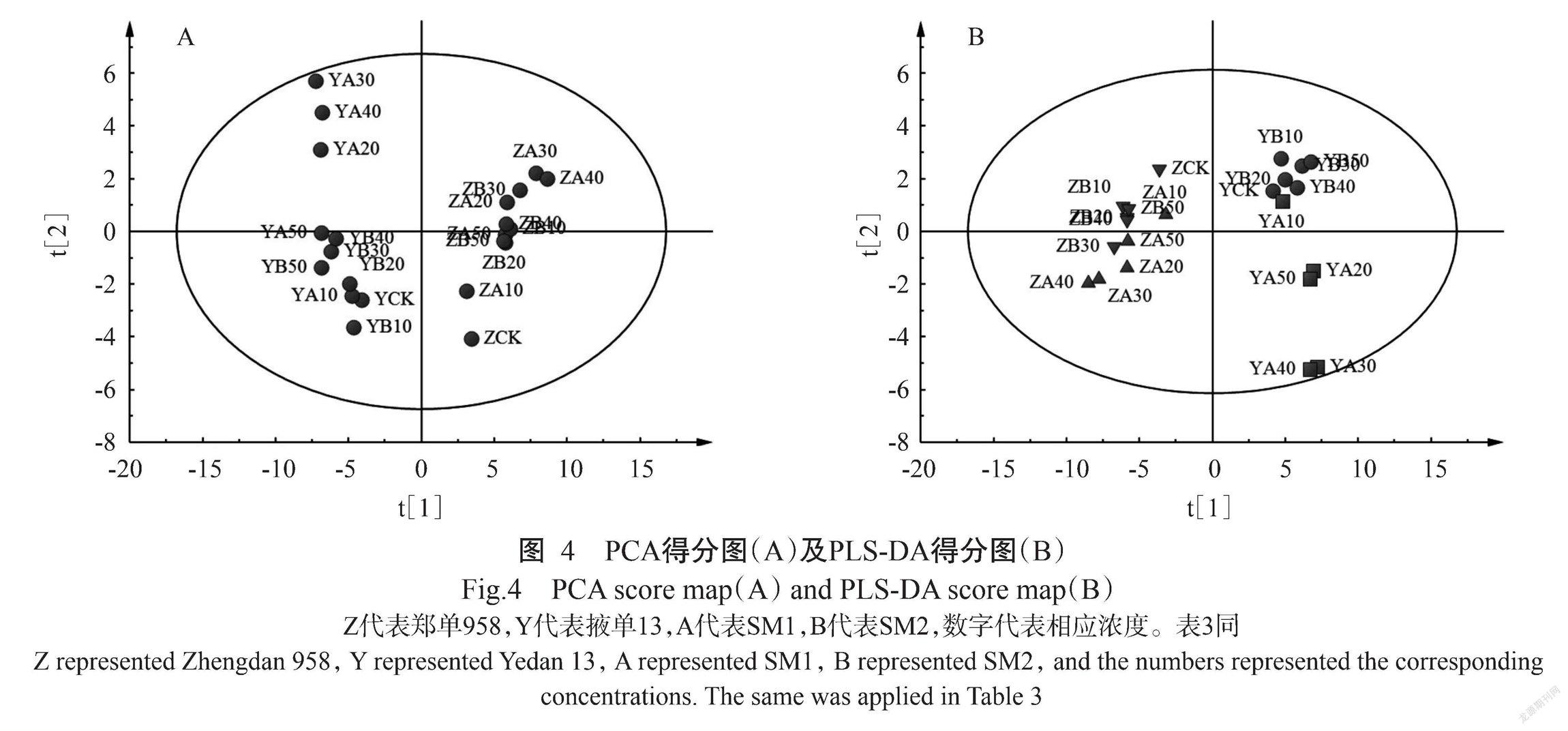
2. 3. 2 代谢物的多元统计分析 将3组生物学重复代谢物数据取平均值后进行多元统计分析,经PCA分析得到得分图(图4-A),结果表明,郑单958和掖单13的样品能较好区分开来,说明SAs对玉米的影响具有显著的品种差异性;掖单13样品分散性较郑单958样品大,表明掖单13受SAs的影响大于郑单958。就单一品种而言,不同SAs处理的样品分离效果不理想。因此,需进一步进行PLS-DA分析。
通过PLS-DA得分图(图4-B)可见,SM1、SM2处理总体上能分开,掖单13样品的分离效果优于郑单958样品。就SM1处理而言,2个玉米品种均表现出20和50 mg/L间代谢物差异较小,30和40 mg/L间代谢物差异较小。就SM2处理而言,掖单13各样品间差异不明显,郑单958样品中,30 mg/L与其他样品差异明显。由此可见,在同一浓度范围内,SM1對玉米的影响大于SM2。经过200次置换模型检验,Q2值与R2Y值均低于相应的原始值,因此判断模型未存在过拟合现象,可依据其给出的变量投射重要度(Varia-ble importance for the projection,VIP)筛选差异代谢物。
2. 3. 3 差异代谢物分析 将PLS-DA分析中得到的VIP值结合单因素方差分析P值筛选差异代谢物(VIP>1,P<0.05),结果(表2)显示,最终筛选出12种差异代谢物,为8种氨基酸和4种有机酸。由此可见,SAs对玉米幼苗中氨基酸类和有机酸类代谢物含量的影响较大。
对差异代谢物进行含量的变化倍数分析,log2FC>0表示上调,log2FC<0表示下调,|log2FC|>1表示显著上调或显著下调。由表3可知,就掖单13样品中的氨基酸含量而言,SM1、SM2在各浓度处理均表现上调趋势,除YA10、YB10、YB20和YB50外,其他浓度均表现出显著上调作用。就郑单958样品中的氨基酸含量而言,除ZB20外,SM1、SM2在各浓度处理均表现上调趋势,但均未达显著水平。表明SAs对掖单13样品影响较显著。在掖单13样品中,SM1和SM2处理在10~40 mg/L范围内,氨基酸含量随着浓度的上升而上升,在40 mg/L表现出最大的积累作用,而在50 mg/L时积累减少。就2个品种玉米样品中的有机酸含量而言,SM1、SM2对其含量的影响或上升或下降,但均未达显著水平。
2. 4 SAs对玉米幼苗代谢途径的影响
将差异代谢物导入Metabo Analyst平台进行分析,依据P<0.05或Impact>0.2的判断标准,最终筛选出具有显著意义的7条代谢通路(表4)。表4中标注了代谢路径参与的总代谢物个数(Total)及匹配到的玉米幼苗的代谢物个数(Hits)。Impact值表示经拓扑分析得出的代谢路径影响值。变化显著的代谢通路中超过半数通路与氨基酸的代谢和合成有关,说明SAs对玉米幼苗中氨基酸的代谢途径具有明显影响。
3 讨论
3. 1 SAs对玉米幼苗生长和生理指标的影响
有研究表明,经抗生素处理后,水芹的生长在低浓度被促进,而在高浓度时呈现出显著的抑制效果。对SAs处理下的玉米幼苗生长进行研究,观察到随着SAs浓度的升高,各生长指标表现出低促高抑的效果,与鲍陈燕等(2016)关于抗生素胁迫对水芹生长影响的研究结果一致。这种低促高抑现象的出现,有可能是当SAs处于较低浓度时,抗逆活动的激发使代谢活动同步增强,进而促进了植物的生长(周文礼等,2009),这一猜想也在本研究的代谢组学分析中得到印证。而SAs处于较高浓度时,植物对SAs的吸收和转运活动增强,内环境的稳定被破坏(迟荪琳等,2018),最终导致生长被抑制。
光合作用是植物最重要的环节之一,其正常稳定运作是能量来源和营养积累的基础,同时也是响应逆境的重要体现(Abbasi et al.,2015)。叶绿素是植物光合作用的重要组成部分,其含量的变化能客观反映生物体受到的影响。本研究结果表明,高浓度SAs能抑制玉米幼苗叶绿素的合成。有研究表明,高浓度的抗生素能抑制光合色素和光合作用相关蛋白的合成,使叶绿体产生结构性损伤,最大光合速率下降,呼吸作用升高,最终影响光合作用(Kasai,2004;Vannini et al.,2011)。
抗氧化酶系统是植物体内清除活性氧自由基的重要保护酶系统,植物在受到逆境胁迫时,体内的抗氧化酶体系会被激活(魏瑞成等,2011;李亚宁等,2017)。在SAs处理下,SOD、POD和CAT活性均被激活,表明玉米幼苗受到了氧化损伤。本研究中,抗氧化酶活性的变化趋势为先上升后下降,表明植物抗氧化体系抵御逆境的能力有一定限度。当SAs濃度超过植物的耐受极限,SAs的富集作用使氧化损伤作用加深,或与关键酶活位点相结合形成络合物,最终使酶活性降低(宫晓双等,2019)。
3. 2 SAs对玉米幼苗代谢物及代谢途径的影响
经代谢组学分析玉米幼苗中差异代谢物含量变化,发现氨基酸含量变化显著,但有机酸含量变化较小。差异氨基酸含量随SAs浓度的升高呈先上升后下降的变化趋势,说明随着SAs胁迫的加剧,玉米幼苗的抗逆活动增强,但超过一定的耐受限度后,抗逆活动会减弱,进而受到损害。
氨基酸是生物体受到环境压力的响应者,其变化对研究生物体抗逆性有重要的指导作用。燕辉等(2012)研究发现,在盐胁迫条件下,花棒叶片的丙氨酸、脯氨酸等主要游离氨基酸不断合成与积累,参与渗透调节与离子运输等活动,以达到缓解逆境伤害的作用。本研究中,除丙氨酸外,SAs胁迫导致差异氨基酸含量总体上升,与燕辉等(2012)的研究结果相似。此外,4条与氨基酸相关的代谢途径变化显著,表示玉米幼苗在SAs胁迫时氨基酸活动较活跃,是响应SAs胁迫的重要调节物质。氨基酸是合成抗逆蛋白的重要原料。本研究中,氨酰基-tRNA的生物合成途径变化显著,表明蛋白质的合成也较活跃。在受到SAs胁迫时,玉米幼苗可合成相关的抗逆蛋白,减轻逆境损伤,维持细胞液和细胞结构的稳态,保证生物体的正常生长发育及代谢(王艳蓉等,2015)。
有机酸是参与植物抗逆的渗透性物质。本研究中,各SAs处理使有机酸含量或上升或下调,但影响均不显著,推测玉米幼苗在响应SAs胁迫时,有机酸参与抗逆但可能不是主要环节。戴凌燕等(2017)在甜高粱响应苏打盐碱胁迫的研究中发现氨基酸生物合成和二羧酸代谢途径达显著水平。有报道发现,棉花在遭遇盐胁迫时,乙醛酸和二羧酸代谢途径与抗逆相关性较大,主要通过平衡植物体内局部代谢紊乱,加快能量代谢以提高耐性(王德龙等,2010)。本研究与上述前人研究结果一致,表明乙醛酸和二羧酸的代谢路径在玉米幼苗响应逆境的调控中起积极作用。
异喹啉生物碱是一类具有清除自由基和抗脂质过氧化活性的物质(程轩轩等,2006)。耐盐番茄(张国儒等,2018)、耐冷黄芪(华梦艺等,2017)在逆境下均能使异喹啉生物碱生物合成的相关基因上调达显著水平,使植物体减轻或消除因胁迫而造成的伤害。本研究中,异喹啉生物碱的生物合成途径变化显著,推测其可能在玉米幼苗SAs胁迫响应中起重要作用。
4 结论
SAs对玉米幼苗生长和生理指标的影响呈低促高抑的效果。高浓度SAs使玉米幼苗生长受抑,光合色素含量降低,遭受氧化损伤,具有不可忽视的生态毒性。氨基酸类代谢物为玉米幼苗响应SAs的关键代谢物,可从氨基酸动态变化的角度进一步研究玉米响应抗生素的机理。总体来说,郑单958对SAs的耐性比掖单13强,可做为抗逆研究的供试材料。
参考文献:
鲍陈燕,顾国平,章明奎. 2016. 兽用抗生素胁迫对水芹生长及其抗生素积累的影响[J]. 土壤通报,47(1):164-172.[Bao C Y,Gu G P,Zhang M K. 2016. Effects of veterinary antibiotics stress on growth and antibiotics accumulation of Oenanthe javanica DC[J]. Chinese Journal of Soil Science,47(1):164-172.] doi:10.19336/j.cnki.trtb. 2016.01.026.
苍晶,赵会杰. 2013. 植物生理学实验教程[M]. 北京:高等教育出版社. [Cang J,Zhao H J. 2013. Experimental course of plant physiology[M]. Beijing:Higher Education Press.
陈姗,许凡,张玮,唐文乔,王丽卿. 2019. 磺胺类抗生素污染现状及其环境行为的研究进展[J]. 环境化学,38(7):1557-1569. [Chen S,Xu F,Zhang W,Tang W Q,Wang L Q. 2019. Research progress in pollution situation and environmental behavior of Sulfonamides[J]. Environmental Chemistry,38(7):1557-1569.] doi:10.7524/j.issn. 0254-6108.2018091901.
程轩轩,王冬梅,杨得坡. 2006. 异喹啉类生物碱的生物活性和构效关系研究进展[J]. 中草药(12):1900-1904. [Cheng X X,Wang D M,Yang D P. 2006. Advances in studies on biological activity and structure-activity relationships of isoquinoline alkaloids[J]. Chinese Traditional and Herbal Drugs,(12):1900-1904.] doi:10.3321/j.issn: 0253-2670.2006.12.052.
迟荪琳,王卫中,徐卫红,李桃,李彦华,张春来. 2018. 四环素类抗生素对不同蔬菜生长的影响及其富集转运特征[J]. 环境科学,(2):935-943. [Chi S L,Wang W Z,Xu W H,Li T,Li Y H,Zhang C L. 2018. Effects of tetracycline antibiotics on growth and characteristics of enrichment and transformation in two vegetables[J]. Environmental Science,(2):935-943.] doi:10.13227/j.hjkx.201706250.
戴凌燕,杜吉到,张玉先,朱洪德,殷奎德. 2017. 甜高粱响应苏打盐碱胁迫的蛋白质组学分析[J]. 生态学杂志,36(6):1597-1605. [Dai L Y,Du J D,Zhang Y X,Zhu H D,Yin K D. 2017. Proteomics analysis of sweet sorghum in response to soda saline-alkali stress[J]. Chinese Journal of Ecology,36(6):1597-1605.] doi:10.13292/j.1000-4890. 201706.028.
宫晓双,安婧,张立娜,魏树和. 2019. 典型抗生素复合污染对小白菜生长发育的毒理效应[J]. 生態学杂志,38(2):541-547. [Gong X S,An J,Zhang L N,Wei S H. 2019. Toxicological effects of combined pollution of typical antibiotics on the development of Chinese white cabbage(Brassica rapa) seedlings[J]. Chinese Journal of Ecology,38(2) :541-547.] doi:10.13292/j.1000-4890.201902.004.
华梦艺,顾卓雅,殷甘强,王伟,南蓬. 2017. 低温胁迫下蒙古黄芪差异表达基因分析[J]. 植物生理学报,53(4):581-590. [Hua M Y,Gu Z Y,Yin G Q,Wang W,Nan P. 2017. Analysis of differentially expressed genes profile under chilling stress in Astragalus membranaceus Bge. var. mongolicus(Bge.) Hsiao[J]. Plant Physiology Journal,53(4):581-590.] doi: 10.13592/j.cnki.ppj.2016.0537.
李亚宁,张丽红,殷艳艳,张晨,刘刚,李凤祥. 2017. 典型磺胺类抗生素对油菜叶片叶绿素、可溶性蛋白及抗氧化酶的影响[J]. 环境污染与防治,39(11):1209-1212. [Li Y N,Zhang L H,Yin Y Y,Zhang C,Liu G,Li F X. 2017. Effects of typical sulfonamides on the chlorophyll,soluble protein contents and peroxidases activity of Brassica campestris[J]. Environmental Pollution & Control,39(11):1209-1212.] doi:10.15985/j.cnki.1001-3865.2017. 11.010.
司雄元,熊科胜,徐慧敏,檀华蓉,司友斌. 2018. 磺胺-磺胺二甲基嘧啶污染对小白菜生长和生理指标的影响及相关性分析[J]. 生态与农村环境学报,34(6):554-562. [Si X Y,Xiong K S,Xu H M,Tan H R,Si Y B. 2018. Effects of sulfonamide-sulfamethazine on growth and physio-logical indexes of Brassica campestris and its corre- lation analysis[J]. Journal of Ecology and Rural Environment,34(6):554-562.] doi:10.11934/j.issn.1673-4831. 2018.06.011.
王德龙,叶武威,王俊娟,宋丽艳,樊伟丽,崔宇鹏. 2010. 干旱胁迫下棉花SSH文库构建及其抗旱相关基因分析[J]. 作物学报,36(12):2035-2044. [Wang D L,Ye W W,Wang J J,Song L Y,Fan W L,Cui Y P. 2010. Construction of SSH library and its analyses of cotton drought associated genes under drought stress[J]. Acta Agronomica Sinica,36(12):2035-2044.] doi:10.3724/SP.J.1006.2010. 02035.
王朋,温蓓,张淑贞. 2011. 环丙沙星对玉米芽期抗氧化酶活性及自由基代谢的影响[J]. 环境化学,30(4):753-759. [Wang P,Wen B,Zhang S Z. 2011. Influence of ciprofloxacin(CIP) on antioxidative enzymes and free radical levels in maize(Zea mays L.) early stage development[J]. Environmental Chemistry,30(4):753-759.]
王艳蓉,张治国,吴金霞. 2015. LEA蛋白及其在植物抗逆改良中的应用[J]. 生物技术通报,(3):1-9. [Wang Y R,Zhang Z G,Wu J X. 2015. LEA protein and its application in improvement of stress tolerance in plants[J]. Biotechnology Bulletin,(3):1-9.] doi:10.13560/j.cnki.biotech. bull.1985.2015.04.001.
魏瑞成,裴燕,政晓丽,朱旭博,陈明,王冉. 2011. 金霉素和其代谢物胁迫对小白菜幼苗生长和抗氧化酶活性及累积效应的影响[J]. 草业学报,20(5):102-110. [Wei R C,Pei Y,Zheng X L,Zhu X B,Chen M,Wang R. 2011. Effects of chlortetracycline and 4-epi-chlortetracycline stress on seeding growth,antioxidant enzyme activities and accumulation in pakchoi[J]. Acta Prataculyurae Sinica,20(5):102-110.] doi:10.11686/cyxb20110514.
肖明月,安婧,紀占华,崔爽,李萍. 2014. 六种常见抗生素对小白菜种子萌发及生理特性的影响[J]. 生态学杂志,33(10):2775-2781. [Xiao M Y,An J,Ji Z H,Cui S,Li P. 2014. Toxic effects of six typical antibiotics on seed germination and physiological characteristics of Chinese cabbage[J]. Chinese Journal of Ecology,33(10):2775-2781.] doi: 10.13292/j.1000-4890.2014.0244.
徐冬梅,王艳花,饶桂维. 2013. 四环素类抗生素对淡水绿藻的毒性作用[J]. 环境科学,34(9):3386-3390. [Xu D M,Wang Y H,Rao G W. 2013. Cellular response of freshwater green algae to the toxicity of tetracycline antibiotics[J]. Environmental Science,34(9):3386-3390.]
徐秋桐,顾国平,章明奎. 2016. 土壤中兽用抗生素污染对水稻生长的影响[J]. 农业资源与环境学报,33(1):60-65. [Xu Q T,Gu G P,Zhang M K. 2016. Effects of soil ve-terinary antibiotics pollution on rice growth[J]. Journal of Agricultural Resources and Environment,33(1):60-65.] doi:10.13254/j.jare.2015.0133.
严青云,向垒,吴小莲,姜元能,李彦文. 2016. 环丙沙星高、低积累菜心(Brassica campestris L.)抗氧化性能的差异[J]. 环境科学学报,36(7):2694-2700. [Yan Q Y,Xiang L,Wu X L,Jiang Y N,Li Y W. 2016. Difference of anti-oxidation activities in Chinese flowering cabbages(Brassica campestris L.)with high and low accumulation of ciprofloxacin[J]. Acta Scientiae Circumstantiae,36(7) :2694-2700.] doi:10.13671/j.hjkxxb.2015.0763.
燕輝,彭晓邦,薛建杰. 2012. NaCl胁迫对花棒叶片光合特性及游离氨基酸代谢的影响[J]. 应用生态学报,23(7):1790-1796. [Yan H,Peng X B,Xue J J. 2012. Effects of NaCl stress on leaf photosynthesis characteristics and free amino acid metabolism of Heyedysarum scoparium[J]. Chinese Journal of Applied Ecology,23(7):1790-1796.] doi: 10.13287/j.1001-9332.2012.0234.
张国儒,庞胜群,郭晓珊,单淑玲. 2018. 加工番茄耐盐突变体耐盐相关基因的转录组分析[J]. 分子植物育种,16(18):5884-5896. [Zhang G R,Pang S Q,Guo X S,Shan S L. 2018. Transcriptome analysis of salt-tolerance-rela-ted genes in processing tomato salt-tolerant mutants[J]. Molecular Plant Breeding,16(18):5884-5896.] doi:10. 13271/j.mpb.016.005884.
张兰河,王佳佳,哈雪姣,邱孟超,高敏,仇天雷,王旭明. 2016. 北京地区菜田土壤抗生素抗性基因的分布特征[J]. 环境科学,37(11):4395-4401. [Zhang L H,Wang J J,Ha X J,Qiu M C,Gao M,Qiu T L,Wang X M. 2016. Distribution characteristics of antibiotic resistance genes in vegetable soils in Beijing[J]. Environmental Science,37(11):4395-4401.] doi:10.13227/j.hjkx.201605077.
赵久然,王帅,李明,吕慧颖,王道文,葛毅强,魏珣,杨维才. 2018. 玉米育种行业创新现状与发展趋势[J]. 植物遗传资源学报,19(3):435-446. [Zhao J R,Wang S,Li M,Lü H Y,Wang D W,Ge Y Q,Wei X,Yang W C. 2018. Current status and perspective of maize breeding[J]. Journal of Plant Genetic Resources,19(3):435-446.] doi:10. 13430/j.cnki.jpgr.2018.03.008.
赵秀侠,侯冠军,李静,卢文轩. 2017. 磺胺对苦草(Vallisneria natans(Lour.)Hara)生理生长及细胞超微结构的影响[J]. 安徽大学学报(自然科学版),41(2):92-99. [Zhao X X,Hou G J,Li J,Lu W X. 2017. Physiological respoints and ultrastructure changes of Vallisneria natans(Lour.)Hara under sulfonamides stress[J]. Journal of Anhui University(Natural Science Edition),41(2):92-99.] doi:10.3969/j.issn.1000-2162.2017.02.013.
周文礼,乔秀亭,肖慧,王悠,曲良,唐学玺,张培玉. 2009. 三种抗生素对几种海洋微藻叶绿素a含量影响的初步研究[J]. 海洋环境科学,28(3):268-271. [Zhou W L,Qiao X T,Xiao H,Wang Y,Qu L,Tang X X,Zhang P Y. 2009. Effects of antibiotics on chlorophyll-a level in marine microalgae[J]. Marine Environmental Science,28(3):268-271.] doi:10.3969/j.issn.1672-5174.2007.06.009.
Abbasi G H,Akhtar J,Anwar-ul-Haq M,Malik W,Ali S,Chen Z H,Zhang G P. 2015. Morpho-physiological and micrographic characterization of maize hybrids under NaCl and Cd stress[J]. Plant Growth Regulation,75(1):115-122. doi:10.1007/s10725-014-9936-6.
Ahmed M B M,Rajapaksha A U,Lim J E,Vu N T,Kim I S,Kang H M,Lee S S,Ok Y S. 2015. Distribution and accumulative pattern of tetracyclines and sulfonamides in edible vegetables of cucumber,tomato,and lettuce[J]. Journal of Agricultural and Food Chemistry,63(2):398-405. doi:10.1021/jf5034637.
Fiehn O. 2002. Metabolomics—The link between genotypes and phenotypes[J]. Plant Molecular Biology,48(1-2):155-171. doi:10.1023/A:1013713905833.
Grenni P,Patrolecco L,Rauseo J,Spataro F,Di Lenola M,Aimola G,Zacchini M,Pietrini F,Di Baccio D,Stanton I C,Gaze W H,Caracciolo A B. 2019. Sulfamethoxazole persistence in a river water ecosystem and its effects on the natural microbial community and Lemna minor plant[J]. Microchemical Journal,149:103999. doi:10.1016/j.microc.2019.103999.
Gutiérrez I R,Watanabe N,Harter T,Glaser B,Radke M. 2010. Effect of sulfonamide antibiotics on microbial diversity and activity in a Californian Mollic Haploxeralf[J]. Journal of Soils and Sediments,10(3):537-544. doi:0.1007/s11368-009-0168-8.
Kasai K. 2004. Guanosine tetra- and pentaphosphate synthase activity in chloroplasts of a higher plant:Association with 70S ribosomes and inhibition by tetracycline[J]. Nucleic Acids Research,32(19):5732-5741. doi:10.1093/nar/ gkh916.
Kumar K,Gupta S C,Baidoo S K,Chander Y,Rosen C J. 2005. Antibiotic uptake by plants from soil fertilized with animal manure[J]. Journal of Environmental Quality,34(6):2082-2085. doi:10.2134/jeq2005.0026.
Riaza L,Mahmooda T,Coyneb M S,Khalida A,Rashida A,Hayatc M T,Gulzard A,Amjade M. 2017. Physiological and antioxidant response of wheat(Triticum aestivum) seedlings to fluoroquinolone antibiotics[J]. Chemosphere,177:250-257. doi:10.1016/j.chemosphere.2017.03.033.
Michelini L,Reichel R,Werner W,Ghisi R,Thiele-Bruhn S. 2012. Sulfadiazine uptake and effects on Salix fragilis L. and Zea mays L. plants[J]. Water,Air,& Soil Pollution,223(8):5243-5257. doi:10.1007/s11270-012-1275-5.
Vannini C,Domingo G,Marsoni M,De Mattia F,Labra M,Castiglioni S,Bracale M. 2011. Effects of a complex mixture of therapeutic drugs on unicellular algae Pseudokirchneriella subcapitata[J]. Aquatic Toxicology(Amsterdam, Netherlands),101(2):459-465. doi:10.1016/j.aquatox.2010.10.011.
收稿日期:2021-02-06
基金項目:国家自然科学基金项目(41771357)
通讯作者:刘海学(1965-),http://orcid.org/0000-0001-7330-4660,博士,研究员,主要从事作物遗传育种研究工作,E-mail:hxliu 1965@sina.com
第一作者:贲蓓倍(1995-),http://orcid.org/0000-0003-3862-4444,研究方向为作物遗传育种,E-mail:1073195427@qq.com
