大青杨PuZFP103基因的序列特征及逆境胁迫的表达分析
2021-02-02马苗苗吕婉秋LEEJoobin杨静莉
张 馨,马苗苗,吕婉秋,LEE Joobin,杨静莉
(东北林业大学林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040)
大青杨(Populusussuriensis)是我国东北地区营造速生丰产林的主要树种之一[1]。该树种用途广泛,其木材可供建筑、舟船、造纸业、家具用胶合板等使用,很多国家已将其列为重要工业用材树种之一,并且大青杨也是恢复森林生态环境及短周期工业用材林最为理想的树种[2]。该树种喜光、耐寒、速生,且有极强的适应能力,也因为其容易成活等特点,被认为是造林成效最显著的树种之一[3]。由于大青杨抗逆性强,生长速度快,木材积蓄积量大,所以是生产纸浆材的重要造林树种[4]。近年来,由于气候变化,干旱和高盐等非生物胁迫已严重影响林木的生长发育,其引起的林木死亡现象在世界各地频繁发生[5],所以提高树木抗逆性相关问题越来越受到人们的重视。目前,通过分子及基因组分析等方法已经鉴定了很多植物响应逆境胁迫的基因,包括调控胁迫诱导基因表达的转录因子(transcription factor,TF)[6]。转录因子通过调控下游复杂的信号网络以响应逆境胁迫的植物信号转导并从中发挥着重要作用[7]。
锌指蛋白(zinc finger proteins,ZFP)是一类含有锌指结构域的蛋白,是植物中最大的一类转录因子家族[8]。在真核生物中,锌指蛋白属于TF家族,具有高度保守的锌指结构域[9]。ZFP可通过ZF结构域与DNA、 RNA或其他调控元件相互作用[10]。根据半胱氨酸残基和组氨酸残基的数量和位置,锌指蛋白可以分为C2H2、C2HC、C2HC5、C2C2、CCCH、C3HC4、C4、C4HC3、C6 和 C8等不同亚类[11]。越来越多的研究表明,C2H2型锌指蛋白在植物应激反应中起着非常重要的作用。很多与植物抗逆性相关的C2H2型锌指蛋白已被鉴定[4]。目前,有关C2H2转录因子在植物中的功能研究主要局限于拟南芥(Arabidopsisthaliana)和水稻(Oryzasativa)等模式植物中,在杨树基因功能研究中鲜见。因此,后续的研究工作需进一步探讨ZFP基因家族在大青杨中的功能和作用机理。研究发现,矮牵牛 (Petuniahybrid) Zinc pyrithione Protein(ZPT)是植物界第一个被报道的C2H2型ZFP[12]。因此,C2H2型ZFP在植物中也被称为ZPT,通常有1~4个ZFs,而其含有2个C2H2型锌指结构域的ZPT2家族也可以说是ZPT中最大的亚类之一[13]。目前,很多研究已证实了ZPT2蛋白在植物响应非生物胁迫的应答过程中起至关重要的作用[14]。在不同的非生物胁迫处理下,水稻OsZFP182、OsZFP252、OsZFP245和OsZFP179等基因的表达水平都稳定增加,这些基因的过表达也明显改善了转基因水稻的抗旱性、耐寒性和耐盐性[15]。其中OsZFP179参与植物体内对盐胁迫的应答,OsZFP252响应盐和干旱,过表达OsZFP252可以提高水稻植株的脯氨酸和可溶性糖含量,而且盐和干旱胁迫相关的应答基因表达量都明显上调[16]。甘薯(Dioscoreaesculenta)bZFP1基因通过参与调控ABA信号转导、脯氨酸合成及活性氧清除进而增强了转基因拟南芥对盐胁迫与干旱胁迫的耐性[17]。目前研究最多的是属C2H2 型锌指蛋白,而C2H2 型锌指家族的基因又可以分为Q、M、C和Z类型[18-19]。其中,Q 型 C2H2 型锌指家族的基因功能研究最多[20]。例如,ZFP245是水稻中Q型锌指蛋白基因,过表达ZFP245通过调节脯氨酸含量和清除活性氧来提高水稻对低温和干旱胁迫的耐受性[21]。ZFP179也属于这类基因,ZFP179的过表达显著提高了水稻的耐盐性[22]。以上研究表明,C2H2锌指蛋白转录因子在植物抗逆性中起着重要作用,特别是在盐胁迫和干旱胁迫下。本研究的PuZFP103属于Q 型 C2H2 型锌指家族的基因。
本试验以大青杨为材料克隆出PuZFP103基因,对其氨基酸序、保守结构域等进行了生物信息学分析,并利用(real time quantitative polymerase chain reaction, RT-qPCR)分析了PuZFP103在NaCl、PEG6000及脱落酸(abscisic acid,ABA)非生物胁迫处理下的表达特性,为进一步明确PuZFP103响应逆境的生物学功能提供依据。
1 材料与方法
1.1 植物材料
试验材料取自东北林业大学林木遗传育种国家重点实验室保存的野生型大青杨,均采集于东北林业大学校园内。用升汞(HgCl2)和次氯酸钠(NaClO)进行消毒和扩繁,并由植物组织培养获得无菌大青杨,置于25 ℃组培室培养,培养基为1/2 MS培养基,光周期为16 h光照/8 h黑暗,待植株长到8~10 cm选取同一无性系大青杨植株,用于以下试验。
1.2 试验方法
1.2.1 大青杨RNA的提取及cDNA的合成
采用CTAB法提取3 周大的无菌大青杨的总RNA,使用南京诺唯赞生物生产的HiScript©Ⅱ Q RT SuperMix for qPCR (+gDNA wiper) (www.vazyme.com)按照说明书将其反转录成cDNA。反转录成功的cDNA置于-20 ℃保存待用。
1.2.2 大青杨PuZFP103基因的克隆
本试验参照毛果杨(P.trichocarpa)基因组(https://phytozome.jgi.doe.gov/pz/portal.html)序列对其中同源的一个成员进行克隆及生物信息学分析,并将其命名为PuZFP103。引物设计应用NCBI引物设计在线网站(www.ncbi.nlm.nil.gov/tools/primer-blast)和Primer Premier 5.0软件。在PuZFP103基因CDS序列两端分别设计引物(正向引物为5′-ATGGATTTCCAACCAAACAC-3′以及反向引物为5′-AAGCCTTAAAGACAAATCTA-3′),本试验所涉及的引物及测序均由哈尔滨擎科生物公司(www.tsinke.com)合成。以合成的cDNA为模板,使用北京全式金生物技术有限公司(www.transgen.com.cn)2×EasyTaqPCR SuperMix 克隆试剂盒进行PCR。反应体系为:cDNA模版1 μL、正反向引物各1 μL、2×EasyTaqPCR SuperMix 10 μL、去离子水7 μL,反应体系为20 μL。PCR反应条件为:94 ℃预变性2 min,94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸15 s,共35个循环,72 ℃延伸2 min。该基因全长共507 bp,经PCR扩增反应后,将其产物经过1.2%琼脂糖凝胶电泳检测,将得到的目的条带与DL 2000 DNA Marker对比,条带单一,大小符合,为目的条带。接下来使用购自OMEGA公司(www.omegabiotek.com.cn)快速琼脂糖凝胶DNA回收试剂盒回收目的条带。将胶回收产物连接到PEASY-T1载体购自中美泰和生物技术(北京)有限公司(taihegene.com),置于恒温培养箱25 ℃(购自上海精宏试验设备有限公司),5 min连接,使用购自OMEGA公司(www.omegabiotek.com.cn)核酸纯化试剂盒将其纯化。纯化产物利用热激法转化大肠杆菌,转化完毕的菌液涂至含有浓度卡那霉素(Kan, Kanamyceticus)的LB固体培养基平板,倒置于37 ℃恒温培养箱恒温过夜培养。第2 天早上挑取单克隆,用1 mL不加卡那霉素的LB液体培养基摇菌后送到测序公司进行测序。保存测序成功的菌液并摇菌进一步提取质粒待用。
1.2.3 植物表达载体pBI121-PuZFP103-GFP的构建
1)设计载体的双酶切引物。通过分析杨树PuZFP103基因全长序列及pBI121-GFP载体图谱设计本基因可用的双酶切引物,笔者找到pBI121-GFP(载体来自本实验室保存)载体图谱的多克隆位点,从中找出该载体可用的全部酶切位点,再将杨树PuZFP103基因全长CDS序列输入到BioEdit软件中,找出不能用于该基因的酶切位点,最后排除两者共有的酶切位点进行双酶切引物的设计。根据以上的设计原则本试验选取了两个酶切位点分别是XbaI和SpeI酶切位点,正向双酶切引物为5′-GCTCTAGAATGGATTTCCAACCAAACAC-3′;反向双酶切引物为5′-GCACTAGTAAGCCTTAAAGACAAATCTA-3′。
2)目的基因PCR扩增及胶回收。酶切引物合成后,以连接PEASY-T1-PuZFP103阳性质粒为模板进行PCR扩增。使用2×EasyTaqPCR SuperMix 克隆试剂盒进行PCR扩增。反应程序:94 ℃预变性2 min;98 ℃变性10 s,58 ℃退火30 s,68 ℃延伸30 s,共35个循环;68 ℃延伸7 min。在120 V电压下,电泳检测目的条带是否正确,若条带正确则使用快速琼脂糖凝胶DNA回收试剂盒对其进行胶回收。
3)pBI121-GFP载体质粒的提取。在无菌的150 mL小瓶中加入20 mL含50 mg/mL卡那霉素的LB液体培养基,再加入15 μL含pBI121-GFP质粒的大肠杆菌菌液,放置于37 ℃摇床(购自太仓华利达,HZ9210K)上160 r/min震荡培养12 h,使用购自OMEGA公司(www.omegabiotek.com.cn)的质粒小提试剂盒提取pBI121-GFP载体质粒。
4)目的基因与载体的双酶切反应,纯化及连接。将所要的胶回收产物和pBI121-GFP质粒同时在37 ℃恒温培养箱(购自太仓华利达,HZ9210K)内进行XbaI和SpeI双酶切,内切酶购自北京全式金生物技术有限公司(www.transgen.com.cn),酶切体系如下: pBI121-GFP载体质粒100 μL、PuZFP103胶回收产物(小于2 000 ng)、10×FlyCutBuffer 20 μL、XbaI 2 μL、SpeI 2 μL、ddH2O;共200 μL体系。双酶切反应条件为37 ℃,酶切反应时间为3 h。最后使用核酸纯化试剂盒购自OMEGA公司(www.omegabiotek.com.cn)进行纯化。使用T4连接酶北京全式金生物技术有限公司(www.transgen.com.cn)将目的基因与载体在16 ℃恒温培养箱过夜连接,连接体系为10 μL体系:PuZFP103基因XμL,pBI121-GFP载体(10-X) μL,5×T4 DNA Ligase Buffer 2 μL、T4 DNA Ligase 1 μL、ddH2O;X满足公式3×C1×X=C2×(10-X),其中C1为pBI121-GFP质粒纯化后浓度,C2为目的片段纯化后的浓度。
5)大肠杆菌的转化及PCR检测。取过夜连接的连接产物通过热激转化法,转化至大肠杆菌DH5α中(菌株来自本实验室保存),吸取500 μL无抗生素LB液体培养基于灭菌的1.5 mL的无菌离心管中,并置于37 ℃恒温摇床震荡培养1 h,1 h后将菌液置于离心机(德国Eppendorf超高速冷冻离心机)5 000 r/min,离心5 min,离心后弃去上清液,留取大约120 μL上清进行菌体重悬,然后将菌液涂到含有50 mg/L卡那霉素的LB固体培养基平板中,将烧过的无菌涂棒放凉后将菌液涂至平板。在37 ℃摇床中倒置过夜培养后,用无菌的镊子夹取无菌的小枪头挑取单克隆菌落至无抗液体LB中摇菌3 h,然后吸取菌液进行菌液PCR及电泳检测,并抽取500 μL阳性菌液送去公司进行测序,将剩余菌液中加入50%甘油按体积比1∶1充分混匀后放入液氮迅速冷冻后置于-80 ℃冰箱保存。测序比对正确后的菌液使用高纯度质粒小提试剂盒提取PuZFP103-pBI121-GFP质粒。
1.2.4PuZFP103基因生物信息学分析
利用ExPASy服务器中的Protparam(https://web.expasy.org/protparam/)工具分析蛋白的理化性质;通过软件 Predictprotein (https://www.predictprotein.org/)预测C2H2编码蛋白质的二级结构;利用网站 Phyre 2 (http://www.sbg.bio.ic.ac.uk/phyre2)对PuZFP103基因编码的蛋白质进行三级结构预测;根据ExPASy服务器中的ProtScale程序分析大青杨PubZFP103蛋白的疏水性;通过TMHMM Server v.2.0对蛋白进行跨膜区分析;使用SignalP 3.0 server (http://www.cbs.dtu.dk/services/SignalP-3.0/)对蛋白进行信号肽预测;使用COILS Server(https://embnet.vital-it.ch/software/COILS_form.html)对蛋白卷曲螺旋预测分析;通过http://www.bio-soft.net/sms/index.html查询该基因的同源核苷酸及氨基酸序列;NCBI在线预测蛋白结构域;通过http://multalin.toulouse.inra.fr/multalin/multalin.html进行氨基酸多序列比对;通过http://multalin.toulouse.inra.fr/multalin/cgi-bin/multalin.pl网址进行氨基酸结构比对;使用 BLAST软件对氨基酸进行同源性分析,以ZFP蛋白的氨基酸序列为基础,在NCBI数据库中检索到具有较高大青杨ZFP同源性的其他物种的氨基酸序列进行比较,然后使用MEGA 5.05软件通过邻接法构建系统进化树。
1.2.5 亚细胞定位
提取上述构建好的瞬时表达载体pBI121-PuZFP103-GFP及对照载体pBI121-GFP质粒。将提取的质粒包裹金粉制备成pBI121-PuZFP103-GFP的DNA微弹及对照载体pBI121-GFP的DNA微弹。取新鲜洋葱的倒数第2~4层鳞茎内表皮,用干净的手术刀切下面积为2 cm2的洋葱块,小心撕取其内表皮,平铺到MS培养基上,然后用基因枪(PDS-1000/He Particle Delivery)利用氮气对洋葱受体进行轰击。将轰击后的洋葱受体密封置于常温暗处培养24 h,然后使用激光共聚焦微镜(OlympusFV1000MPE)观察亚细胞定位情况。
1.2.6 非生物胁迫处理取材
选取在无菌的1/2 MS培养基中生长3周,生长良好且状态一致的大青杨幼苗为研究对象,将其连根整株插入到添加200 mmol/L NaCl、200 μmol/L ABA及质量分数10%的聚乙二醇PEG6000的1/2 MS培养基条件下分别进行胁迫处理3、6、12、24、48、72 h。处理后用液氮速冻材料并于-80 ℃冰箱保存备用。
1.2.7 RT-qPCR分析基因的表达模式
利用CTAB法分别提取胁迫处理前后的大青杨叶和根部位的总RNA,使用南京诺唯赞生物生产的HiScript©Ⅱ Q RT SuperMix for qPCR (+gDNA wiper) 反转录酶将其反转录成cDNA作为模版,放置在-20 ℃冰箱内保存备用。每个处理组均有0 h空白对照,F2R2为荧光定量引物F2:(5′-3′)ATCCTCTTCTTCTTCATCATC,R2;(5′-3′)GACGATCCTGACCGTGGG);PtrActin-F PtrActin-R为内参引物[PtrActin-F(5′-3′):TGTTGCCCTTGACTATGAGCAGGA ;PtrActin-R(5′-3′):CGGAATCTCTCAGCTCCAATGGT]。通过荧光定量PCR仪(Applied Biosystems, Foster City, CA, USA)使用南京诺唯赞生物公司生产的ChamQTM Universal SYBR© qPCR Master Mix进行RT-qPCR分析,每个样品3次生物学重复,用 2-ΔΔCt计算法计算其相对表达量。RT-qPCR总反应体系为20 μL: cDNA模版1 μL、正反向引物各1 μL、2×EasyTaqPCR SuperMix酶10 μL、去离子水7 μL。反应程序:94 ℃预变性30 s;94 ℃变性5 s,60 ℃,30 s,共45个循环;55~99 ℃每隔0.5 ℃读板1次,持续1 s。
2 结果与分析
2.1 大青杨PuZFP103基因的克隆
以生长3 周大的无菌大青杨为材料分别提取根和叶的总RNA。利用总RNA反转录成cDNA,以2 000 marker为对照,扩增该基因的CDS序列,扩增产物长度为504 bp(图1),片段大小与预期符合。
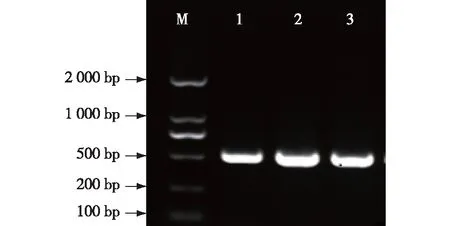
M.DNA marker DL 2000;1~3.PuZFP103克隆产物 PCR production of PuZFP103。图1 PuZFP103基因PCR扩增结果Fig.1 The PCR amplication results of PuZFP103
2.2 大青杨PuZFP103基因生物信息学分析
2.2.1 大青杨PuZFP103基因的DNA、蛋白序列分析及蛋白结构域预测
通过在线查询得知大青杨PuZFP103基因共编码168 个氨基酸(图2A)。利用ExPASy服务器中的Protparam工具分析蛋白的理化性质,分析结果氨基酸的数量为168,分子质量为18 673.64 u,理论等电子点为8.53,总原子数2 559,分子式为C800H1247N249O256S7。在组成蛋白的20种氨基酸中,丝氨酸(Ser)所占比例最高,占18.50%;色氨酸(Trp)所占比例最低,占0.60%,不包括含硒半胱氨酸(Sec)和吡咯赖氨酸(Pyl)。蛋白的不稳定指数为69.35,不稳定蛋白脂肪指数为59.23,总平均亲水性为-0.531。正电荷残基数 (天冬氨酸Asp+谷氨酸Glu)为16。负电荷残基数 (精氨酸Arg+赖氨酸Lys)为18。此蛋白的亲和性的平均水平 (GRAVY)为-0.756。利用Smart软件对序列进行分析(图2B),发现PuZFP103具有1个ZnF-C2H2结构域,位于该氨基酸序列的43~65位,隶属于C2H2家族。
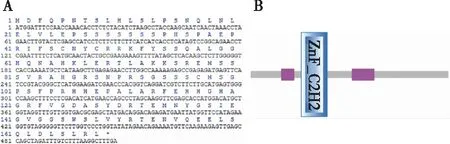
图2 PuZFP103基因序列及其推导的氨基酸序列 (A)和PuZFP103蛋白的结构域预测(B)Fig.2 Nucleotide and deduced polypeptide sequences of PuZFP103 (A)and conserverd domain prediction of PuZFP103 coded protein(B)
2.2.2 编码蛋白的二三级结构预测分析
通过软件 Predictprotein (https://www.predictprotein.org/)预测C2H2编码蛋白质的二级结构。经网站分析该蛋白包含3种结构分别为螺旋(helix)、折叠(strand)和无规则卷曲(loop),分别占总氨基酸的 8.33%、5.95%和 85.71%(图 3A)。利用网站 Phyre 2 (http://www.sbg.bio.ic.ac.uk/phyre2)对PuZFP103基因编码的蛋白质进行三级结构预测,如图(图3B)所示PuZFP103基因编码的蛋白质三级结构与预测的二级结构相符,N端区域主要由α-螺旋和β-折叠组成,C端区域主要由无规则卷曲组成。
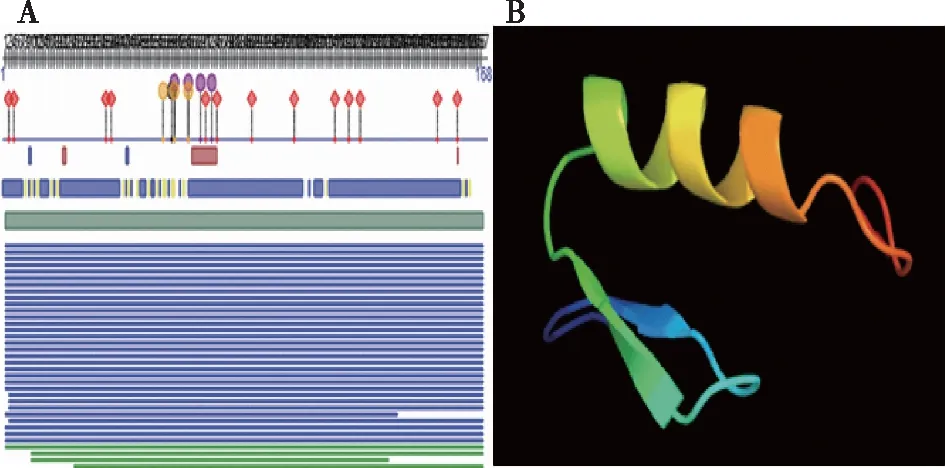
图3 PuZFP103蛋白质二级结构(A)和 三级结构(B)预测Fig.3 Protein secondary stucture (A) and tertiary structure (B)prediction of PuZFP103 protein
2.3 大青杨PuZFP103基因的理化特性
2.3.1 大青杨PuZFP103基因编码的蛋白疏水性
大青杨PuZFP103基因编码的蛋白疏水性根据ExPASy服务器中的ProtScale程序分析,计算基于K-D法的蛋白质疏水性。分析结果发现,第145 位的氨基酸为丝氨酸,疏水性最大,分值为0.944;第87位的氨基酸为精氨酸,亲水性最大,分值为-2.356。根据图4中数据可以看出,负分数的氨基酸数大于正分数的氨基酸数,这说明该蛋白的亲水性大于疏水性,推测该蛋白质为亲水性蛋白质。
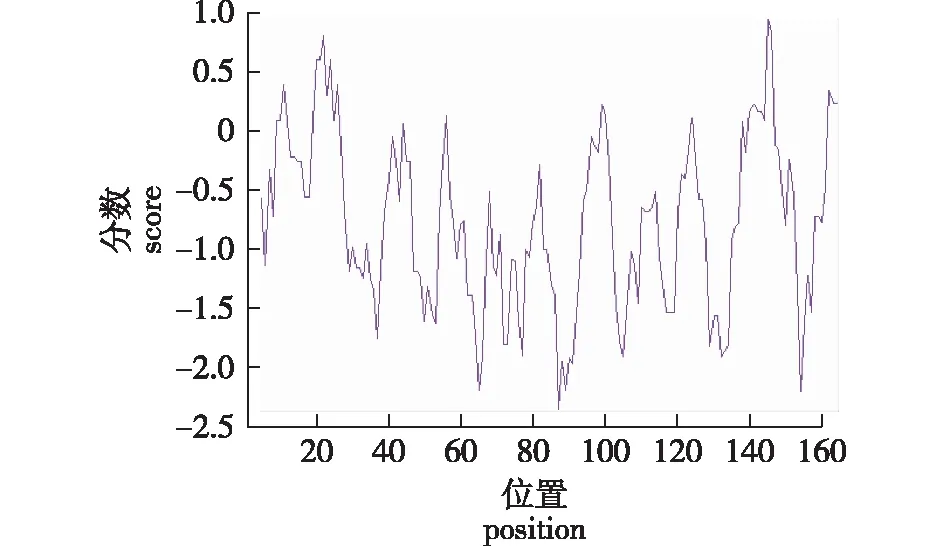
图4 PuZFP103蛋白的疏水性预测Fig.4 Hydrophobicity analysis prediction PuZFP103protein
2.3.2 蛋白跨膜区分析
通过TMHMM Server v.2.0对蛋白进行跨膜区分析 (图5)。发现该蛋白不存在跨膜区结构,并没有典型的跨膜螺旋区,也没有氨基酸位于细胞膜内,参考该蛋白的疏水性区域分析结果,表明该蛋白并不是一个与细胞信号传导有关的膜受体蛋白。
图5 PuZFP103蛋白质的跨膜结构Fig.5 Protein transmembrane structure diagrams ofPuZFP103 protein
2.3.3 蛋白信号肽及蛋白卷曲螺旋预测分析
使用SignalP 3.0 server (http://www.cbs.dtu.dk/services/SignalP-3.0/)对蛋白进行信号肽预测,预测蛋白质N端没有信号肽,预测为Non-secretory蛋白质。该蛋白不存在信号肽序列,可以高效表达。使用COILS Server对蛋白卷曲螺旋预测分析,结果 (图6)表明,3条曲线分别代表窗口系数(window width)为14、21、28时预测卷曲螺旋的位置。该软件预测出1个卷曲螺旋。因为卷曲螺旋一般由7个氨基酸构成,所以该选项都是7 的倍数。
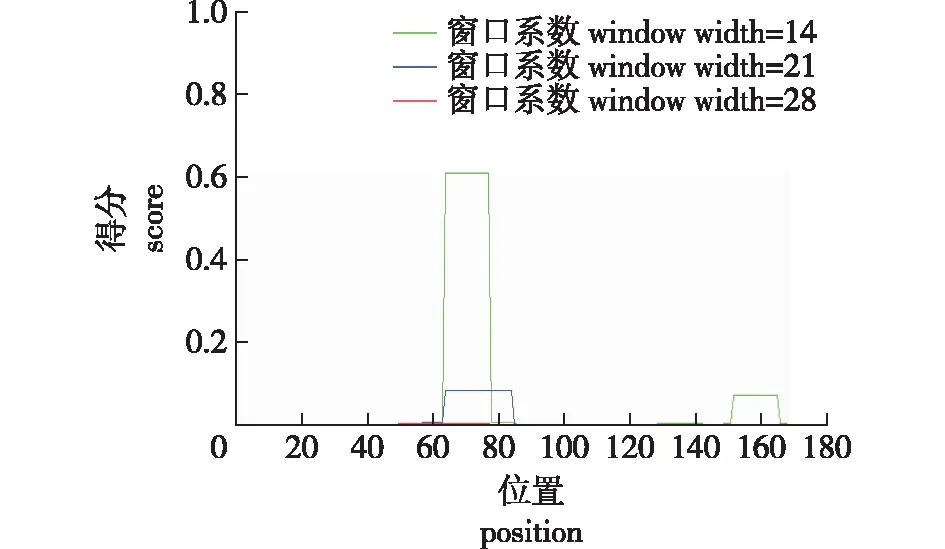
图6 PuZFP103蛋白的卷曲螺旋预测Fig.6 Coiled coils prediction of PuZFP103 protein
2.3.4 氨基酸序列比对及系统进化树构建
通过在线比对氨基酸序列,将大青杨PuZFP103基因编码的氨基酸序列与其他植物同源基因的氨基酸序列进行同源性比对(图7),结果显示,大青杨PuZFP103与毛果杨、胡杨(P.euphratica)、毛白杨(P.tomentosa)和番木瓜(Caricapapaya)有较近的遗传距离,所以预测他们有更近的亲缘关系。而与可可(Theobromacacao)、锦葵(Malvacathagensis)、榴莲(Duriozibethinus)遗传距离较远,因此预测他们亲缘关系较远。
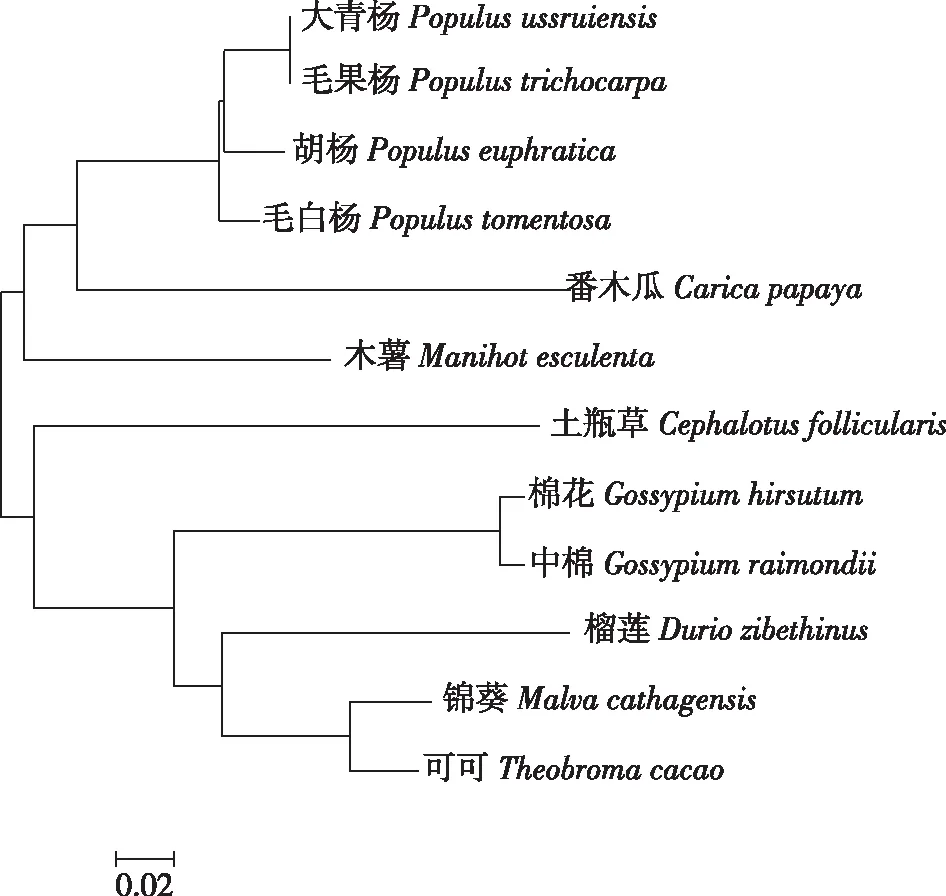
图7 大青杨PuZFP103基因与其他物种直系同源蛋白系统进化树分析Fig.7 The phylogenetic tree analysis of PuZFP103 gene in P. ussuriensis and homologous proteins inother species
2.4 大青杨PuZFP103基因的亚细胞定位
利用基因枪分别将金粉包裹的pBI121-PuZFP103-GFPDNA微弹质粒和对照pBI121-GFPDNA微弹质粒通过基因枪轰击洋葱内表皮,使用激光共聚焦显微镜观察(图8)。从图8中可以看到对照组pBI121-GFP空载体在整个洋葱细胞内均能发绿色荧光,为组成型表达;pBI121-PuZFP103-GFP的融合表达载体在35S启动下PuZFP103-GFP只在细胞核中有荧光,这进一步证明PuZFP103定位在细胞核中,具有转录因子的一般特征。
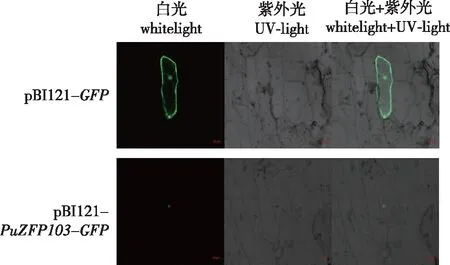
图8 PuZFP103基因的亚细胞定位Fig.8 The subcellular localization analysis of the PuZFP103 gene
2.5 大青杨PuZFP103基因的表达模式分析
观察在非生物胁迫下其不同时间和不同部位的PuZFP103基因表达量情况。在 NaCl、ABA和PEG 6000胁迫条件下,基因PuZFP103的表达量在根和叶中都较为明显,结果如图9所示。NaCl胁迫后PuZFP103基因在大青杨叶片中的表达量逐渐升高,胁迫6 h时的表达量最高,之后逐渐下降,在48 h又升高。而在大青杨根中表达量呈明显的上升趋势,胁迫3 h和24 h的表达量都很高(图9A),所以预测其可能在根中发挥作用。ABA胁迫后,PuZFP103基因在大青杨叶中的表达量在6 h之前是下降的,在12 h后逐渐升高,72 h时又降低,而在大青杨根中表达量只有在6 h时表达量最高,在其他时间点都是下调(图9B),因此预测ABA可能发挥负调控作用,有待进一步研究确定该结果。PEG 6000胁迫后PuZFP103基因在大青杨的叶中的表达量在3 h之前的表达量是微微下调的,在6 h后逐渐升高,在24 h时表达量最高,而在根中表达量都比0 h高,在6 h时表达量升高近4.5 倍(图9C)。
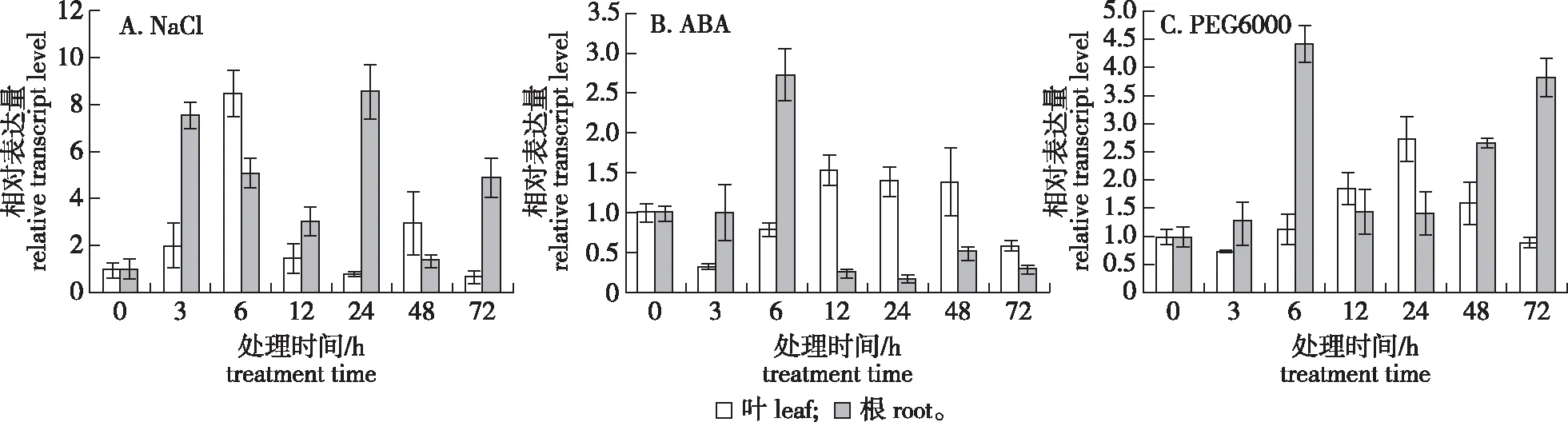
图9 大青杨PuZFP103基因在NaCl ABA和PEG 6000处理下的表达量的分析Fig.9 The RT-qPCR analysis of the expression pattern of PuZFP103 gene in P. ussuriensisunder treatment of NaCl,ABA and PEG 6000
3 讨 论
C2H2型锌指蛋白是最常见的锌指蛋白之一,在真核生物的生长发育及应对胁迫的响应中起重要作用[23]。近年来,国内外对植物C2H2型锌指蛋白的研究明显增多,发现了多种与植物生长发育和抗逆性相关的C2H2锌指蛋白[24]。笔者从大青杨中克隆出PuZFP103基因,利用生物信息软件对其进行基因组分析以及基因功能的研究。分析结果表明,该基因包括有504 bp的CDS序列,共编码168 个氨基酸。通过对PuZFP103氨基酸序列进行蛋白质结构预测,发现该蛋白包括C2H2锌指蛋白结构域,预测该蛋白为亲水性蛋白。对蛋白进行信号肽预测,预测蛋白质N端没有信号肽,为Non-secretory蛋白质,该蛋白不存在信号肽序列,可以高效表达。同时对蛋白卷曲螺旋预测分析,检测到1个卷曲螺旋。预测C2H2编码蛋白质的二级结构,结果显示螺旋结构(helix)、折叠(strand)和无规则卷曲(loop)分别占总氨基酸的 8.33%、 5.95%和85.71%。此外,基于系统进化树分析结果,推测PuZFP103基因可能参与大青杨生长发育与逆境响应过程。因为有研究表明,与生长发育或胁迫相关的ZFP蛋白物种在进化树分类中能够与大青杨聚为一个亚类[25]。通过亚细胞定位分析,证明该基因位于细胞核中。这些生物信息学分析结果将为深入探究PuZFP103基因在大青杨中的功能奠定一定的理论基础。
本研究对生长状态一致的大青杨分别进行NaCl、PEG 6000及ABA非生物胁迫处理,探究其在不同组织和不同非生物胁迫下的表达特性。在激素与非生物胁迫应答分析中, NaCl和PEG处理后,PuZFP103在叶片和根表达量主要呈现上调趋势,说明该基因对NaCl和PEG 6000信号途径主要呈现正响应模式。通过ABA处理后,PuZFP103的根在6 h时基因表达量上调,其他时间点都下调的趋势,在叶片中的表达量变化也不明显,这说明PuZFP103基因对ABA产生了应答。可以推测,PuZFP103基因可能在大青杨适应这3 种非生物胁迫因子过程中起重要作用。表明PuZFP103基因可能参与了大青杨对渗透胁迫的信号传导,属于逆境胁迫下的调控因子。大量研究表明,ZFP蛋白参与植物对逆境胁迫的响应过程[26]。甘薯中ZFP1经ABA、PEG 6000和NaCl处理后表达量提高[27]。大豆(Glycinemax)中GsZFP1的表达在盐、冷胁迫和ABA胁迫下在叶片中被诱导表达,在干旱、ABA和冷胁迫下在根部被诱导表达[28]。研究证明,马铃薯(Solanumtuberosum)的StZFP1可能通过ABA依赖途径响应盐和干旱的胁迫,在拟南芥中 C2H2 型锌指蛋白ZFP6通过整合植物激素信号从而调节毛状体的初始化[29]。
笔者从大青杨中克隆出PuZFP103基因,并通过生物信息学、不同胁迫下的组织特异性及表达模式等分析预测基因结构和功能。生物信息学和RT-qPCR分析结果初步证明PuZFP103基因可能在大青杨响应胁迫过程中扮演重要角色,明确了其保守结构域为ZnF C2H2,进而明确了PuZFP103基因属于C2H2家族亚类基因,在激素与非生物胁迫应答分析中,ZFP103基因均呈现出表达量的变化,说明该基因可能参与了植物激素的应答与胁迫响应的过程。具有一定的抗旱和耐盐等能力,但是关于其信号传导与调控机制的研究需进一步的研究。本研究可为大青杨ZFP103基因的功能研究及抗逆机制的深入研究提供理论基础。
