TAT-E4orf4 融合蛋白对胃癌细胞增殖、迁移和凋亡的影响
2021-03-04殷洁烽徐统球孙元水
殷洁烽 徐统球 张 喆 孙元水
胃癌是消化系统最常见的肿瘤之一,其发生与幽门螺杆菌感染和长期摄入腌制食品有关[1]。早期胃癌的治疗方法主要有内镜手术、腹腔镜手术、开放手术和综合治疗。对于未发生转移的胃癌患者,日本JGCA 指南推荐实施根治性手术[2]。然而我国早期胃癌发现率低,常错过最佳手术时机,而仅能使用卡培他滨+顺铂/奥沙利铂等药物行姑息性治疗[3]。化疗药物不仅价格昂贵,还可能导致恶心、呕吐、食欲减退或骨髓增生抑制等不良反应。腺病毒早期区4 第4编码蛋白(adenovirus early region 4 open reading frame 4 protein,E4orf4 protein)是一种新型的细胞毒因子,不仅参与腺病毒的基因剪切和表达等过程,而且能特异性杀伤癌细胞[4]。Brestovitsky 等[5]以pcDNAE4orf4 的表达载体诱导肿瘤细胞后,死亡率达到80%~90%。为研究E4orf4 对正常细胞的影响,Robert 等[6]以E4orf4 处理人来源肿瘤细胞和原代细胞,发现E4orf4 仅对肿瘤细胞具有杀伤作用,而对正常细胞作用影响很小。E4orf4 蛋白穿透细胞膜进入细胞内是E4orf4 发挥杀伤作用的前提,但是部分E4orf4 无法穿透细胞膜。转导肽能通过内吞方式进入细胞内,而且无细胞毒性[7]。本研究通过基因工程技术构建pET28-his-TAT-E4orf4 原核表达载体,研究TATE4orf4 融合蛋白对胃癌细胞增殖、迁移和凋亡的影响。
1 实验材料
1.2 试 剂 胰蛋白酶(批号90305)和RIMP 1640培养基(批号42401042)购自Invitrogen 公司;胎牛血清(批号10091148)购自复蒙公司。半胱氨酸天冬氨酸蛋白酶3(Caspase-3)(批号sc-7272)、半胱氨酸天冬氨酸蛋白酶9(Caspase-9)(批号sc-73548)、酪氨酸激酶Src(Sarcoma,Src)(批号sc-166860)、细胞外调节蛋白激酶(Extracellular Regulated Protein Kinases,ERK)(批号sc-271269)、磷酸化细胞外调节蛋白激酶(Phospho-Extracellular Regulated Protein Kinases,p-ERK)(批号sc-7383)、蛋白激酶B(Protein Kinase,BPKB 或AKT)(批号sc-5298)、细胞周期蛋白依赖激酶(cell division cycle 55p,CDC55p)(批号sc-1906)、酵母核苷二磷酸酶1(yeast nucleoside diphosphatase 1,YND 1)(批号sc-53506)和蛋白磷酸酶2A(protein phosphatase 2A,PP2A)(批号:sc-13601)鼠单克隆抗体购置Santa Cruz 公司,β-actin 鼠单克隆抗体(批号A01010)和HRP 标记的二抗(批号14-19-06)购自北京中杉金桥。蛋白提取试剂盒(批号BB-3101)及ECL 发光试剂盒(批号P10100)购自江苏凯吉生物公司;BCA 试剂盒(批号P0011)、Annexin VFITC 和PI 试剂盒(批号C1067M)购自碧云天公司。
2 实验方法
2.1 细胞培养 胃癌AGS 细胞株用含有10%的胎牛血清、1×104U/L 的青霉素和100mg/mL 的链霉素的RIMP 1640 培养基,置于37℃、5% CO2的CO2孵箱中培养。待细胞融合度为90%,用0.25%的胰酶消化,进行传代,取对数期细胞进行试验。
2.2 构建融合细胞穿膜肽的E4orf4 原核表达载体及鉴定 采用聚合酶链式反应(polymerase chain reaction,PCR)重组法克隆E4orf4 表达基因,将TAT 编码序列与之融合,构建融合分子TAT-E4orf4,同样以E4orf4 分子作对照。在两分子两端均加入6 个组氨酸(His)标签。利酶切链接法将融合分子克隆到原核表达载体pTAT-HA,转化到大肠杆菌DH5α,菌落PCR 筛选,酶切电泳和DNA 测序验证。将表达载体转化到BL21-DE3 菌株,菌落PCR 鉴定,IPTG 诱导蛋白表达,改变培养温度和诱导时间优化表达条件,免疫印迹鉴定目的蛋白,筛选高表达蛋白菌株。先用His-tag 亲和系统纯化,再用离子交换层析进一步纯化复性。
2.3 细胞增殖实验 按2.1 进行细胞培养,待胃癌AGS 细胞90%融合后,胰酶消化、离心,取100μL 胃癌AGS 细胞以浓度为1×104细胞/mL 接种在96 孔板中培养24h,设置四组,每组设五个重复孔。前三组分别加入含相等浓度的pET28、pET28-his-E4orf4 和pET28-his-TAT-E4orf4 蛋白纯化液,剩余一组加入等体积的RIMP 1640 培养基,设置为空白对照组。置于CO2孵箱孵育8h,每孔均加入20μL 的0.5%MTT 溶液,孵育4h 后终止培养,加入150μL 的 DMSO 后摇床上避光孵育15min。待结晶物溶解后,酶标仪检测OD 值。细胞活性值=(实验组平均吸光值-调零孔平均吸光值)/(对照组平均吸光值-调零孔平均吸光值)。
2.4 细胞划痕实验 如上设置分组,将胃癌AGS 细胞消化后离心,细胞悬液密度稀释到2×104个/mL。将Transwell 小室置于6 孔板中,AGS 细胞悬液接种于Transwell 上室,下室加入不含血清的RMIP 1640培养基,并加入等浓度的pET28、pET28-his-E4orf4和pET28-his-TAT-E4orf4,空白对照组加入等体积的不含血清的培养基。孵育24h,经洗涤、固定、染色后,倒置显微镜下观察胃癌AGS 细胞迁移的数量。随机选取5 条直径,在每一条直径上选取5 个视野拍照,采用Picpic 软件计数,比较穿过Transwell 小室的细胞平均数目。
2.5 流式细胞仪检测细胞凋亡 如上处理胃癌AGS 细胞后,收集各组细胞,经PBS 洗涤后重悬,加入Annexin V-FITC 避光孵育。1000r/min 离心5min,弃上清,重悬细胞后,加入PI 染色液,混匀后于4℃避光孵育,再次洗涤后,立即用流式细胞仪检测。
2.6 Western blot 检测Caspases、Src 和CDC55p 信号通路蛋白表达 如上处理四组胃癌细胞24h,先后以PBS 液洗涤、RIPA 裂解、离心后收集总蛋白,经BCA 法定量、变性、SDS 电泳、转膜、封闭和洗涤后,4℃过夜孵育一抗(1:1000),TBST 洗涤后,室温孵育二抗2h(1:200)。最后TBST 洗涤,加入增强化学发光液(ECL,enhanced chemiluminescence)液,置于凝胶成像系统中检测目的蛋白表达变化。
2.7 统计学方法 应用SPSS 17.0 统计软件数据分析,实验所得数据以均数±标准差()表示,上述所有实验均重复3 次以上。采用单因素方差分析,以P<0.05 为差异有统计学意义。
3 实验结果
3.1 pET28-TAT-E4orf4 原核表达载体的构建及诱导表达 经Xho I/Eco R 双酶切原核表达载体后,经琼脂糖电泳上的预期范围内出现一条的片段(见图1A)。通过对温度、pH、IPTG 浓度进行优化,经SDS-PAGE 电泳分析重组蛋白的表达。结果显示,1号、3 号、5 号和9 号菌株重组蛋白表达水平明显高于其他菌株(见图1B)。对上述菌株进一步培养,收集菌液,用His-tag 亲和系统纯化,再用离子交换层析进一步纯化复性,收集重组融合蛋白进行相关实验。
3.2 TAT-E4orf4 融合蛋白对胃癌细胞增殖的影响 采用MTT 法检测不同刺激后胃癌AGS 细胞的增殖能力。结果显示,相对于空白对照组(1.01±0.12),pET28 刺激后,胃癌AGS 细胞的增殖活性值为(1.03±0.26);而pET28-his-E4orf4 刺激后胃癌细胞增殖活性降低至(0.69±0.44),与pET28 组比较,差异有统计学意义(P<0.05)。融合细胞穿膜肽后(pET28-his-E4orf4 刺激),细胞增殖活性进一步下降,达到(0.41±0.06),与前两组比较,差异均有统计学意义(P<0.05)。
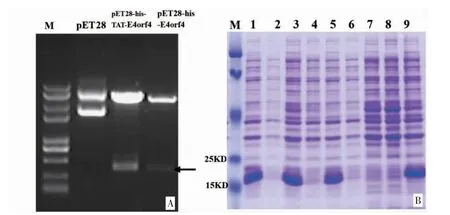
图1 pET28-TAT-E4orf4 表达载体的鉴定及诱导
3.3 TAT-E4orf4 融合蛋白对胃癌细胞迁移的影响 Transwell 实验结果发现,空白对照组的穿过Transwell 小室的细胞数目为(796.48±114.35)个,pET28 刺激后,穿过小室的细胞数目为(803.71±127.39)个,二者比较,差异无统计学意义(P>0.05)。经pET28-his-E4orf4 刺激后,AGS 细胞减少至(287.06±78.51)个,与前两组比较,差异有统计学意义(P<0.05)。而pET28-his-TAT-E4orf4 刺激,穿过小室的细胞数目仅为(163.21±36.77)个,与pET28-his-E4orf4 组比较,差异有统计学意义(P<0.05),见图2。

图2 TAT-E4orf4 融合蛋白对胃癌细胞迁移的影响(标尺=200μm)
3.4 TAT-E4orf4 融合蛋白对胃癌细胞凋亡的影响流式细胞仪检测结果发现,AGS 细胞凋亡率的变化趋势与MTT 结果相一致。空白对照组胃癌AGS 细胞的凋亡率为6.28%,pET28 刺激后胃癌AGS 细胞的凋亡率无明显增加。而pET28-his-E4orf4 刺激后,胃癌AGS 细胞上升至20.50%。而pET28-his-E4orf4 刺激后,凋亡率进一步增加,达到33.40%。见图3。
3.5 TAT-E4orf4 融合蛋白对胃癌细胞Caspase-3 和Caspase-9 表达的影响 采用Western blot 检测Caspase-3 和Caspase-9 的表达变化。经PET28、PET28-his-E4orf4 和PET28-his-TAT-E4orf4 刺激胃癌AGS 细胞后,Caspase-3 表达水平无明显变化,与Blank 组比较,差异无统计学意义(P>0.05)。与Caspase-3 变化趋势相似,四组胃癌AGS 细胞的Caspase-9 的表达水平差异也无统计学意义(P>0.05),见图4。

图3 TAT-E4orf4 融合蛋白对胃癌细胞凋亡的影响

图4 TAT-E4orf4 融合蛋白对Caspase-3 和Caspase-9 表达的影响
3.6 CPP-E4orf4 对胃癌细胞Src 信号通路蛋白表达的影响 PET28 刺激胃癌AGS 细胞后,Src 表达水平无明显变化,而pET28-his-E4orf4 刺激后Src 表达水平明显下降,与空白对照组和PET28 组比较,差异有统计学意义(P<0.05)。而融合细胞穿膜肽后,胃癌AGS 细胞的Src 表达水平进一步下降,与pET28-his-E4orf4 组比较,差异有统计学意义。与Src 变化趋势相反,pET28-his-E4orf4 和pET28-his-TATE4orf4 刺激胃癌AGS 细胞后ERK、p-ERK 和AKT表达水平均下降,而后者的下降趋势更明显,差异有统计学意义(P<0.05)。见图5。
4 讨论
胃癌细胞增殖和凋亡间的平衡紊乱是胃癌发生发展的重要分子机制[8]。胃癌细胞凋亡主要通过细胞内和细胞外两大途径,前者是由死亡信号诱导线粒体释放细胞色素C,与Caspase-9 形成凋亡复合体,使Caspase-9 酶原活化并释放,进而激活Caspase-3及其下游信号蛋白,而后者又包含Fas/Fas 配体、Caspase 家庭蛋白以及Src 介导的相关信号通路等途径[9-10]。

图5 细胞穿膜肽对Src-ERK-AKT 信号轴的影响
研究发现,胃癌组织促凋亡蛋白Caspase-3 表达含量明显低于癌旁组织[11]。有不少研究者试图通过生物治疗、放射治疗、细胞毒类药物和中医中药等多种方法来诱导胃癌细胞凋亡,抑制其增殖。Kim 等[12]报道低剂量的顺铂可激活Fas 受体,使Caspase-3 表达增加,促进胃癌细胞凋亡。Nishimura 等[13]利用腺病毒构建Caspase-8 表达载体,导入胃癌细胞株,其凋亡率显著增加。亦有研究证实Src 介导的相关信号通路在抑制肿瘤发生发展及影响细胞凋亡等生物学行为方面具有重要影响,靶向沉默Src 相关信号通路可调控胃癌细胞侵袭和迁移[14]。
近年发现部分溶瘤病毒编码的某些特殊蛋白具有很好的抗肿瘤作用,如鸡贫血病毒编码的凋亡素、自主细小病毒编码的非结构蛋白1 和腺病毒编码的E4orf4。其中,以E4orf4 研究最为深入。E4orf4 是由人腺病毒基因早期第4 转录区第4 开放读码框编码产生,含114 个氨基酸残基。E4orf4 蛋白与PP2A 或Src等分子伴侣作用,诱导细胞死亡[15]。E4orf4 与PP2A的B55 亚基的结合,能激活mTOR 信号通路,调控蛋白翻译。此外,Src 家族激酶可使E4orf4 发生磷酸化,继而在细胞质和细胞膜上聚集,导致双核肌动蛋白-肌球蛋白网络的组装重塑,细胞膜起泡,最终引起细胞死亡[15]。Zhou 等[16]构建EGF-E4orf4 表达载体,刺激胃癌BCG-823 细胞株后,结果发现BCG-823 细胞的增殖能力显著下降,对转移瘤小鼠的抑制率达到49%。另有文献报道,E4orf4 蛋白与分子伴侣Src相互作用可抑制Src-ERK-AKT 信号通路的激活,降低下游促凋亡蛋白(the proapoptotic BH3-onlyprotein,BAD)的磷酸化,增强BAD 与早期凋亡蛋白(Bcl-2 和Bcl-xl)相互作用,促进细胞凋亡[17]。
上述研究证实,E4orf4 对胃癌细胞具有很好的杀伤作用,而E4orf4 穿过细胞膜是其发挥作用的前提。目前常用其他试剂来增强细胞通透性,破坏细胞稳态。TAT 转导肽是来源于HIV 转录活化蛋白tat 的11 肽,通过内吞方式进入细胞内。将TAT 与目的蛋白制成的融合蛋白转导效率几乎可达到100%,而且无明显细胞毒性。基于上述研究现状,本研究将细胞穿膜肽与E4orf4 融合,构建pET28-his-TAT-E4orf4原核表达载体,分析TAT-E4orf4 融合蛋白对胃癌细胞增殖、迁移和凋亡的影响。结果发现,增加细胞穿膜肽后E4orf4 能显著抑制胃癌AGS 细胞增殖和迁移,促进AGS 细胞凋亡。Western blot 结果发现E4orf4 对胃癌细胞的杀伤作用及对凋亡的影响与Caspase 信号通路无关,但E4orf4 能活化Src-ERKAKT 信号通路,介导AGS 细胞凋亡,从而发挥抗肿瘤的效应。
