自交可育红花甜荞种质资源创新利用研究
2021-02-09李春花加央多拉田娟孙墨可王春龙董玉迪郭来春魏黎明孙连池任长忠
李春花 加央多拉 田娟 孙墨可 王春龙 董玉迪 郭来春 魏黎明 孙连池 任长忠
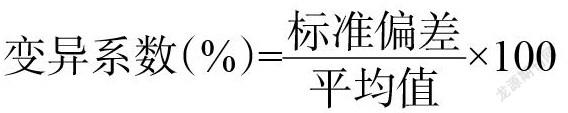





摘要:【目的】對等花柱型白花甜荞和异花柱型红花甜荞的杂交后代F2群体进行遗传分析,为甜荞资源创新及新品种选育提供理论依据。【方法】对等花柱型白花甜荞与短花柱型红花甜荞的杂交后代F2群体进行花色、花柱型、开花天数、生育天数和株高等7个农艺性状测定,并进行遗传分析及主成分分析。【结果】在F2群体中,白花单株占27.92%,粉红花和浅粉花单株占72.08%,表明红花对白花为不完全显性;同型花(等花柱型)和异型花(短花柱型和长花柱型)分离比为2∶1,可推测出等花柱型对异型花柱为显性。7个农艺性状中,开花天数、生育天数、主茎节数和茎粗的变异系数分别为9.97%、7.16%、9.88%和9.57%,小于10.00%;单株粒数、株高和一级分枝数的变异系数分别为59.85%、21.75%和21.80%,均大于20.00%。开花天数与株高呈显著负相关(P<0.05,下同),与一级分枝数呈显著正相关;生育天数与株高、主茎节数、一级分枝数和茎粗4个株型相关性状均呈不显著负相关(P>0.05,下同);4个株型相关性状中,株高与一级分枝数呈显著正相关,与主茎节数和茎粗均呈极显著正相关;单株粒数作为产量性状,与开花天数呈极显著负相关,与生育天数呈显著负相关,与4个株型相关性状均呈极显著性正相关。主成分分析的前2个因子的特征值大于1.000,累积贡献率为56.083%,反映了所测农艺性状的主要信息,第1公因子主要是由株高、主茎节数、茎粗和单株粒数4个因子决定,故称为株型因子;第2公因子中开花天数的因子载荷最大,故称为生殖因子,这2个公因子既相互促进,又相互制约。F2群体的各农艺性状中均出现超亲分离,虽然未出现粉红色等花柱型单株,但出现了70株浅粉色等花柱型的单株,其中8株为既比父本早熟又比母本高产的浅粉色等花柱型。【结论】开花天数、生育天数、主茎节数和茎粗的变异较小,稳定性较高,而单株粒数、株高和一级分枝数的变异较高,稳定性较低。在等花柱甜荞与红花甜荞的杂交育种中应协调好生殖因子和株型因子的关系,要根据各性状间的相关性进行有目的地选择,有望获得早熟高产稳产的红花甜荞。
关键词:红花甜荞;等花柱资源;遗传多样性;杂交后代;资源创新
中图分类号:S517.035.1 文献标志码: A 文章编号:2095-1191(2021)10-2751-07
Abstract:【Objective】The genetic analysis of F2 population of hybrid progenies of homo-flower white buckwheat and heteromorphous safflower buckwheat would provide a theoretical basis for resource innovation and new varieties of common buckwheat. 【Method】 Seven agronomic traits such as flower color,style type,flowering days,number of growing days,plant height were measured on the F2 populations of hybrid progenies of homo-flower white buckwheat and short style safflower buckwheat, and carried out genetic diversity and principal component analysis.【Result】In the F2 population,individual plants with white flowers accounted for 27.92%,pink flowers and light pink flowers accounted for 72.08%,indicating that safflower was not completely dominant to white flowers. In addition,the separation ratio of homotypes (equal styles) and heteromorphic flowers (short styles,long styles) was 2∶1,and it could be inferred that the isome-tric styles were dominant heteromorphic styles. Among the 7 agronomic traits,the coefficients of variation of flowering days,growing days,main stem node number and stem diameter were 9.97%,7.16%,9.88% and 9.57%,respectively,which were less than 10.00%; the coefficients of variation of number of seeds per plant, plant height and number of first branch were 59.85%,21.75% and 21.80%, respectively, which were all greater than 20.00%. The flowering days was significantly negatively correlated with plant height (P<0.05, the same below), and was significantly positively correlated with the number of first branch; the number of growing days were not significantly negatively correlated with plant height, number of the main stalk section, number of first branch and stem diameter (P>0.05, the same below). Among the four plant type-related traits, plant height was significantly positively correlated with the number of first branch, and extremely significantly positively correlated with the number of the main stalk section and stem diameter. As a yield trait, the number of seeds per plant was extremely negatively correlated with flowering days, was significantly negatively correlated with number of growing days, and was extremely significantly positively correlated with four plant type-related traits. The eigenvalues of the first two factors in the principal component analysis were greater than 1.000,and the cumulative contribution rate was 56.083%,which reflected the main information of the agronomic traits measured. The first common factor was mainly determined by four factors:plant height, number of the main stalk section, stem diameter and number of seeds per plant, and was called plant type factor; the second common factor had the largest load of flowering days, which was called reproductive factor. These two common factors both promoted and restricted each other. Super parental separation was found in all agronomic traits of F2 population. Although no pink homo-flower individual plants were found, but 70 light pink homo-flower individual plants were observed. Eight of them were pink homo-flower individual plants which were earlier than their fathers and more productive than their mother. 【Conclusion】 The variations of flowering days, number of growing days, number of the main stalk section and stem diameter are small and stability is high, while the variation of number of seeds per plant, plant height and number of first branch of first order is high and stability is low. In the cross breeding of homo-flower white buckwheat and heteromorphous safflower buckwheat, the relationship between reproductive factors and plant type factors should be well coordinated, and the target selection should be carried out according to the correlation of each trait, so as to obtain safflower buckwheat with early maturity, high yield and stable yield.
Key words:safflower buckwheat; isostyle resource; genetic diversity; hybrid offspring; resource innovation
Foundation item: National Natural Science Foundation of China (31860412); National Technical System Program of Oat and Buckwheat Industry(CARS-07-C-2)
0 引言
【研究意义】甜荞(Fagopyrum esculentum Monch,2n=16)为蓼科(Polygonaceae)荞麦属(Fagopyrum)的一年生短日照植物,是花柱异长自交不亲和的虫媒和风媒传粉作物(陈稳良等,2020),具有生育期短、耐旱耐贫瘠等特点,是我国大部分作物无法生长的自然环境恶劣地区的主要粮食和经济作物,也可根据当地情况作为填闲作物或救灾防荒作物(何健等,2002;杨耐德,2006;Alamprese et al.,2007;Jiang et al.,2007)。红花甜荞是我国甜荞主产区的主栽品种,广泛分布于陕甘宁及内蒙古长城沿线区,其籽粒红色,营养丰富、品质佳、口味特殊,具有多种人体所需的营养物质,开花时呈粉红色或红色而得名,且花色鲜艳、花期长、花色一致稳定,在观光、美化环境、插花艺术等方面也具有广阔的应用前景(冯佰利等,2005;林汝法等,2005)。因此,开展等花柱甜荞与红花甜荞杂交后代的遗传多样性分析对其培育花型、花色等性状稳定的自交可育观赏型红花甜荞品种具有重要意义。【前人研究进展】由于甜荞依赖虫媒和风媒来实现授粉结实,加之近年来日本白花甜荞品种在我国荞麦主产区的推广种植,最终导致农田种植的甜荞品种间易发生杂交,导致花色、粒色等纯合性缺失(鲍国军和曹亚凤,2013),天然结实率低,仅为4%~20%,产量低且不稳定(方正武和刘志雄,2015;张柯彬等,2017;陈稳良等,2020)。自1987年,陆续在云南、西藏、四川等地发现了等花柱(即雌雄蕊等长)的自交可育野生甜荞资源(陈庆富,2012;杨丽娟和陈庆富,2018),在隔离条件下,其自交结实率远大于栽培甜荞,在中、高密度种植条件下表现出高产特性,产量高于普通栽培品种(赵绪明,2015)。在荞麦的表型中,花柱性状是可稳定表达遗传的性状,在群体中易于观察分类,且该性状与甜荞的自交结实性密切相关,因此,该特性有利于选育农艺性状稳定的自交可育早熟高产红花甜荞品种(Chen,2016)。栽培甜荞普遍是异型花,在一个群体中通常有两种花柱类型,即长花柱型和短花柱型,是根据雌雄蕊的长度所分,其中短花柱型表现为雌蕊短,雄蕊长,雄蕊的平均长度为66.94 µm,雌蕊平均长度18.22~30.19 µm;长花柱型表现为雌蕊长,雄蕊短,雄蕊的平均长度为35.37 µm,雌蕊平均长度为47.69~64.08 µm,植株个体均是异型异株,而等花柱甜荞是同型花,仅有一种类型,即等花柱型,雌雄蕊长度相同,雌蕊和雄蕊平均长度为35.02~50.78 µm,在野生甜荞中此类花型较常见(侯亚芳,2018)。研究表明,栽培甜荞的花柱性狀是由S位点控制,短花柱型相对于长花柱型为显性,基因型为SS和Ss,长花柱型甜荞基因型为ss(Shaikh et al.,2002;Wang et al.,2005;Fesenko,2010)。等花柱甜荞的花柱性状由Sh位点控制,基因型为ShSh和Shsh,而Sh位点与S位点可能为等位位点,显性关系表现为SS和Ss对Sh为显性,Sh对ss为显性(Wang et al.,2005)。杜莹(2015)对栽培甜荞与等花柱甜荞的杂交后代花柱进行调查,结果发现后代花型可细分为完全等花柱型、稍长花柱型、长花柱型、稍短花柱型和短花柱型5种,表明栽培甜荞与等花柱甜荞的杂交后代花型种类较丰富。郭超(2016)在等花柱型甜荞与栽培甜荞的杂交F3和F4代中发现,5种花柱类型在分离比例、结实性和单株产量上均表现出显著差异,表明可通过改良花柱类型提高自交结实率,从而提高产量。【本研究切入点】针对目前红花甜荞和栽培甜荞在田间发生杂交导致花色、粒色等不纯合,以及授粉依赖昆虫而导致红花甜荞产量不稳定且偏低的问题,培育自交可育红花甜荞是非常有效的解决方法,但目前鲜见研究报道。【拟解决的关键问题】对等花柱型白花甜荞与短花柱型红花甜荞的杂交后代F2群体进行花色、花柱型、开花天数、生育天数和株高等7个农艺性状测定,并进行遗传分析及主成分分析,寻求自交可育红花甜荞新资源创新手段和新品系筛选方法,为培育性状稳定的早熟高产自交可育的观赏型红花甜荞新品种打下基础。
1 材料与方法
1. 1 试验材料
母本材料BQ-98是由吉林省白城市农业科学院燕麦工程研究中心在多年承担国家区域试验中发现,再经本课题组筛选、纯化的白色花等花柱型可育新品系。父本材料西农T1351是西北农林科技大学农学院培育而成的红色异花柱型品种,该品种花红色鲜艳、叶脉红色、主茎紫红色,籽粒形成至乳熟期颜色为红色,持续时期10~20 d。
1. 2 试验方法
试验在吉林省白城市农业科学院的温室(122º47′E,45º37′N)进行。试验地属温带大陆性季风气候,年平均降水量为399.9 mm,无霜期平均为144 d。温室内设有地暖、大型抽风机、侧窗和顶窗,自动控制日照仪,可以增强空气流通,及时调节室内温湿度和日照长度。于2019年9月将母本材料BQ-98和父本材料西农T1351分别种植在营养钵中,每盆种3粒,出苗后的10~20 d每盆留1株生长健壮的植株,培育至开花初期进行杂交,具体操作:在头戴式放大镜下用镊子对开花前未授粉的母本花蕾进行去雄,同时取短花柱父本已开裂花朵的花药,将花药的花粉涂抹于已去雄的母本雌蕊柱头上进行授粉,再套袋,待籽粒成熟后即得杂交种子。杂交成功与否是通过以F1代植株的花色判断,白色花为杂交失败,粉色花为杂交成功。于2020年3月将杂交种子20粒分别种植在20个营养钵中,开花期淘汰白色花的单株及病株、劣株,剩余15株F1代单株成功自交获得F1代自交种子,并分别收获保存。从15株F1代单株中选取单株粒数最多(154粒)的F1代自交种子,于2020年7月与亲本各12粒进行盆栽,每盆1粒。
1. 3 测定项目
开花期调查花柱类型(长花柱、短花柱和等花柱)、花色和开花天数(出苗到初花天数)。待籽粒70%~80%成熟时收获,并调查记录生育天数(出苗到收获天数)、株高、主茎节数、一级分枝数、茎粗和单株粒数。
1. 4 统计分析
利用Excel 2010对调查的农艺性状数据进行整理分析,使用SPSS 19.0进行主成分分析和相关分析。利用表型性状的变异系数(CV)表示表型性状离散程度,计算公式:
2 结果与分析
2. 1 甜荞亲本及杂交后代F2群体花柱和花色类型的分离表现
由表1可知,母本材料全部为白色等花柱型,父本材料中有8株粉红色短花柱型,4株为粉红色长花柱型;在F2群体中,白花单株为43株,占27.92%,其中,等花柱型为33株,短花柱型为8株,长花柱型为2株;粉红花单株为4株,占2.60%,均为短花柱型;浅粉花单株为107株,占69.48%,其中等花柱型为70株,短花柱型为30株,长花柱型为7株。由此还可知,在F2群体中,白花单株占27.92%,粉红花单株和浅粉花单株占72.08%,表明红花对白花为不完全显性;等花柱型单株为103株,占66.88%,短花柱型单株为42株,占27.27%,长花柱型单株为9株,占5.84%,等花柱型远多于短花柱型和长花柱型,且同型花(等花柱型)和异型花(短花柱型和长花柱型)分离比为2∶1,推断等花柱型相对于异花柱型为显性;在F2群体中浅粉色等花柱型的单株为70株,占45.45%,表明从杂交后代中有望筛选出粉色花等花柱型的单株。
2. 2 甜荞亲本及杂交后代F2群体的农艺性状表现
由表2可知,在开花天数、生育天数、株高、主茎节数、一级分枝数、茎粗和单株粒数7个农艺性状方面,F2群体的变异范围均超过父母本的变异范围,且母本的开花天数、生育天数、一级分枝数和单株粒数平均值大于父本,但父本的株高、主茎节数和茎粗平均值大于母本,F2群体的这7个农艺性状平均值均在两亲本之间;母本的7个农艺性状变异系数均比父本小,而F2群体的7个农艺性状变异系数大于两亲本。F2群体的开花天数、生育天数、主茎节数和茎粗变异系数分别为9.97%、7.16%、9.88%和9.57%,均小于10.00%,单株粒数、株高和一级分枝数的变异系数分别为59.85%、21.75%和21.80%,均大于20.00%。综上所述,开花天数、生育日数、主茎节数和茎粗的变异小,遗传稳定性较高;而单株粒数、株高和一级分枝数的遗传多样性丰富,适宜筛选优质材料。
2. 3 杂交后代F2群体农艺性状间的相关分析结果
由表3可知,开花天数与生育天数呈正相关,但未达显著水平(P>0.05,下同),与株高呈显著负相关(P<0.05,下同),与一级分枝数呈显著正相关;生育天数与株高、主茎节数、一级分枝数和茎粗4个株型相关性状均呈负相关,未达显著水平;4个株型相关性状中,株高与一级分枝数呈显著正相关,与主茎节数和茎粗均呈极显著正相关(P<0.01,下同);单株粒数作为产量性状,与开花天数呈极显著负相关,与生育天数呈显著负相关,与4个株型相关性状均呈极显著性正相关,其中,株高与单株粒数的相关系数最大,为0.515%。
2. 4 主成分分析结果
对杂交后代F2群体的7个农艺性状进行主成分分析,结果如表4所示。前2个因子的特征值大于1,且对总方差的贡献最大,其特征值分别为2.634和1.292,总方差贡献率分别为37.625%和18.458%,累积方差占总方差的56.083%。第3~7个因子的特征值均小于1.000,忽略不计,只保留前2个公因子作进一步分析。
对公因子进行方差最大正交旋转,得到旋转后因子的载荷矩阵如表5所示。第1公因子主要是由株高、主茎节数、茎粗和单株粒数4个因子决定,其因子载荷分别为0.787、0.706、0.715和0.709,故称为株型因子;第2公因子中开花天数的因子载荷最大,为0.847,故称为生殖因子。综上所述,这2个公因子既相互促进,又相互制约,因此,在荞麦高产育种中应协调好二者的关系。
2. 5 目标单株选择
目标单株为既比父本早熟又比母本高产的粉红色等花柱型的单株。虽然F2群体中未出现粉红色等花柱型单株,但出现了70株浅粉色等花柱型的单株,其中8株为既比父本早熟又比母本高产的浅粉色等花柱型(图1),表明通过等花柱甜荞与红花甜荞的杂交有望获得早熟高产稳产的红花甜荞。
3 讨论
本研究结果表明,在杂交后代F2群体中出现同型花(等花柱型)和异型花(短花柱型和长花柱型)的分离比为2∶1,由此推测等花柱型对异型花柱型为显性,与侯亚文(2018)研究发现等花柱型相对于长花柱型为显性的结果一致,而与Wang等(2005)研究发现短花柱型相对于等花柱型显性的结果不一致,并且分离比不符合孟德尔分离规律,其原因可能有等花柱型虽然表现出自交亲和性,但其仍保留自交不亲和性的基因(Yasui et al.,2012),且控制花型的S位点基因作用机理比较复杂(Matsui et al.,2003)而所导致。本研究结果还表明,F2群体的不同农艺性状中均出现超亲分离现象,说明可在后代中筛选出超亲性状的优良株系。变异系数是性状遗传多样性的具体体现,变异系数越大,遗传背景越丰富,越有利于品种选育(Alizadeh et al.,2015)。此外,7个农艺性状中,单株粒数的变异系数最大,为59.85%,表明单株粒数的遗传多样性最丰富,具有较大的改良潜力;生育天数的变异系数最小,为7.16%,表明其他性状具有较小的变异性和较高的稳定性。
Li等(2012)研究认为,苦荞开花天数与株高呈显著正相关。李月等(2013)研究认为,苦荞主茎分枝数与主茎节数和单株粒数呈显著正相关;主茎节数与株高和单株粒数呈显著正相关;株高与单株粒数呈显著正相关。汪灿等(2014)研究認为,甜荞株高与主茎节数和分枝数呈显著正相关;主茎节数与分枝数呈显著正相关;株高、主茎节数和分枝数与单株粒数无显著相关。侯亚芳(2018)研究认为,甜荞株高与节数、一级分枝数和单株粒数呈显著正相关;节数与一级分枝数和单株粒数呈显著正相关;一级分枝数与单株粒数呈显著正相关。本研究结果表明,株高与主茎节数、一级分枝数、茎粗和单株粒数呈显著正相关;开花天数与株高呈显著负相关,与前人的研究结果较相似,显著水平存在差异是可能与环境不同有关,而开花天数和株高的相关性差异可能与材料不同有关。此外,孟第尧和张先炼(1998)研究认为,在甜荞新品种选育时,应以单株粒数、丰产株型和千粒重为主要选育目标。本研究通过主成分分析将农艺性状分为株型因子和生殖因子2个成分,这2个决定因子从不同角度反映甜荞各性状间的关系,与孟第尧和张先炼(1998)研究结果基本一致。
4 结论
开花天数、生育天数、主茎节数和茎粗的变异较小,稳定性较高,而单株粒数、株高和一级分枝数的变异较高,稳定性较低。在等花柱甜荞与红花甜荞的杂交育种中应协调好生殖因子和株型因子的关系,要根据各性状间的相关性进行有目的地选择,有望获得早熟高产稳产的红花甜荞。
参考文献:
鲍国军,曹亚凤. 2013. 红花甜荞麦新品种平荞7号选育[J]. 甘肃农业科技,(5):3-5. [Bao G J,Cao Y F. 2013. Breeding report of a new safflower buckwheat variety Pingqiao 7[J]. Gansu Agricultural Science and Technology,(5):3-5.] doi:10.3969/j.issn.1001-1463.2013.05.001.
陈庆富. 2012. 荞麦属植物科学[M]. 北京:科学出版社. [Chen Q F. 2012. Plant sciences on genus Fagopyrum[M]. Beijing:Science Press.]
陈稳良,李秀莲,史兴海,梁改梅,刘龙龙,赵建栋,秦秀珍. 2020. 甜荞等花柱资源与栽培甜荞杂交初步研究[J]. 植物遗传资源学报,21(4):1030-1035. [Chen W L,Li X L,Shi X H,Liang G M,Liu L L,Zhao J D,Qin X Z. 2020. A preliminary study on the hybridization of common buckwheat isostyle resources and cultivated buckwheat[J]. Journal of Plant Genetic Resources,21(4):1030-1035.] doi:10.13430/j.cnki.jpgr.20191219002.
杜莹. 2015. 甜荞不同花柱类型杂交后代遗传表现及自交不亲和性的SSR标记分析[D]. 杨凌:西北农林科技大学. [Du Y. 2015. Heterosis performance and self-incompatibility using SSR markers in different flower of common buckwheat pillar type[D]. Yangling:Northwest A & F University.]
方正武,刘志雄. 2015. 甜荞花同源异型基因FeMADS1的克隆和序列结构分析[J]. 西南农业学报,24(3):82-87. [Fang Z W,Liu Z X. 2015. Cloning and sequence analysis of FeMADSI gene from Fagopyrum esculentum[J]. Acta Agriculturae Boreali-Occidentalis Sinica,24(3):82-87.] doi:10.7606/j.issn.1004-1389.2015.03.013.
冯佰利,姚爱华,高金峰,高小丽,柴岩. 2005. 中国荞麦优势区域布局与发展研究[J]. 中国农学通报,21(3):375-377. [Feng B L,Yao A H,Gao J F,Gao X L,Chai Y. 2005. Study on regional distribution and development of buckwheat in China[J]. Chinese Agricultural Science Bulletin,21(3):375-377.]
郭超. 2016. 自花结实甜荞杂交后代花柱类型、结实率分离及其SSR标记[D]. 杨凌:西北农林科技大学. [Guo C. 2016. The style types and segregation of fecundity and SSR molecular marker about the hybrids from the self-fertile common buckwheat[D]. Yangling:Northwest A & F University.]
何健,张国治,张虹,次仁欧珠. 2002. 荞麦营养成分的检测及分析[J]. 河南农业大学学报,36(3):302-304. [He J,Zhang G Z,Zhang H,Ciren O Z. 2002. Determination and analysis of nutrient ingredients of buckwheat[J]. Journal of Henan Agricultural University,36(3):302-304.] doi:10.16445/j.cnki.1000-2340.2002.03.026.
侯亚方. 2018. 自交可育甜荞杂交后代主要性状的遗传特性分析[D]. 杨凌:西北农林科技大学. [Hou Y F. 2018. The inheritance analysis of main agronomic traits about the hybrids from the self-fertile buckwheat[D].Yangling:Northwest A & F University.]
李月,石桃雄,黃凯丰,唐晓辛,何娟,简永,陈庆富. 2013. 苦荞生态因子及农艺性状与产量的相关分析[J]. 西南农业学报,26(1):35-41. [Li Y,Shi T X,Huang K F,Tang X X,He J,Jian Y,Chen Q F. 2013. Correlation analysis of tartary buckwheat seed yield with ecological factors and agronomic traits[J]. Southwest China Journal of Agricultural Sciences,26(1):35-41.] doi:10.16213/j.cnki.scjas.2013.01.031.
林汝法,柴巖,廖琴,孙世贤. 2005. 中国小杂粮[M]. 北京:中国农业科学技术出版社. [Lin R F,Chai Y,Liao Q,Sun S X. 2005. Minor grain crops in Chiana[M]. Beijing:China Agricultural Science and Technology Press.]
孟第尧,张先炼. 1998. 普通甜荞产量的主成分分析[J]. 上海师范大学学报:自然科学版,27(4):50-53. [Meng D Y,Zhang X L. 1998. Principal component analysis of yield of common buckwheat[J]. Journal of Shanghai Normal University:Natural Science Edition,27(4):50-53.]
汪灿,阮仁武,袁晓辉,易泽林. 2014. 甜荞农艺性状与产量关系的多重分析[J]. 中国农学通报,30(12):234-238. [Wang C,Ruan R W,Yuan X H,Yi Z L. 2014. Multiple analysis of relationship between agronomic traits and yield in Fagopyrum esculentum[J]. Chinese Agricultural Science Bulletin,30(12):234-238.]
杨丽娟,陈庆富. 2018. 荞麦属植物遗传育种的最新研究进展[J]. 种子,37(4):52-58. [Yang L J,Chen Q F. 2018. Recent advances in genetic breeding of buckwheat[J]. Seed,37(4):52-58.] doi:10.16590/j.cnki.1001-4705.2018.04.052.
杨耐德. 2006. 荞麦的营养特性及其加工技术探讨[J]. 农产品加工,(5):47-48. [Yang N D. 2006. Discuss on the nutrient characteristic and process technology of buckwheat[J]. Academic Periodical of Farm Products Processing,(5):47-48.] doi:10.3969/j.issn.1671-9646-B.2006.05.013
张柯彬,陈炳全,刘志雄. 2017. 甜荞FaesAP2基因的克隆与表达分析[J]. 植物科学学报,35(3):354-361. [Zhang K B,Chen B Q,Liu Z X. 2017. Cloning and expression analysis of the FaesAP2 gene from Fagopyrum esculentum(Polygonaceae)[J]. Plant Science Journal,35(3):354-361.] doi:10.11913 /PSJ.2095-0837.2017.30354.
赵绪明. 2015. 等花柱甜荞结实性表现及生产潜力研究[D]. 杨凌:西北农林科技大学. [Zhao X M. 2015. The study on the performance of fecundity and potential productivity of the same type of flowers[D]. Yangling:Northwest A & F University.]
Alamprese C,Casiraghi E,Pagani M A. 2007. Development of gluten-free fresh egg pasta analogues containing buckwheat[J]. European Food Research and Technology,225: 205-213. doi:10.1007/s00217-006-0405-y.
Alizadeh K,Fatholahi S,Jaime A. 2015. Variation in the fruit characteristics of local pear(Pyrus spp.) in the northwest of Iran[J]. Genetic Resources and Crop Evolution,62(5):635-641. doi:10.1007/s10722-015-0241-7.
Chen Q F. 2016. Recent progresses on interspecific crossbreeding of genus Fagopyrum Mill[C].The 13th International Symposium on Buckwheat. Korea:Chungbuk National University:285-292.
Fesenko I N. 2010. Inheritance of flower homostyly of auto-gamous species Fagopyrum tataricum Gaertn. in interspecific crosses with heterostylous cross-pollinator F. cymosum Meisn[J]. Russian Agricultural Sciences,36(5):324-326. doi:10.3103/S1068367410050022.
Jiang P,Burczynski F,Campbell C,Pierce G,Austria J A,Briggs C J. 2007. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum,F. tataricum,and F. homotropicum and their protective effects against lipidperoxidation[J]. Food Research International,40(3):356-364. doi:10.1016/j.foodres.2006.10.009.
Li C H,Kobayashi K,Yoshida Y, Ohsawa R. 2012. Genetic analyses of agronomic traits in Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn.)[J]. Breeding Science,62(4):303-309. doi:10.1270/jsbbs.62.303.
Matsui K,Tetsuka T,Nishio T,Hara T. 2003. Heteromorphic incompatibility retained in self-compatible plants produced by a cross between common and wild buckwheat[J]. New Phytologist,159(3):701-708. doi:10.1046/J. 1469-8137.2003.00840.X.
Shaikh N,Guan L,Adachi T. 2002. Failure of fertilization associated with absence of zygote development in the interspecific cross of Fagopyrum tataricum×F. esculentim[J]. Breeding Science,52(1):9-13. doi:10.1270/jsbbs. 52.9.
Wang Y J,Scarth R,Campbell C. 2005. Sh and Sc-two complementary dominant genes that control self-compatibility in buckwheat[J]. Crop Science,45(4):1229-1234. doi:10.2135/cropsci2003.0560.
Yasui Y,Mori M,Aii J,Abe T,Matsumoto D,Sato S,Hayashi Y,Ohnishi O,Ota T. 2012. S-loeus early flowering 3 is exclusively present in the genomes of short-styled buckwheat plants that exhibit heteromorphic self-incompatibility[J]. PLoS One,7(2):e31264. doi:10.1371/journal.pone.0031264.
(責任编辑 陈 燕)
