碱胁迫下不同水稻品种微观结构响应解析
2020-12-17赵海新黄晓群陈书强杨丽敏杜晓东张志强蔡永盛潘国君
赵海新,黄晓群,陈书强,杨丽敏,杜晓东,张志强,蔡永盛,潘国君
(黑龙江省农业科学院水稻研究所,黑龙江 佳木斯 154026)
不同植物物种对逆境适应性存在差异,因其应对调解机制不同[1],产生竞争优势,即使这种调解机制差别微小,也可能造成物种分离与进化[2]。自然逆境主要包括生物(病虫草害等)和非生物类[3](地理状况、土壤类型、极端气候等),盐碱化土壤是植物生长非生物类逆境。物种、品种及基因家族[4]对这种逆境压力表现适应性差异[5],这种差异形成源于基因与蛋白质组[6],表现于植物生长状况和微观结构变化[7-8]。认识植物对盐碱胁迫反应,是判断植物耐盐碱性重要基础。植物微观结构对盐碱胁迫响应的相关报道较多,研究植物种类也较广泛,认为植物叶绿体是亚细胞结构对盐分最敏感的细胞器[9-10],可造成膜系统紊乱解体[11],叶片PSⅡ反应中心遭到破坏[12]。高强度胁迫使叶绿体迅速老化膨胀,基粒片层结构扭曲解体[4],基质变稀,嗜锇颗粒空腔化[5,10],线粒体结构与数量亦有相应变化[13],甚至引发细胞程序性死亡[14]。
水稻分布地域广泛,覆盖地域土壤类型丰富,其中包含大量盐碱型土壤。水稻对盐碱环境具有适应性,但品种间存在差异[5,14]。研究主要集中于形态与生理机能方面[15]。通过一些微观领域研究也表明,品种间叶片细胞超微结构存在差异[5],是否与对盐碱环境适应性有关则未见报道。本研究利用碱性盐胁迫苗期水稻,解析根系细胞形态和不同品种叶片细胞及亚细胞结构响应机制,从微观角度解析水稻受碱胁迫微观表征和各细胞器响应机制。
1 材料与方法
1.1 试验材料
供试材料:龙粳27、绥粳5号、藤系138和越光。龙粳27和绥粳5号源自于中国;藤系138和越光源自于日本。
1.2 试验地点与气候条件
试验地位于黑龙江省农业科学院水稻研究所,地理位置N46°49′、E130°22,属于温带大陆性季风中温带湿润气候区,年均气温3℃,有效活动积温2 590℃,无霜期130 d,历年年均降水量530 mm。
1.3 试验设计
1.3.1 幼苗前期培养
种子经0.1%高锰酸钾浸泡消毒24 h后,撒播在盛有石英砂托盘中,播量为100 g·m-2,沙子厚度2.5 cm,其中覆沙0.5 cm,淋水后,放入恒温培养箱,温度30℃,72 h后出齐苗取出,在室温下生长至2叶。
1.3.2 幼苗后期培养
培养盒上口尺寸为30 cm×20 cm,侧面和底部用遮光纸封闭,其上覆盖厚度1 cm泡沫载板,载板均匀打穿20个直径1.0 cm培养孔(相邻孔边距5 cm×5 cm),每孔选取一株2叶期幼苗,用薄海绵条包卷主茎后,插入培养孔,使其固定在培养板上,于木村培养液中室外培养,每2 d更换一次培养液。
1.3.3 处理药剂与时期
药剂使用含Na2CO3和NaHCO3成分混合碱,两者质量比为1∶3,在水溶液中质量浓度为0.20%。设对照(CK)和0.20%处理(CL),均为3次重复,室外处理。当幼苗第3叶龄叶片完全展开后,将培养液更换为含混合碱木村培养液(下称处理液,室温,pH 9.01),对照CK生长于木村培养液(室温,pH 6.0),每2 d更换一次处理液和培养液,10 d后,摘取顶端第一片完全展开叶片中段开展测定和观察。
1.4 指标测定与方法
维管束、中柱和根直径:取对照和处理各20条主根,电子显微镜下观察拍摄横截面,由电子显微镜图像处理软件获取数据。各指标比值乘以100%即获得各项指标间比值。
细胞长宽与宽度:取对照和处理各20条主根,电子显微镜下观察拍摄纵剖面,由电子显微镜图像处理软件获取数据,纵向为细胞长度,横向为宽度,通过计算获取各项指标间比值。
维管束数量:电子显微镜观察对照和处理各20条根,取平均值。
维管束、中柱和根横截面积:在标准纸上打印待测图像,剪取待测指标图形,称重。使用软件获得标准长度为边长,在同一张标准纸上绘制正方形,计算面积并称重,通过比重法计算即获得相关数据。各指标比值乘以100%即获得各项指标间比值。
光合色素:采用李合生方法测定叶绿素a、叶绿素b、类胡萝卜素含量[16]。
细胞、叶绿体、线粒体及淀粉体面积:数据获取同上述比重法,通过计算获取各项指标间比值。
1.5 观察目标及处理方法
1.5.1 根系细胞观察方法
利用德国LeicaDM4000B电子显微镜,主要观察冠根各部位横截面及纵剖面细胞排列情况。切片用甲基红溶液染色后,立即在目镜放大率为10×122,物镜放大率为×10倍下观察照相。
1.5.2 叶片超微结构处理方法
制作切片参照洪健等方法并加以改进[17]。取主茎第4叶中间部分切成1 cm小段,迅速放至2.5%戊二醛中前固定,抽气使材料下沉,于4℃固定2 d。pH 7.2磷酸缓冲液冲洗,再用2%四氧化饿固定1.5 h,pH 7.2磷酸缓冲液再次冲洗。使用乙醇逐级脱水,再用100%乙醇∶丙酮=1∶1、纯丙酮各置换1次。用纯丙酮:环氧树脂812包埋剂逐级浸透(24 h)和包埋,浸透后在恒温培养箱中,按40℃17 h,45℃24 h,60℃24 h聚合,ULTRACUTE型超薄切片机切片,醋酸双氧铀、柠檬酸铅在25℃下分别对样品染色15~20 min,双重蒸馏水冲洗干净后待检,JSM25610LV透射电镜下观察拍照。
1.6 数据处理与分析
使用Excel 2013、DPS 18.10和SPSS 24.0软件处理和分析数据。
2 结果与分析
2.1 碱胁迫对水稻根细胞的影响
根据观察结果,品种间根细胞形态变化趋势相同,本试验部分以龙粳27和越光为例。
2.1.1 水稻根横截面细胞
由图1根横截面细胞变化可知,对照根系各层位细胞排列整齐、清晰、紧凑,细胞形状规则,立体感强,具有活性(见图1-1,2),表皮覆有浓密根毛组织(见图1-H),皮层薄壁细胞由内向外,呈由小到大辐射状排列。经碱胁迫10 d后,皮层薄壁细胞排列相比对照明显松散凌乱,染色后颜色分布不均匀,表皮根毛组织短小且呈芽状(见图1-3)。处理后根系维管柱初生大维管束组织数量相比对照减少1~3条,具有未完全发育的大维管束(见图1-6-m);对照内皮层和中柱鞘细胞排列紧密,细胞形态一致,处理组排列松散,脊状区域明显,已木质化。对照枝根数量较少,但经碱胁迫后枝根较多。胁迫明显打破水稻根细胞排列整齐性和紧凑性,抑制大维管束形成,未发育完全大维管束增加,抑制根毛生长,促进根系木质化和枝根发育。已处理水稻主根横截面具有较多褐色区域(见图1-3-n),其颜色改变程度与渗出细胞液有关,说明这些区域细胞生理功能已发生改变。碱胁迫处理后,龙粳27和越光变化趋势一致,两个水稻品种根系维管束数量、维管束与中柱直径比值极显著下降,中柱与根直径比、维管束与根直径比显著上升(见表1)。由面积比数据可知(见表2),中柱内维管束面积因胁迫,比例极显著降低,根系内中柱占横截面比和维管束在根横截面积中比显著和极显著上升。胁迫后维管束数量降低,但其直径和面积占根系比却显著增加,说明处理后单条维管束直径相对增加,且细胞内维管束面积之和相对增加。表明水稻为适应碱性环境,通过提高维管柱和维管束在根横截面中占比,保持物质运输畅通,以此提高抗逆性。
2.1.2 水稻根纵剖面细胞
对根纵剖面观察(见图2)表明,对照细胞排列整齐有序,呈圆柱状,皮层薄壁细胞立体感强,具有活性。处理后各层位细胞明显缩短,立体感下降,中部薄壁细胞呈多角形,靠近外皮层薄壁细胞呈较小的长方体,且部分细胞纵向长度小于宽度,靠近内皮层薄壁细胞呈圆形或短柱状(见图2-4,7),摆列有序性相比对照降低,且表皮细胞脱落(见图2-5-o),中柱鞘细胞明显变短细。与横截面细胞排列不同,纵剖面细胞排列相比横截面紧密。处理后水稻皮层薄壁和中柱鞘细胞均有深色斑点(见图2-7,靠近内皮层薄壁细胞部分区域颜色明显变深。

表1 碱胁迫对维管束、中柱与根直径的影响Table 1 Effects of alkali stress on the diameter of vascular bundle,central column and root
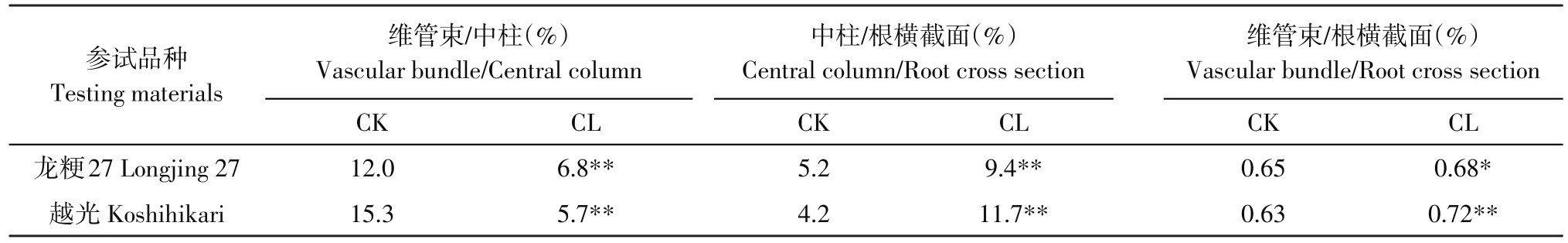
表2 碱胁迫对维管束与中柱占根系横截面积比的影响Table 2 Effect of alkali stress on the ratio of vascular bundle and central column to root cross-sectional area

图2 根纵剖面细胞结构Fig.2 Cell struture of root radicle cross section
由图2可知,碱胁迫明显抑制水稻根系细胞纵向伸长生长,造成根系生长缓慢,细胞立体感因胁迫而降低,说明根系已木质化。皮层薄壁细胞、中柱鞘部分细胞因胁迫死亡。靠近内皮层薄壁细胞受害较重,部分细胞群生理已发生变化。表皮细胞相比对照脱落较早,纵剖面图亦发现正在形成枝根横截面,对照较少,表明碱胁迫促进水稻枝根形成。
由表3可知,处理后供试材料薄壁和中柱细胞长宽比极显著降低,表明碱胁迫抑制根细胞伸长生长,且达极显著。龙粳27细胞长度受抑制程度相比越光更显著。中柱细胞长与薄壁细胞长比表明,品种之间出现反向变化趋势。相对中柱细胞,碱胁迫对龙粳27薄壁细胞长度抑制作用更显著,对越光根系薄壁细胞和中柱细胞长度的抑制为等比例抑制,即虽然细胞体积缩小,但缩小比例相同。中柱细胞宽与薄壁细胞宽比表明,碱胁迫对薄壁细胞宽度抑制作用更显著,其中对龙粳27抑制作用达极显著,对越光抑制作用达显著。
综合以上分析表明,碱胁迫对水稻根系细胞形态产生显著或极显著影响,对薄壁细胞抑制作用强于中柱细胞。但抑制作用因品种产生差异,本研究表明,龙粳27根细胞对碱胁迫更为敏感,细胞形态和体积产生较大变化,而对越光抑制主要表现为体积缩小。
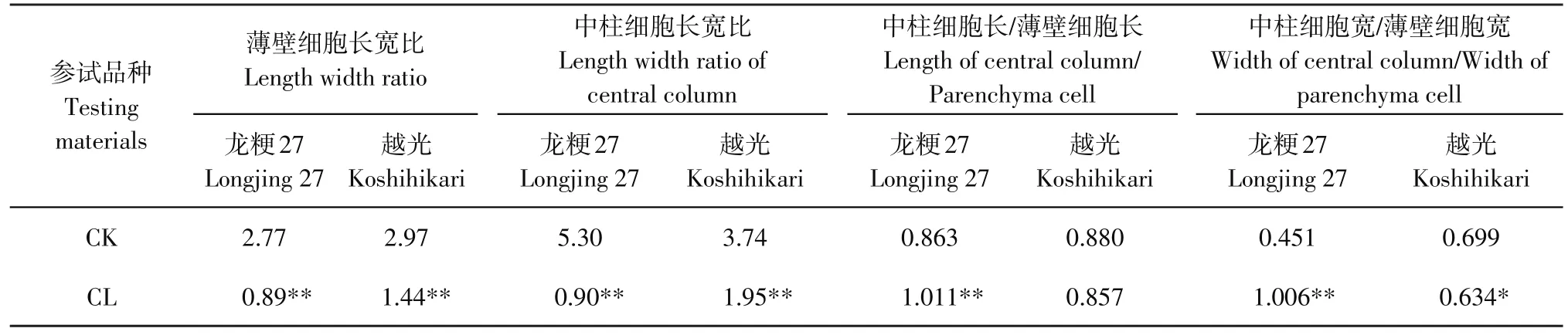
表3 碱胁迫对根细胞长度与宽度的影响Table 3 Effects of alkali stress on the length and width of root cells
2.2 碱胁迫对光合色素的影响
2.2.1 对叶绿素a、叶绿素b和类胡萝卜素的影响
由表4可知,碱胁迫后,各参试品种叶片叶绿素a、b、类胡萝卜素含量均呈极显著下降,品种间下降幅度差异显著,龙粳27和藤系138光合色素含量下降幅度均高于绥粳5号和越光,且达显著或极显著。龙粳27和藤系138叶绿素a/b值呈显著下降,表明碱胁迫对两份品种叶绿素a抑制作用强于叶绿素b;绥粳5号和越光叶绿素a/b值未下降或下降趋势不显著,表明对叶绿素a/b为等比例抑制。
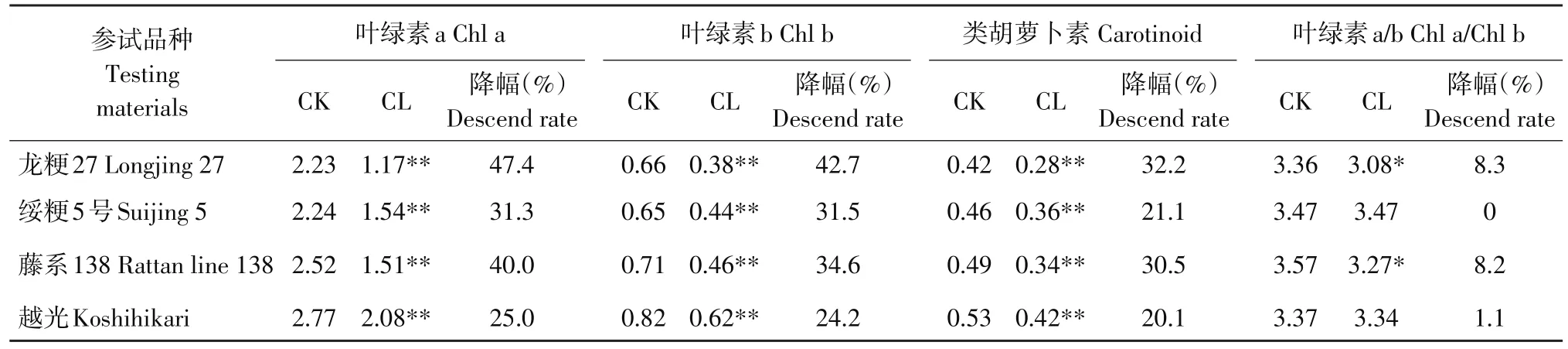
表4 碱胁迫对光合色素的影响Table 4 Effects of alkali stress on photosynthetic pigments (mg·g-1)
所有参试品种光合色素含量均极显著下降,表明碱胁迫对水稻叶片光合色素合成影响显著(见表4),品种龙粳27和藤系138叶绿素a对碱胁迫最敏感,其次是叶绿素b,类胡萝卜素敏感性相对较弱;绥粳5号和越光叶绿素a和b对碱胁迫敏感性无差异,类胡萝卜素敏感性最弱。以上分析说明,碱胁迫抑制光合色素合成,但不同水稻品种各光合色素下降幅度和敏感性差异显著。
2.2.2 叶绿体、线粒体及淀粉体在细胞中比表面积变化
叶片细胞中叶绿体、线粒体和淀粉体占细胞比表面积因品种而产生差异(见表5)。碱胁迫后,龙粳27、绥粳5号和越光叶绿体、线粒体在细胞中比表面积显著或极显著增加,藤系138极显著降低;龙粳27和藤系138淀粉体在叶绿体中比表面积极显著下降,品种越光增加,且达极显著,绥粳5号不显著;线粒体与叶绿体比表面积数据表明,龙粳27和越光线粒体面积极显著增加,藤系138极显著降低,绥粳5号不显著;线粒体和淀粉体比表面积数据表明,龙粳27和藤系138极显著增加,绥粳5号和越光不显著。
通过以上分析认为,碱胁迫对亚细胞数量和体积影响显著,这种影响将对各细胞器结构和功能产生重要作用。不同水稻品种耐碱性强弱应与各细胞器数量、体积、结构和功能变化差异联系紧密。
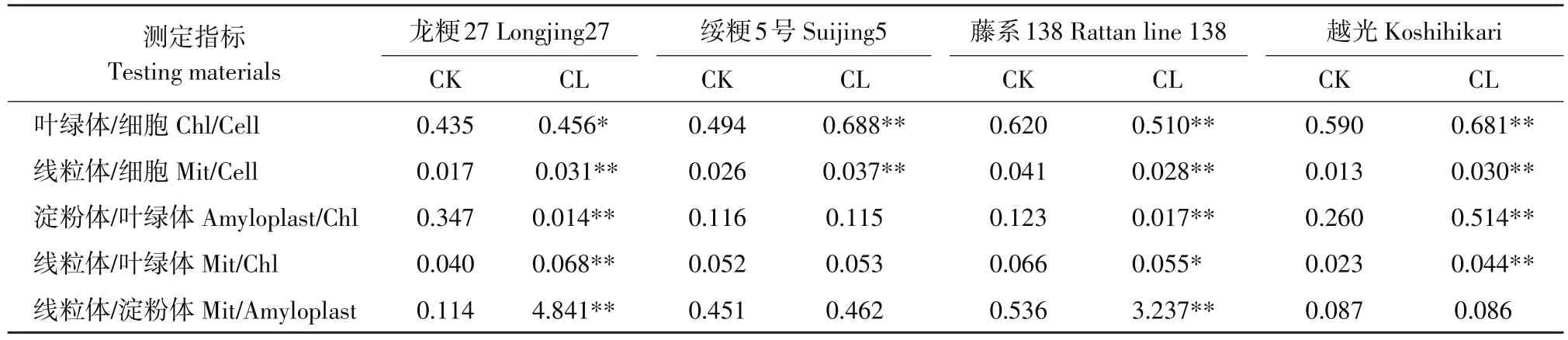
表5 超微图像中细胞器比表面积分析Table 5 Specific surface area analysis of organelles and cells in ultramicro images
2.3 碱胁迫对水稻叶片细胞超微结构的影响
2.3.1 对龙粳27超微结构的观察
龙粳27对照细胞内叶绿体呈梭形,片层结构丰富,基粒类囊体分布均匀且较宽,基粒片层较多,可见基质类囊体(见图3)。叶绿体中镶嵌大量由体积较大的淀粉粒组成的淀粉体(见图3-2),周围分布数量较少的线粒体及细胞核(图3-1,3),嗜锇颗粒少。如图3-7所示,经碱处理后,细胞中仍存在大量叶绿体,但分布凌乱且有堆积现象。叶绿体中镶嵌数量少、体积较小淀粉体。叶绿体中多数基粒片层肿胀(见图3-6),且类囊体偏向叶绿体一侧,叶绿体基粒宽度明显变细、混乱(见图3-4,8),表明类囊体降解。与对照相比线粒体仍分布在叶绿体周围,数量相对丰富,但明显肿胀,内脊稀疏,嗜锇颗粒少量增多,表明有伤害。
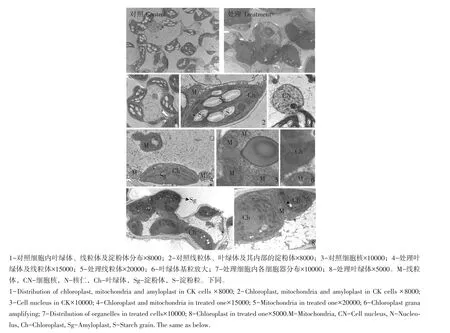
图3 龙粳27对照与0.20%碱处理超微结构Fig.3 Ultrastructure of CK Longjing 27 and the one treated with 0.2%concentration alkali
由以上分析认为,碱胁迫对龙粳27苗期微观伤害主要是叶绿体和线粒体,依叶绿体内部结构变化和所含淀粉体数量和体积情况分析,伤害影响光合产物形成,线粒体数量虽增加,但由于内脊数量少,且发生肿胀,因此呼吸消耗增强功能下降,且叶绿体合成光合产物少,淀粉合成减少,转运糖类也减少,使植物体生理机能处于紊乱饥饿状态,造成生长量减少,最终枯黄。
2.3.2 对绥粳5号超微结构的观察
如图4所示,未经碱处理绥粳5号细胞器结构清晰,线粒体呈线形、椭圆形或圆形,嵴清晰可见(见图4-1),形状丰富,表明代谢旺盛;大部分叶绿体呈梭形或椭圆形,基粒片层结构清晰,基粒厚度较宽,堆积的类囊体数量较多,两相邻基粒间距合理,连接基粒之间基质片层较为清晰(见图4-3)。镶嵌在叶绿体中淀粉体数量较多(见图4-2)。如图4-5所示,处理后细胞叶绿体上嗜锇颗粒丰富,叶绿体数量多,线粒体形状各异,内嵴丰富,大量分布在叶绿体周围(见图4-5、6),叶绿体内部基粒之间距离不均匀,基粒之间基质类囊体清晰,部分叶绿体基粒排序走向曲折紊乱,表明受害较重。部分叶绿体含有大量体积巨大淀粉粒(见图4-8),将叶绿体基粒挤压至边缘,但基粒较清晰。部分淀粉体内小淀粉粒与大淀粉粒之间有连丝相接,大淀粉粒又与淀粉体叶绿体被膜之间以连丝相接,增殖的小淀粉粒亦通过连丝由大淀粉粒提供淀粉化合物继续生长,表明碱处理后,绥粳5号淀粉合成与分化较为旺盛。
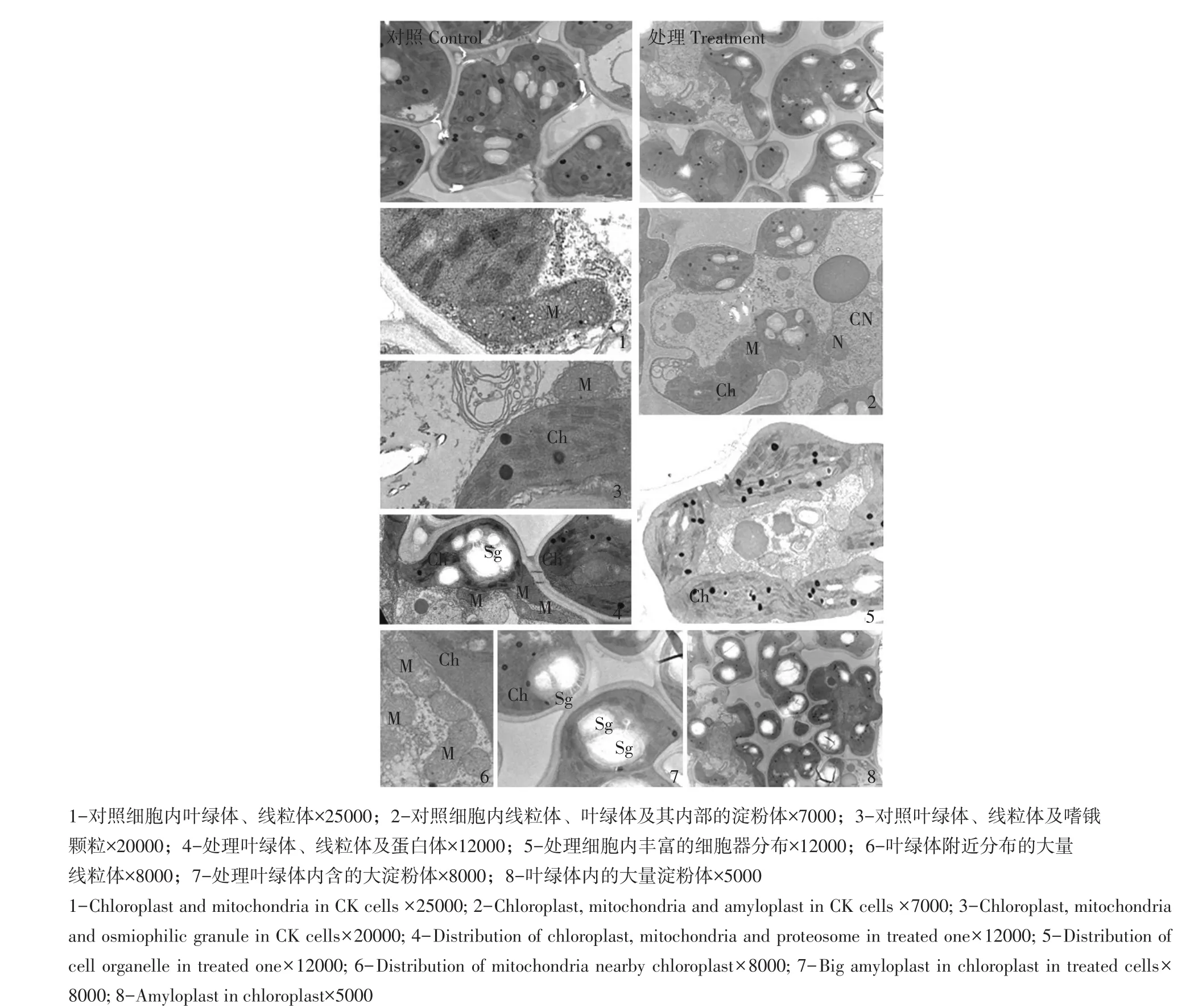
图4 绥粳5号对照与0.20%碱处理超微结构Fig.4 Ultrastructure of CK Suijing 5 and the one treated with 0.2%concentration alkali
与对照相比,叶绿体内淀粉体体积较大,说明碱处理后对绥粳5号叶绿体内淀粉合成酶活力影响较小,光合产物存储现象严重,图4-7淀粉连丝表明淀粉合成、储存、分生分化功能依然较强,淀粉粒体积过大,导致部分叶绿体受损较重,说明淀粉体分化分生被动发生,产生这种现象原因应在糖类运移环节,既光合产物运出叶绿体效率低,致使淀粉体体积过大。线粒体内嵴较清晰,表明其受损情况较轻。以上分析表明碱胁迫后,绥粳5号叶片内部分叶绿体形状虽变化较大,但保持一定光合功能,打破光合产物运移平衡,是造成生理紊乱的主要原因。绥粳5号综合表现受害较轻,是一种耐盐碱较强品种。
2.3.3 对藤系138超微结构的观察
如图5所示,大量线粒体分布在对照叶绿体周围(见图5-1),结构正常,内脊比较清晰。叶绿体形状呈梭形,基粒分布广泛均匀,结构正常,可见基粒类囊体和基质类囊体,叶绿体内含较丰富淀粉体(见图5-2,3)。
处理后大量线粒体分布在叶绿体周,且内脊清晰,说明其代谢功能旺盛,叶绿体内基粒厚度明显变薄(见图5-4)或肿胀(见图5-6),排序紊乱,内含淀粉体颗粒减少,嗜锇颗粒数量增多,嗜锇程度变深。
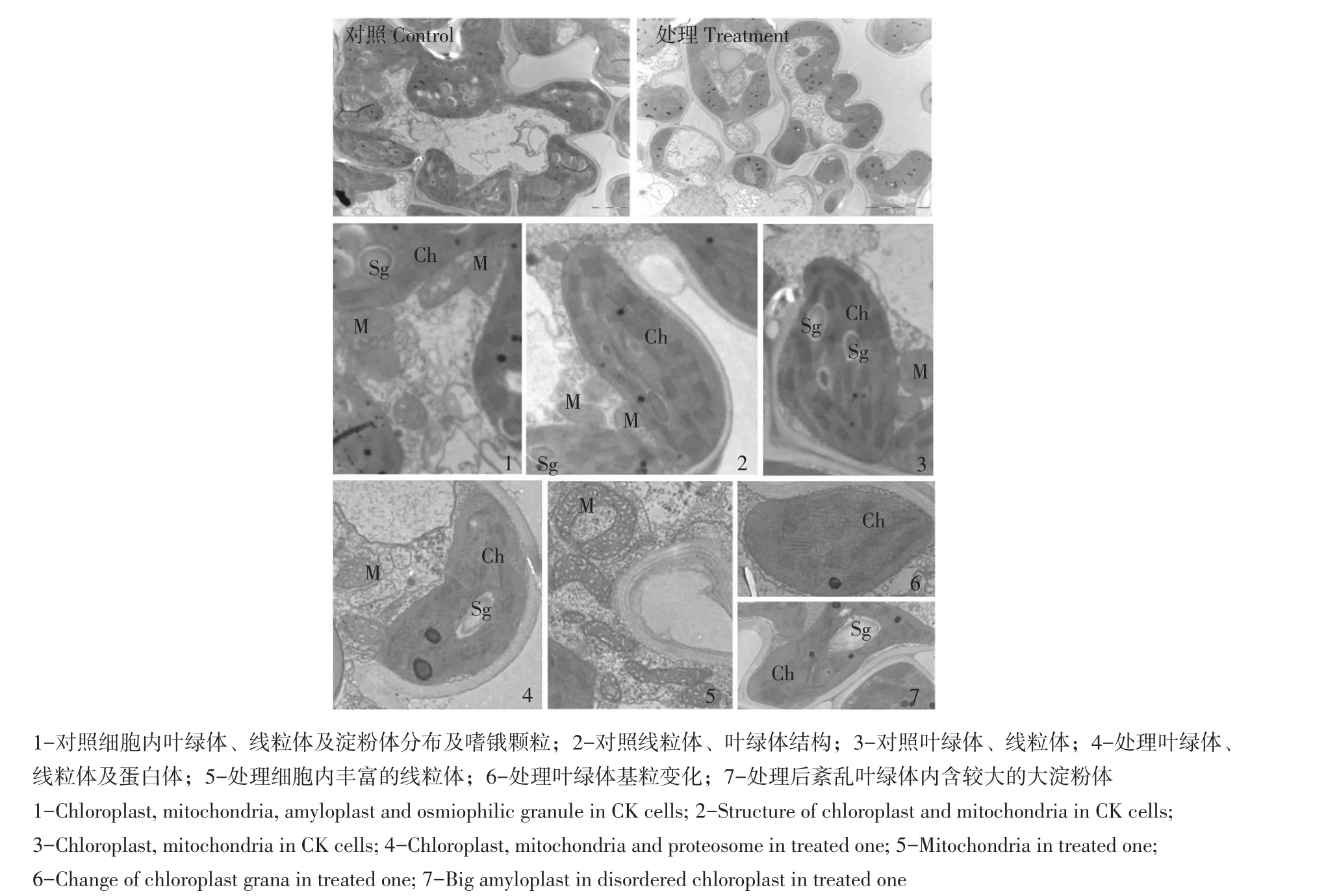
图5 藤系138对照与0.20%碱处理超微结构Fig.5 Ultrastructure of CK Tengxi 138 and the one treated with 0.2%concentration alkali
碱胁迫后藤系138叶绿体基粒类囊体数量减少,部分球形化,排序紊乱,合成淀粉体数量减少,但对线粒体结构与功能、糖类运移功能等影响较小。综上所述认为,碱胁迫主要破坏藤系138叶绿体结构,因此光合功能下降,合成产物减少,因此细胞代谢原材料供应不足,是造成植株受害严重主要原因。
2.3.4 对越光超微结构的观察
对照叶绿体镶嵌数量丰富的淀粉体,大部分叶绿体形状呈梭形,基粒类囊体数量较多,排列有序。线粒体数量较多,形状各异,分布在叶绿体周围(见图6-2),内脊较多且清晰(见图6-3),表明代谢旺盛。碱处理后淀粉体数量和体积明显大于对照,每个淀粉体内均有1~3个较大淀粉粒,围生着数量更多的小淀粉体(见图6-4)。处理后由于淀粉体体积增加过大,导致叶绿体形状呈近圆形,基粒狭长,类囊体变稀疏(见图6-5,8),但广泛分布在叶绿体中。线粒体数量相比对照明显下降且肿胀(见图6-6,9),内脊数量减少,说明代谢功能受影响,呼吸作用增强。
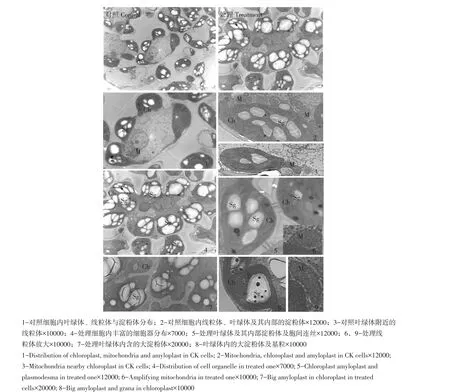
图6 越光对照与0.20%碱处理超微结构Fig.6 Ultrastructure of CK Koshihikari and the one treated with 0.2%concentration alkali
碱处理后叶绿体类囊体变化较大,但结构清晰,因此光合功能影响相对较小。根据淀粉体数量及体积变化表明,淀粉体向葡萄糖转化受阻。线粒体受害较重,因此品种越光受害生理特征为,胁迫后叶绿体结构虽受影响,但光合作用仍继续,糖类转运功能受阻,线粒体结构和功能受严重破坏,造成合成的光合产物大量积存于叶片中,形成的淀粉体无法转运,植株生长受阻。
3 讨论与结论
3.1 胁迫对植物根细胞的伤害
植物细胞在胁迫条件下,其形态发生相应变化,分生组织发生非正常分化[17],郭望模等指出盐胁迫下叶片下表皮细胞变小[18],本研究发现薄壁细胞和中柱细胞长度极显著缩短,与此结论相似。内外皮层细胞栓质化与木质化被认为是植物适应逆境的重要特征,植物根部通过木质化促进水分子运输,并限制Na+、Ca2+、Mg2+等离子自由运移到外部,以提高抗逆性[19-20],本研究观察到皮层细胞木质化现象,表明碱胁迫下水稻根系结构向适应逆境转变。维管束是主要输水组织,黄振英等报道显示部分胁迫可促使植物根系维管柱径向加粗、木质化程度加深,导管数量增加,以提高根运输能力[19,21],但本研究表明碱胁迫造成水稻根系大维管束数量减少,发育未完全的维管束增多,但部分图像及数据显示,维管束所占根径向面积比增加,可能是水稻根系对逆境环境的适应性响应,与上述报道结果一致。
3.2 胁迫对植物细胞亚结构的解析
关于逆境胁迫植物细胞超微结构变化,国内外学者均认为,逆境对植物叶绿体、线粒体等细胞器超微结构产生破坏或降解作用[22-23]。本研究表明,碱胁迫导致叶绿体变形,基粒肿胀或片层数量减少,但此种变化在品种间存在差异,部分学者研究结论与此观点一致[4,10]。超微结构变化是复杂和动态的过程,保持逆境中生理功能平衡是微观结构改变的根本原因。因此无法单一从某一个细胞器数量、形态或位置改变而定性。本研究通过观察和分析分别来自中国和日本水稻品种,发现微观结构变化具有综合性或整体性特点,同时这种变化具有共性和品种个性差异的特征。
叶绿体是对逆境胁迫最敏感的细胞器,本研究发现基粒类囊体片层结构减少或部分减少,通过基质连接的部分类囊体排列呈混乱或扭曲状态;部分水稻品种其类囊体偏向叶绿体一侧(龙粳27),更多品种(绥粳5号、藤系138、越光)类囊体位置变化不明显。线粒体数量和结构变化也因品种而异,龙粳27线粒体数量相对增加,肿胀明显,内脊变少,呼吸作用强烈,消耗增高,无法开展三羧酸循环与氧化磷酸化途径。同时合成淀粉粒体积大幅下降,表明叶绿体功能受损严重,光合产物3-磷酸甘油酸减少,机体生理运转效率严重降低,该品种整体机能处于匮乏状态。碱胁迫后绥粳5号叶绿体中存储大量体积巨大的淀粉粒,前人研究也发现类似现象,并依此判断植物抗逆性状[24]。胁迫后绥粳5号部分淀粉粒之间以及部分大淀粉粒与细胞膜之间产生连丝。本研究认为,受到胁迫且结构遭破坏的叶绿体,光合功能无法增强,由于光合作用合成的糖类无法正常输出而迅速转化合成淀粉,导致淀粉在叶绿体中大量累积形成淀粉粒;大淀粉粒通过连丝向细胞膜输送转化的糖类,但膜系统已受到损伤[25],大部分糖类又转运形成淀粉颗粒,或分生出小淀粉粒,但这种分化分生合成过程为被动存储。胁迫产生大量线粒体,线粒体轻微肿胀,说明呼吸作用增强。嗜锇颗粒变化可能是由于叶绿体类囊体膜降解导致脂质类物质在叶绿体或液泡中大量聚集。品种藤系138,胁迫后线粒体结构较为正常,叶绿体损伤严重,叶绿体基粒类囊体崩解(见图5-5,6),说明已无法光合作用,其他存储少量淀粉粒的叶绿体,类囊体结构严重扭曲,较大淀粉体位于叶绿体中央位置,可能已失去转化功能。尽管线粒体形态和功能正常,但缺少能源供应。处理后品种越光叶绿体内淀粉体数量和体积明显大于对照,致使叶绿体形状近于圆形,类囊体片层减少,走向扭曲。与绥粳5号相似,但两者差异明显,即越光无连丝现象,因此淀粉体与糖之间的转化过程相比绥粳5号较弱,处理后越光线粒体表面比极显著增加,但结构模糊,说明呼吸消耗增大,功能受损较重,虽叶绿体参与光合作用,但糖类转化和光合产物转运受阻。
