双价RNA干扰载体转基因水稻抗飞虱研究
2021-01-20王爱娜
杨 颖,王爱娜
(渭南职业技术学院,陕西 渭南 714026)
通过导入Bt基因获得转基因抗虫植株,已在棉花、番茄、油菜、玉米等作物上获得成功,并且有的通过商业化释放,在农业生产方面产生良好的经济和生态效益。目前,从微生物、植物和动物来源的抗性基因已被成功用于转基因研究的候选基因,在转基因作物抗虫研究应用方面,不同类型的苏云金芽孢杆菌内毒素编码基因Cry已被广泛用于不同作物对鳞翅目、双翅目、鞘翅目或者膜翅目的昆虫的抗性[1]。然而,昆虫对杀虫毒素迅速进化产生的抗性,使毒素对害虫的无效性,对非靶向生物和微环境的影响是现有转基因抗虫方法的主要缺陷[2]。考虑到目前作物保护现状,有必要探索出一种更加经济、环境友好型的害虫防治方法,因此利用植物介导的RNAi方法来抵抗褐飞虱有着潜在的价值[3]。
为利用RNAi技术培育抗褐飞虱水稻,我们先前采用重组PCR将已克隆获得的褐飞虱V-ATPase D亚基基因片段和V-ATPase H亚基基因片段进行融合,融合基因V-ATPase D-H (VAD-H)以反向重复的方式连入改造后的pUCm-T,通过体外转录dsRNA饲喂褐飞虱若虫致其死亡率增加[4],为进一步验证体外稳定表达dsRNA的转基因植株在抗飞虱方面的作用,本研究通过将hp(VAD-H)定向插入到携带有筛选标记抗草丁膦bar基因的双元表达载体pCAMBIA-1300上,从而产生发夹样pcambia1300-Ubi-hp(VAD-H)-RNAi植物表达载体,经转化获得转基因植物,飞虱取食转基因植株后对其生长发育进行测定。
1 材料与方法
1.1 试验材料
1.1.1 材料 试验所用褐飞虱由本校植物抗逆研究中心提供,在光照培养箱内(温度25±2℃,相对湿度60%~70%,15 h光照/9 h黑暗)利用水稻幼苗进行饲养繁育。
1.1.2 主要试剂 本试验所用大肠杆菌(E.coli)菌株,已构建载体pUCm-hp(VAD-H)来自本实验室。引物由擎科生物合成,植物RNA提取试剂盒购自康为世纪,PrimeSTAR HS DNA Polymerase、内切酶和T4 ligase购自Takara公司,cDNA合成和qPCR试剂盒均购自全式金公司。
1.2 试验方法
1.2.1 植物双元载体pCAMBIA1300-hp(VAD-H)的构建及水稻转化 已获取的含有发夹结构的重组载体pUCm-hp(VAD-H),利用pUCm-T载体上带有的酶切位点HindⅢ和Sac I双酶切pUCm-hp(VAD-H),将发夹结构插入经改造携带有筛选标记抗草丁膦bar基因的双元表达载体pCAMBIA-1300,从而产生发夹样pcambia1300-Ubi-hp(VAD-H)-RNAi植物表达载体,该载体中目的干扰基因由Ubi启动子驱动,选择标记bar基因由组成型35s启动子驱动。载体经农杆菌介导转化TN1水稻植株。转化植株采用草丁膦进行筛选,筛选植株在温室内进行培养。
1.2.2 转基因水稻饲喂褐飞虱 通过喷施除草剂筛选抗性植株到T3代水稻幼苗进行生测,选取长势一致的4叶期阳性转化植株及非转化植株(对照)转移到生测装置内,每个水培杯内种一株,植株根上部用塑料圆盘固定,并罩有一端扎有密孔尼龙网的透明塑料圆柱体,每个植株上放置10只12h内孵化的初孵1龄若虫,每天记录飞虱的数量和龄期,持续12 d,该试验重复10次,试验结束后收集飞虱,放置-80℃再进行后续实验。生测实验在人工气候室内进行,温度25±2℃,相对湿度60%~70%,15 h光照/9 h黑暗。
1.2.3 实时定量RT-PCR检测 用康为世纪的植物RNA提取试剂盒(DNase I)提取总RNA,经反转录试剂盒合成cDNA。cDNA稀释后作为模板,进行qRT-PCR,所用引物如表1所示。反应体系为20μL:2×TransStart Tip Green qPCR SuperMix 10 μL,Forward Primer(10 μmol·L-1)0.4 μL,Reverse Primer(10 μmol·L-1)0.4 μL,cDNA模板(20 ng·μL-1)1μL,ddH2O 8.2 μL。qPCR反应程序为:94℃ 30 s;94℃5 s,60℃30 s,40个循环。ABI Prism 7300为实时定量仪器,飞虱β-actin基因(GenBank:EUl79846)作为内参,基因相对表迖量计算采用Livak和Schmittgen[5]的方法。
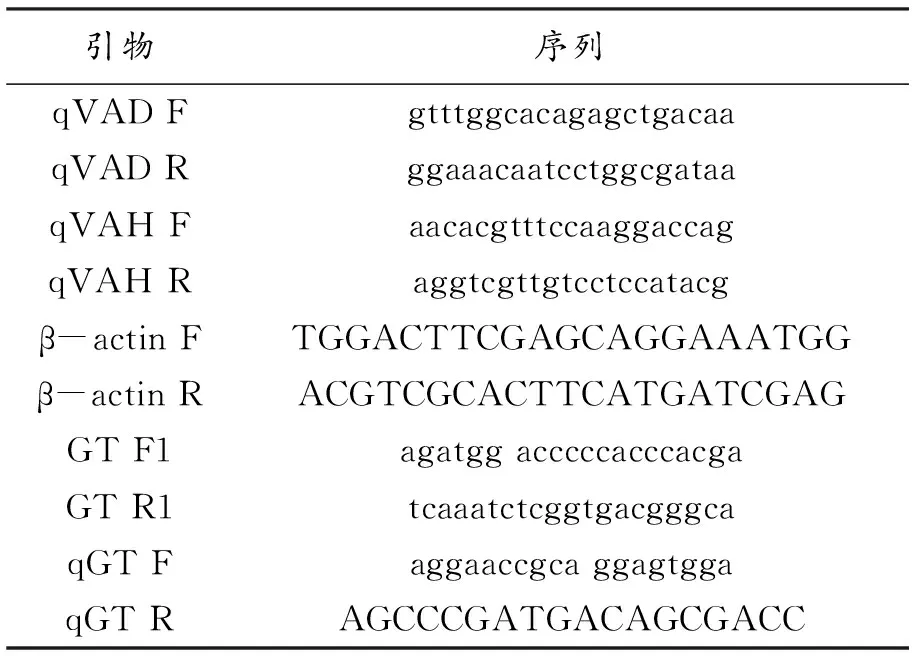
表1 引物序列
2 结果与分析
2.1 dsRNA转基因水稻获取及鉴定
将构建的含有筛选标记抗草丁膦bar基因的dsRNA干扰表达载体1300-Ubi-hp(VAD-H)-RNAi,通过农杆菌介导转化TN1水稻愈伤组织,采用草丁膦进行筛选,进而获取转基因水稻植株。在温室内培养,T0代转化植株收获的种子(T1)播种待幼苗长至三叶期时喷施草丁膦,选取除草剂抗性植株(T1)收获种子(T2),继续种植筛选抗性植株到T3代,单株种植选取抗性不分离株系即为纯合体,从抗草丁膦的阳性转化植株和对照TN1植株中分别提取基因组DNA,用特异引物GTF1和GTR1扩增除草剂基因进行PCR检测,结果如图2a中可见所选取植株均为阳性转化植株,随后对植株进行除草剂基因的表达量检测,如图2b我们选取A3和A6两个植株进行后续试验。
2.2 体外喂食转基因水稻后对飞虱影响
2.2.1 取食转基因水稻对飞虱生长发育影响 为探究体外稳定表达dsRNA的转基因植株对飞虱发育水平的影响,我们选取T3代植株A3和A6,待其长到四叶期幼苗用以喂食褐飞虱。体外饲喂试验所用褐飞虱为TN1水稻上的初孵幼虫,随后将飞虱接种到转基因水稻A3和A6 植株上,褐飞虱取食VAD-H转基因水稻后,如图3所示该转基因水稻对褐飞虱1龄若虫的发育历期影响明显,差异显著外,对其他龄期若虫发育历期影响无明显变化(图3)。同时褐飞虱若虫总发育历期,相比对照明显延长。
我们对飞虱成虫存活寿命,世代周期同时进行了测定。取食转基因水稻后,短翅型雌虫寿命受到影响,与对照组相比较差异显著,而对于短翅和长翅型雄性成虫的寿命无显著影响,此外,飞虱的世代周期相较于对照也明显延长(图4)。
图2 转化植株的检测
(a)T3代转化植株PCR检测。M,DL2000 marker;1,ck;2-7,不同转化植株;
(b)T3代转化植株的qRT-PCR。A1-A6,不同转化植株。
2.2.2 取食转基因水稻对飞虱靶基因影响 为检测飞虱取食转基因植株后对其体内靶标mRNA的影响,利用荧光定量PCR对靶标基因的转录水平进行测定,试验所用的3龄褐飞虱起初在TN1水稻上进行饲喂,随后转移到转基因水稻A3及A6上持续饲喂4~8 d。对饲喂非转基因TN1和转基因水稻幼苗的褐飞虱分别抽提RNA,反转录后检测褐飞虱体内靶基因的相对表达量。
褐飞虱取食VAD-H转基因水稻,靶基因VATPase H的转录水平开始降低,A3和A6植株饲喂的飞虱其靶基因转录水平在第八天分别降低20.4%和28.6%(图5A)。
同时靶基因V-ATPase D的转录水平也显著降低,A3和A6植株上飞虱该基因转录水平在第八天分别降低了40.3%和43.5%(图5B)。
3 讨论
获得在植物体内稳定表达昆虫基因dsRNA的转基因植物,成功介导昆虫体内RNAi效应,使其靶基因被沉默甚至正常生理活动受到一定影响,对于减轻农作物虫害具有潜在的应用价值。Baum等利用表达玉米根叶甲VATPase基因dsRNA的转基因玉米,饲喂玉米根叶甲幼虫,幼虫出现发育受阻或死亡现象[6]。作为影响水稻生产的主要害虫褐飞虱,其通过刺吸式口器,吸食植株维管束韧皮部汁液内的自由氨基酸和氨基类化合物,对水稻造成严重的生理损害。Zha等[7]选取了在褐飞虱中肠内高表达并且参与糖转运的三个基因即NlHT1、Nlcar和Nltry,构建并转化获得表达上述基因dsRNA的转基因植株,褐飞虱取食转基因水稻后其体内对应基因的表达量下降了40%~70%。
我们先前克隆了飞虱V-ATPase D和V-ATPase H亚基基因片段,利用重组PCR获得融合基因VAD-H,通过体外转录dsRNA饲喂褐飞虱并引起飞虱死亡率增加,在此基础上构建了针对该融合基因的RNAi植物表达载体,获取体外稳定表达dsRNA的纯合株系,进一步检测转基因植物对飞虱生理水平的影响。当褐飞虱的初孵幼虫接种到转基因植株上,取食VAD-H转基因水稻后,其1龄若虫的发育历期有所延长,而且飞虱若虫总发育历期,相比对照也明显延长(图3)。此外我们对取食dsRNA的褐飞虱靶基因转录水平,进行荧光定量PCR,结果表明飞虱体内靶基因表达水平均发生不同程度降低,并且降低水平随饲喂时间延长显著性下降(图5)。表明摄入表达融合基因dsRNA的转基因植物,能够引发褐飞虱体内RNAi效应。
Li等研究在转基因水稻表达产生高浓度dsRNA时才能引发高效率的环境性RNAi,并且可以阻碍褐飞虱的生长发育甚至能够导致褐飞虱死亡[8],因此当外界不能持续提供dsRNA时,干扰效果会受到限制。笔者研究中飞虱取食转基因水稻后尽管生长发育相比对照受到一定的影响,但没有出现明显的致死表型,出现上述问题可能由于飞虱摄取dsRNA的量不足,或实验体系不能引发RNAi强穿透能力,使得飞虱未出现死亡。因此在利用RNAi技术培育转基因抗虫植物时,需注意包括基因类型,基因片段大小,表达载体选择等影响干扰效率的因素,从而充分发挥其干扰效应在害虫防治方面的作用[7]。
