基于CLIMEX和ArcGIS的灰茶尺蠖在中国的潜在适生区预测
2020-12-15陈李林周浩赵杰
陈李林,周浩,赵杰
基于CLIMEX和ArcGIS的灰茶尺蠖在中国的潜在适生区预测
陈李林1,2,周浩1,2,赵杰3
1. 闽台作物有害生物生态防控国家重点实验室,福建农林大学植物保护学院,福建 福州 350002;2. 福建农林大学安溪茶学院,福建 安溪 362406;3. 西北农林科技大学资源环境学院,陕西 杨凌 712100
基于中国820个气象站点数据、灰茶尺蠖已知地理分布资料和生物学数据,结合CLIMEX模型与ArcGIS软件模拟预测灰茶尺蠖在中国目前及未来的潜在分布范围,评估灰茶尺蠖的潜在分布范围及气候变暖对其分布的潜在影响。结果表明,灰茶尺蠖在中国目前的适生区主要分布在3°51′N~40°6′N,适生区面积占全国总面积的34.27%。中国大部分省份的气候条件均适合灰茶尺蠖种群生长。随着气候的变化,灰茶尺蠖在中国的潜在适生区面积总体增幅不大,但其组成变化较大。至2050年,预测灰茶尺蠖高度适生区面积占比达最大(22.23%)。相比各种能源之间的平衡情景(A1B),较高能源需求情景(A2)下灰茶尺蠖在陕甘宁地区向内陆扩张更快。灰茶尺蠖在中国适生区分布广泛,应加强灰茶尺蠖预测预报,趁早采取防控措施,以保障茶叶优质安全生产。
灰茶尺蠖;气候变化;适生区;潜在分布;预测
茶园尺蠖类害虫种类较多,灰茶尺蠖(Warren)是发生最普遍、分布最广、为害最严重的物种之一,隶属鳞翅目(Lepidoptera)尺蛾科(Geometridae)灰尺蛾亚科(Ennominae)。由于灰茶尺蠖成虫繁殖力和幼虫活动力均较强,且完成一个世代时间较短,发生代数较多,世代重叠严重,极易暴发成灾。大发生时使茶园大面积秃枝,状如火烧,甚至绝产,叶片被取食光的茶树冬季易受冻害,致使茶树早衰,严重影响茶叶的品质和产量[1-3]。2019年4月25日,农业农村部发布农事指导“2019年茶树主要病虫害防控技术方案”,将灰茶尺蠖和茶尺蠖(Prout)列为中国四大茶区重点防控的茶树害虫(www.moa.gov.cn)。灰茶尺蠖和茶尺蠖均以茶叶为食,是形态和习性极其相似的两个近缘种,肉眼几乎难以区分,在过去的生产活动中常被误认为是同一物种,统称“茶尺蠖”[4-5]。白家赫等[5]和罗宗秀[6]对灰茶尺蠖和茶尺蠖两近缘种在全国的分布研究表明,灰茶尺蠖广泛分布于中国各大产茶区,而茶尺蠖主要分布于浙江、江苏、安徽茶区,且与灰茶尺蠖混合发生;据此,本研究将浙江、江苏、安徽以外的省(市、自治区)茶尺蠖的相关分布资料均作为灰茶尺蠖的分布参考资料。
前人研究发现,灰茶尺蠖的生长发育受温度、湿度、光照等因素影响,其中温度对其生长发育影响最大[7-8]。葛超美[7]研究发现,在19~27℃,随着温度升高,灰茶尺蠖各虫态及世代的发育历期显著缩短,卵、幼虫、蛹及世代的发育速率与温度呈极显著正相关;灰茶尺蠖幼虫存活率无显著变化,而预蛹存活率在27℃时显著降低;灰茶尺蠖生长发育的适宜温度为21~23℃,在此温度下,种群增长指数达到峰值且雌成虫产卵量较高。逆境对茶尺蠖发育的影响研究表明,随着温度升高(24~32℃),蛹的发育历期明显缩短,羽化率呈先升高后降低的趋势,在32℃时最低(19.06%)[9]。高温下,雌蛹早于雄蛹羽化,且雌雄比例提高[9]。除了高温胁迫,低温对茶尺蠖发育的影响也十分明显。董道青等[10]研究发现,随冷藏时间延长,茶尺蠖蛹羽化率下降、成虫发育畸形率增加;蛹龄3~4 d的蛹冷藏15~30 d,其羽化率急剧下降,且羽化成虫发育畸形。前人研究结果表明,灰茶尺蠖卵、幼虫、蛹、成虫各虫态发育起点温度分别为8.80~10.84℃、4.57~7.70℃、6.00~9.44℃、10.78~14.91℃,有效积温570.03~631.59 d·℃[11-13]。空气相对湿度为80%~90%,卵孵化率最高,若相对湿度低于75%,卵孵化率和成虫羽化率降低;平地茶园灰茶尺蠖发生相对较重,高山茶园发生相对较轻[8]。
中国国土广袤,大部分区域的气候条件适合灰茶尺蠖生长发育,其寄主植物茶树在中国广泛种植,且近年来许多省(市、自治区)大面积引种茶树,故中国很多省(市、自治区)都有该害虫的踪迹。由于灰茶尺蠖具有较强的飞行和迁移扩散能力[14],在全球气候变暖趋势下,该害虫极大可能向其他未发生省(市、自治区)茶园迁移。所以,对灰茶尺蠖在中国目前及未来的潜在适生区进行分析评估,对促进茶园害虫防控具有重要意义。物种适生性常用模型主要有CLIMEX、MaxEnt、GARP等,CLIMEX模型作为物种分布模型(Species distribution model,SDM),以探究气候对物种的影响来预测物种分布,并作为风险分析工具对物种定殖风险进行预评估。CLIMEX模型采用物种功能特征和生理耐受性来进行建模,计算生长和压力指数,确定反映一个地区适宜性的生态气候指数(Ecoclimatic index,EI),预测物种的时空分布。EI是通过生长和压力指数结合建立的年度气候适宜性指数,该指数描述了通过气候参数计算的物种气候适宜性,范围从0到100。EI值近似为0表示物种长期存活的环境不利,超过30表明气候非常有利。通过EI值,CLIMEX建模绘制昆虫、植物等物种的潜在分布,被广泛用于研究生物入侵、潜在反应和气候变化之间的相互作用,评估害虫种群建立的风险[15-16],如西部樱桃实蝇(Curran)在哥伦比亚、印度、印度尼西亚、马来西亚、台湾、泰国、委内瑞拉和越南的潜在分布[17],埃及伊蚊(L.)分布及其传播登革热病毒的风险[18],以及为害番茄的重要害虫烟粉虱(Gennadius)的种群动态和分布[19]等。ArcGIS具有空间数据统计分析与输出功能,CLIMEX模型与ArcGIS软件相结合实现了由点到面的过渡,提高了分析结果的准确性,能直观地展示预测物种目前和未来的潜在适生区和适生程度图。
本研究根据新闻报道、文献资料等,收集了灰茶尺蠖的已知地理分布和生物学数据,利用CLIMEX 2.0的比较位点模型(Compare locations “1 species”)[20]、ArcGIS 10.4.1软件的插值与叠加功能对灰茶尺蠖在中国目前和未来的潜在适生区做出预测,模拟气候变化条件下灰茶尺蠖在中国的时空分布图,绘制灰茶尺蠖在中国目前和未来的潜在适生区的直观分布图。对目前尚未出现灰茶尺蠖为害以及有潜力成为茶区的省份做出预警,为制定科学的茶园监测和防控体系提供理论依据和技术支持。
1 材料与方法
1.1 数据资料
1.1.1 国内灰茶尺蠖已知地理分布
根据文献资料,灰茶尺蠖分布于中国浙江[5-6]、江苏[6,21]、山东[22]、河南[6,23-24]、湖北[25-28]、湖南[6]、安徽[6,21,29-30]、江西[31-33]、福建[2,6,21]、广东[6,34]、广西[6,35-36]、云南[21,37]、贵州[6,38-39]、四川[6,40-42]、重庆[43]、陕西[44],结合新闻报道及个人采样发现,甘肃、西藏、海南等省(市、自治区)茶园也有分布。
1.1.2 气象数据
CLIMEX 2.0软件自带中国86个气象站点的气象数据,通过整理由中国气象信息中心提供的中国气象站点数据(http://data.cma.cn),将其导入CLIMEX 2.0,使中国的气象站点数增加到820个。由于下载的气象数据集不包含中国台湾气象站点数据,因此不考虑灰茶尺蠖在中国台湾地区的潜在适生区预测分析。
未来的气象数据下载于CLIMond网站(www.climond.org),采用联合国政府间气候变化专门委员会(IPCC)第四次评估报告中提出的,A1B(各种能源之间的平衡)和A2(较高能源需求)两种温室气体排放情景下,空间分辨率为30′的2030、2050年和2070年的气候数据[45]。
1.1.3 地图矢量数据
本研究用图基于国家测绘地理信息局标准地图服务网站(http://bzdt.ch.mnr.gov.cn)下载的审图号为GS(2016)2923号的标准地图制作,底图无修改。
1.2 预测模型与分析软件
根据灰茶尺蠖在中国的已知地理分布,采用CLIMEX 2.0的比较位点模型,并结合ArcGIS 10.4.1的插值功能(反距离权重法,Inverse distance weighting,IDW)对灰茶尺蠖在中国的潜在分布进行预测。CLIMEX共预设42个生物学参数,本研究根据灰茶尺蠖的生物学资料和已知地理分布,选用了CLIMEX中的5个温度参数(DV0、DV1、DV2、DV3、PDD)、4个湿度参数(SM0、SM1、SM2、SM3)和4种胁迫及胁迫积累速率(TTCS、THCS、TTHS、THHS、SMDS、HDS、SMWS、HWS)。参考CLIMEX 2.0自带的与灰茶尺蠖分布相似的温带气候参数值,设定灰茶尺蠖相应参数的初始值,并根据收集到的灰茶尺蠖的生物学资料和已知地理分布进行反复调试,使得当前气候条件下灰茶尺蠖在中国的预测分布范围与已知地理分布范围达到最佳拟合,以此组参数值作为预测灰茶尺蠖适生区的参数值(表1)。
温度参数是根据文献报道、室内试验及灰茶尺蠖在中国的已知地理分布进行调试确定。极端高温数据由室内试验获得。室内研究发现,灰茶尺蠖高龄幼虫在32℃生长发育速度极快,能够正常存活,但羽化后的成虫全部畸形,且产卵量极低;低龄幼虫则出现滞育现象。结合其在中国的已知地理分布进行调试后,最终将温度上限DV3设为35℃。根据灰茶尺蠖室内种群在相对湿度低于30%时,对幼虫生长发育产生极大影响,会导致幼虫死亡,且成虫产卵量极低,最终将湿度下限SM0设为0.2,使得甘肃南部、山东等地区满足分布条件。有效积温PDD根据文献资料[7,10-11]中尺蠖各虫态发育起点温度及有效积温进行调试,并最终赋值为644.44。冷胁迫(CS)指数则根据灰茶尺蠖在中国分布的北部边界和PDD进行调试,通过调试冷胁迫开始积累的阈值TTCS和冷胁迫积累速率THCS调整灰茶尺蠖的分布北界,使其满足在山东、河南北部、陕西中部、甘肃南部以及西藏南部的分布,最终将TTCS设为–4℃,THCS设为–0.003 3。热胁迫(HS)开始积累的阈值TTHS和热胁迫(HS)积累速率THHS的赋值主要通过灰茶尺蠖在中国南部省份的已知地理分布进行调试,最终将TTHS设为35℃,THHS设为0.001 4。干胁迫(DS)主要通过云南和四川这两个省进行调试,使得灰茶尺蠖在云南北部以及四川中部地区满足分布条件,将干胁迫开始积累的阈值SMDS设为0.2,干胁迫积累速率HDS设为–0.053。湿胁迫(WS)不是限制灰茶尺蠖在中国分布的主要因素,将湿胁迫开始积累的阈值SMWS设为1.5,湿胁迫积累速率HWS设为0.000 2,使其符合灰茶尺蠖在中国东南沿海地区的分布。
根据灰茶尺蠖在中国的已知地理分布资料和发生情况,利用表1参数值计算出灰茶尺蠖在中国820个气象站点的EI值,将灰茶尺蠖在中国的适生区分为4类(表2)。随机选取70%的分布数据用于建模,30%的数据用于模拟结果的验证,当验证点均散落在预测得到的适生区内,即模拟结果与灰茶尺蠖已知地理分布具有较好的拟合,说明设定的CLIMEX参数值符合条件,能够较好的用于模拟预测灰茶尺蠖目前以及未来的适生区。利用ArcGIS 10.4.1地图处理模块中的IDW将预测得到的EI值进行插值分析,并结合灰茶尺蠖在中国已知地理分布,对EI值进行分类,得到灰茶尺蠖在中国的适生区分布以及适生程度图。
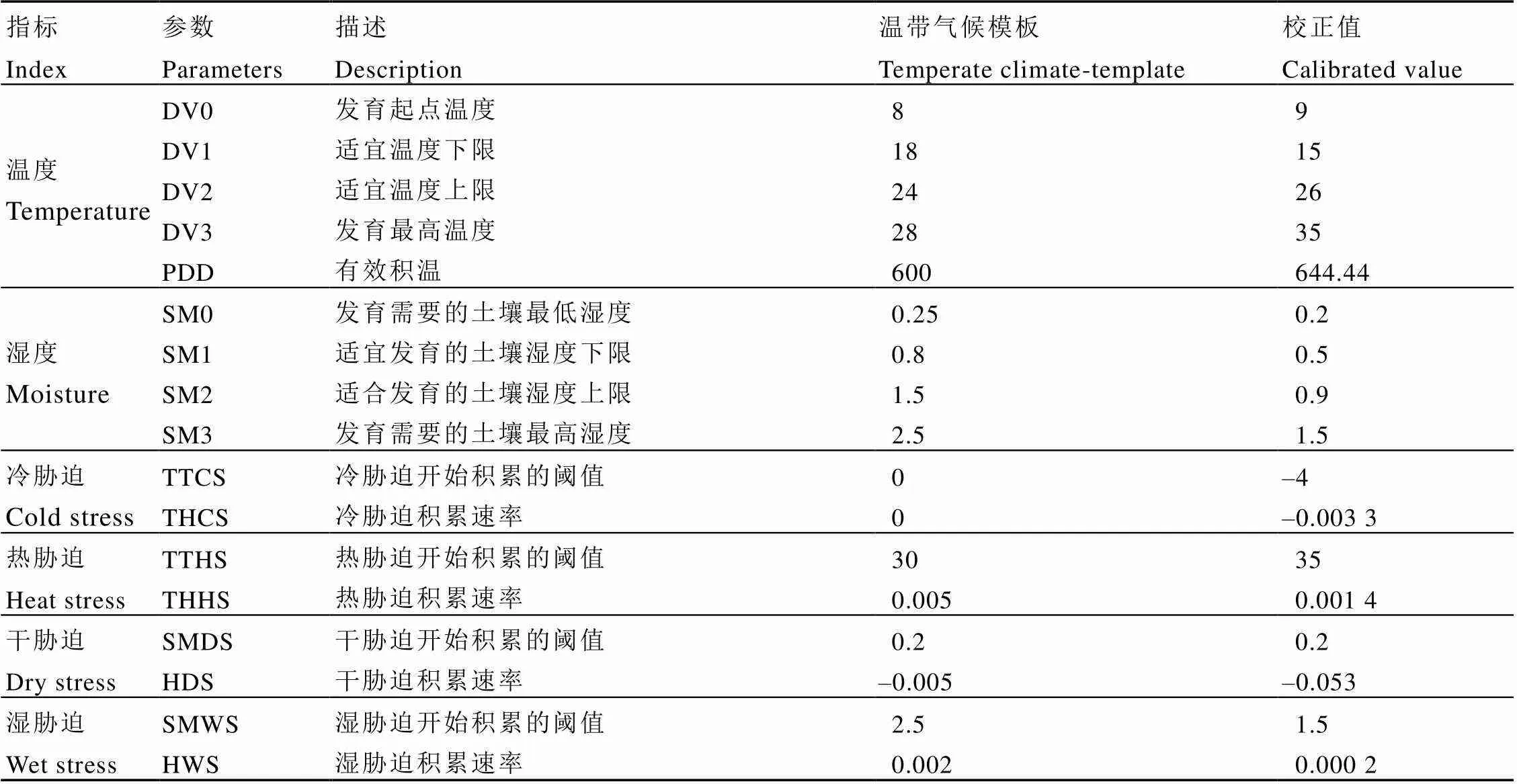
表1 预测灰茶尺蠖在中国分布的CLIMEX参数值
2 结果与分析
2.1 目前气候条件下灰茶尺蠖在中国的适生区分布及适生程度
基于预测得到的灰茶尺蠖在中国的分布与其在中国已知地理分布吻合最佳的CLIMEX参数值及适生等级评判标准,分析中国820个气象站点在目前气候条件下的EI值,得到灰茶尺蠖在中国各个气象站点的适生区及其适生程度。经预测,灰茶尺蠖在中国有459个适生点(EI>0),占全国站点的55.98%。目前气候条件下灰茶尺蠖在中国的适生区主要分布在3°51′N~40°6′N,中国大部分省份的气候条件均适合灰茶尺蠖种群生长(图1)。对插值结果进行重新分类、矢量数据转换等处理后,利用ArcGIS计算面积得知灰茶尺蠖在中国的目前适生区总面积约329.01万km2,占中国总面积的34.27%,其中高、中、低度适生区分别占比18.90%、8.45%、6.92%(图2)。高度适生区主要包括海南、广西、广东、福建、江西、江苏、湖北、重庆和贵州的大部分地区以及浙江东北部、安徽北部、湖南中部、河南南部、山东南部、陕西南部、甘肃东南部、四川东部、西藏南部、云南东南部和西部(图1)。中度适生区主要包括浙江和湖南的大部分地区、湖北南部、安徽南部、云南中部、广西东北部以及山东、河南、陕西、甘肃、四川、贵州和西藏的小部分地区(图1)。低度适生区呈带状穿过辽宁南部、山东、河南北部、山西南部、陕西北部、甘肃南部、四川中部、云南北部以及西藏南部部分地区(图1)。综上,中国的气候条件十分适合灰茶尺蠖的种群建立。

表2 适生等级评判与适生程度划分

图1 灰茶尺蠖在中国的适生区及适生程度(审图号:GS(2020)4372号)
2.2 未来气候条件下灰茶尺蠖在中国的适生区分布及适生程度
根据IPCC的第4次评估报告,温室气体在A1B和A2两种排放情景下,对灰茶尺蠖在中国的未来分布区域进行预测。结果发现,随着气候变化,灰茶尺蠖在中国的潜在适生区持续缓慢增大,虽幅度不大,但其组成变化较大。适生区总面积由目前的34.27%逐步缓慢增加到2070年的34.62%,其中低度适生区向内陆扩张幅度不大,只有西藏南部沿国界地区、陕甘宁中部及辽东半岛低度适生区面积在逐渐增大,而云南原本属中高度适生区的大部分地区适生程度逐渐降低,使得低度适生区面积进一步增大。从2051年到2070年,广东和广西部分地区适生程度也开始下降,因此,灰茶尺蠖的低度适生区面积整体占比不断增大。从2031年到2050年,由于云南中高度适生程度有所下降及贵州和浙江中度适生程度的不断上升,使得中度适生区面积呈下降趋势;从2051年到2070年,广东和广西原本高度适生的部分地区适生程度下降,从而中度适生区占比上升。至2050年,预测灰茶尺蠖高度适生区面积占比达到最大(22.23%),贵州、重庆、湖南、湖北、江西、浙江、江苏、福建、广东、广西的几乎所有县市均为灰茶尺蠖的高度适生区(图2和图3)。
比较两种情景下灰茶尺蠖在中国未来的适生区及适生程度,发现两种情景下的未来气候数据预测得到适生区的变化较为相似,适生区均向北移动,且辽东半岛和西藏的适生区扩张相似,而在陕甘宁地区,A2情景相比A1B情景,适生区向内陆扩张范围更大(图4)。在对目前气候条件下灰茶尺蠖的适生性分析时,云南省楚雄州(24°45′N,101°15′E)DS=54,EI=12;当纬度提高1.5°后,DS=98,EI=0;随着纬度继续提高,四川省凉山州(27°45′N,101°15′E)DS=118,此时开始产生冷胁迫,CS=3,EI=0,推测干胁迫为灰茶尺蠖在云南省的主要限制因素。在对未来气候条件下灰茶尺蠖的适生性分析时,云南省思茅市(23°15′N,101°45′E)在目前气候条件下EI=25,DS=17;随着气温升高DS值逐渐增大,到2070年DS=50,EI=12,该地区适生程度降低,推测限制云南灰茶尺蠖分布的主要因素为干胁迫。广东省茂名市(21°45′N,110°45′E)在目前气候条件下EI=41,是灰茶尺蠖的高度适生区;随着气温逐渐变暖,到2030年,其EI=27,并从2050年开始产生热胁迫,HS=10,EI=17;2070年该地区HS=44,EI=8,适生程度从高度适生区降为低度适生区。在全球气候变暖的趋势下,灰茶尺蠖在中国的适生区增大、适生程度提高,与A1B相比,A2情景下灰茶尺蠖在陕甘宁地区向内陆扩张更快(图4)。
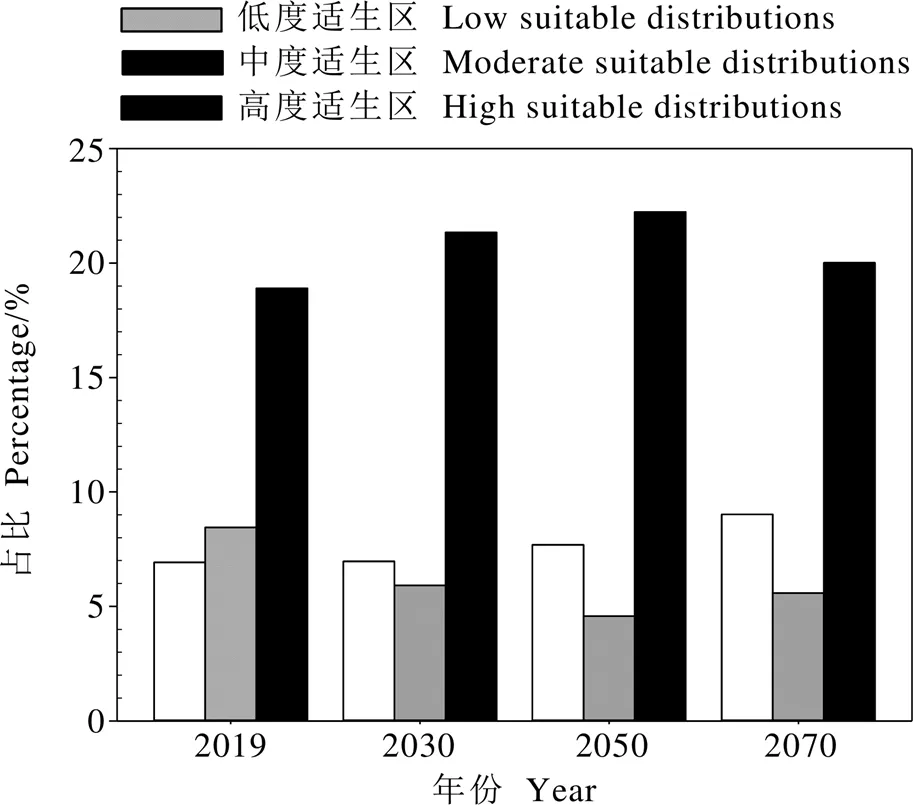
图2 灰茶尺蠖在中国目前和未来各等级适生区的面积占比

图3 温室气体在A2和A1B两种排放情景下灰茶尺蠖在中国未来的适生区及适生程度(审图号:GS(2020)4372号)
中国大部分省份的气候条件均适合灰茶尺蠖的种群生长。通过CLIMEX模型预测,当温度低于–4℃,冷胁迫以THCS=–0.003 3的速率开始积累,如陕西省安康市(33°15′N,108°45′E)CS=2,铜川市(35°15′N,108°45′E)CS=64,榆林市(37°15′N,108°45′E)CS=363。冷胁迫和纬度呈显著正相关(=0.971,=0.00),可见冷胁迫限制灰茶尺蠖向北进一步扩散。当湿度参数低于0.2,干胁迫以HDS=–0.053的速率开始积累,如四川省广元市(32°15′N,106°15′E)DS=0,绵阳市(32°15′N,104°15′E)DS=44,阿坝州(32°15′N,102°15′E)DS=144。干胁迫与经度呈显著负相关(=–0.979,=0.00),可见干胁迫限制灰茶尺蠖向西进一步扩散。热胁迫仅存在于新疆个别地区,如哈密(42°45′N,92°15′E)HS=8,吐鲁番(42°45′N,89°15′E)HS=40,即热胁迫不是限制灰茶尺蠖在中国种群建立的主要因素。同样,湿胁迫对灰茶尺蠖在中国的分布影响甚微。综上,目前气候条件下,灰茶尺蠖在中国分布的主要限制因素是低温胁迫和干胁迫。
2.3 灰茶尺蠖世代发生数及生长指数拟合
CLIMEX模型拟合的灰茶尺蠖在中国的分布区包含了现有的分布区。进一步使用CLIMEX模型对灰茶尺蠖在中国各分布区的世代发生数及生长指数(Growth index,GI)进行拟合。结果表明,灰茶尺蠖在中国浙江、安徽、江苏、湖北、湖南、江西、贵州的世代发生数平均为5代左右,福建、广东、广西、云南每年平均发生5~6代,少数地区可达7代。以浙江省绍兴市(29°45′N,120°45′E)为例,灰茶尺蠖在该地区全年的生长指数存在两个峰值,分别在4月中旬和10月下旬,而温度较高的6月—8月,生长指数一直处于谷底(图5)。

图4 A1B和A2两种温室气体排放情景下灰茶尺蠖在中国适生区变化(审图号:GS(2020)4372号)

图5 CLIMEX预测灰茶尺蠖在浙江省绍兴市的周生长指数
3 讨论
温度和湿度是影响灰茶尺蠖分布的主要因素,冷胁迫和纬度呈显著正相关,限制灰茶尺蠖向北扩散;干胁迫和经度呈显著负相关,限制灰茶尺蠖向西扩散。灰茶尺蠖在中国目前的适生区主要在3°51′N~40°6′N,大部分省份的气候条件均适合灰茶尺蠖的种群建立和生长。随着气候的变化,灰茶尺蠖在中国的适生区持续缓慢增大,虽然总体面积增加幅度不大,但其不同适生等级与不同适生程度面积组成变化较大。至2050年,灰茶尺蠖高度适生区面积占比达到最大,贵州、重庆、湖南、湖北、江西、浙江、江苏、福建、广东、广西的几乎所有县市均为灰茶尺蠖的高度适生区,尤其是浙江、湖南两省的高度适生区由局部上升为绝大多数或全部区域;同时,西藏南部沿国界地区、陕甘宁地区中部及辽东半岛低度适生区面积在逐渐增大,与张晓玲等[46]预测的未来气候(2070年)变化下茶树适宜分布区扩张规律相符。该研究发现,中国未来将保持大部分原有的茶树适宜分布区,并且在中高纬度地区出现茶树适宜分布区的扩张;中国是世界上茶树适宜分布面积增加最多的国家,在RCP 2.6(代表性的浓度路径,Representative concentration pathways,RCPs)情景和RCP 8.5情景下,茶树适宜分布面积分别增加2.7%和5.2%。茶树种植管理者们应因地制宜、科学有效地防控灰茶尺蠖,充分发挥茶园生态系统服务功能,尤其是生态茶园的保益控害生态系统服务功能。充分利用茶园生态系统的自我调节能力,保护茶园的景观多样性和生物多样性,做好灰茶尺蠖的预测预报,明确其防治适期,协调利用农业防治、物理防治、生物防治和化学生态防治等绿色防控措施,将灰茶尺蠖控制在经济阈值水平以下,杜绝长期大量使用化学农药。
通过对灰茶尺蠖在中国各茶区的世代发生数及生长指数的拟合分析,发现灰茶尺蠖在中国浙江、安徽、江苏、湖北、湖南、江西、贵州的世代发生数平均为5代左右,福建、广东、广西、云南平均每年发生5~6代,少部分地区可达7代。本研究发现浙江省绍兴市的灰茶尺蠖世代发生数约为5代,与葛超美[11]的研究结果有一定的差异,葛超美田间观察灰茶尺蠖在该地区的世代发生数为6~7代,推测可能因为茶园田间环境较为复杂,如不同生境管理方式、茶园内温度等微气候存在差异[47-48];还有茶园海拔高度、食物充足与否、防控措施、天敌等原因,以及温度对灰茶尺蠖的生长发育历期影响大、世代重叠严重等造成差异[8,49-52]。湖北省灰茶尺蠖的世代发生数及生长指数的拟合结果与张春蓓等[53]的描述较为吻合,全年发生4代左右。在过去生产活动中,灰茶尺蠖与茶尺蠖常被误认为是同一尺蠖物种。使用传统防治手段时,对两者的防治效果差异不大,直到近年在茶尺蠖核型多角体病毒[nuclear polyhedrosis virus(NPV)]的应用中发现其防效差异较大[54-55]。Zhang等[54]和席羽等[55-56]通过生物测定NPV对不同省份地区茶尺蠖幼虫的毒力,发现相差最大可达724.5倍;通过形态学鉴定、生物学特征观察及正反交试验,将其确定为两个物种。姜楠等[57]利用形态学和COI基因分析将其鉴定为灰茶尺蠖和茶尺蠖两个物种。由于之前的文献报道几乎未将这两个近缘种区分,本研究根据白家赫[5]和罗宗秀[6]的研究结果,以及收集的灰茶尺蠖分布资料、文献报道等,将浙江、江苏、安徽以外省(市、自治区)的茶尺蠖相关分布资料均作为灰茶尺蠖分布的参考资料,因此预测得到的结果可能存在一定的误差。
在灰茶尺蠖物种分布预测时,同时叠加寄主植物茶树的分布分析[46],可以更加准确地预测其分布范围。由于未能获得在中国可用的寄主植物茶树的实际分布范围,故在本研究中寄主植物茶树的分布并未考虑。但是目前茶树在中国大量的引种和种植,本研究所预测得到的灰茶尺蠖在中国的潜在分布范围,可以为目前尚未引种和种植以及已经引种和种植茶树但未发现灰茶尺蠖的地区提供参考依据,尤其是中高度适生区应加强监测,及时防范,以免造成巨大经济损失。后续可以深入研究灰茶尺蠖分布模型的敏感参数;充分考虑灰茶尺蠖寄主植物茶树及其天敌在中国的适生区分布和变化;结合最大熵模型(Maximum entropy model,MaxEnt)等其他预测模型,从而更好地为茶园科学管理和防控虫害等工作提供理论依据和技术支持。
致谢:感谢福建农林大学陈燕婷在论文撰写和修改及软件的实现过程中给予的帮助,感谢扬州大学车通在软件的实现过程中给予的帮助。
[1] 黄毅. 茶园环境和栽培措施对茶尺蠖和绒茧蜂的调控及绒茧蜂EAG研究[D]. 成都: 四川农业大学, 2009. Huang Y. Studies on regulation of tea garden environment and cultivation measures to populations ofandspp. and electroantennogram ofspp. [D]. Chengdu: Sichuan Agricultural University, 2009.
[2] 苏仁先. 尤溪县部分茶园茶尺蠖暴发原因与防治方法[J]. 蚕桑茶叶通讯, 2016(6): 35-36. Su R X. Outbreak reasons and control methods ofin partial tea plantations of Youxi County [J]. Newsletter of Sericulture and Tea, 2016(6): 35-36.
[3] Zhang G H, Yuan Z J, Yin K S, et al. Asymmetrical reproductive interference between two sibling species of tea looper:and[J]. Bulletin of Entomological Research, 2016: 1-8. doi: 10.1017/S0007485316000602.
[4] 唐美君, 郭华伟, 葛超美, 等.NPV对灰茶尺蠖的致病特性及高效毒株筛选[J]. 浙江农业学报, 2017, 29(10): 1686-1691. Tang M J, Guo H W, Ge C M, et al. Pathogenic characters ofnucleopolyhedroviruses onWarren and screening of high efficient strain [J]. Acta Agriculturae Zhejiangensis, 2017, 29(10): 1686-1691.
[5] 白家赫. 茶尺蠖两近缘种的生物学特性差异和分布研究[D]. 北京: 中国农业科学院, 2018. Bai J H. Study on differencial biological characteristics and geographical distribution of the sibling pests,and[D]. Beijing: Chinese Academy of Agricultural Sciences, 2018.
[6] 罗宗秀. 灰茶尺蠖和茶尺蠖性信息素鉴定及相关化学生态学研究[D]. 北京: 中国农业科学院, 2017. Luo Z X. Identification and correlative chemical ecology studies on sex pheromone ofand[D]. Beijing: Chinese Academy of Agricultural Sciences, 2017.
[7] 葛超美. 灰茶尺蠖的生物学特性及其体色遗传规律研究[D]. 北京: 中国农业科学院, 2016. Ge C M. Study on biological characteristics and genetic regularity of body color ofWarren [D]. Beijing: Chinese Academy of Agricultural Sciences, 2016.
[8] 薛东洋, 吴玉池, 曹立新, 等. 茶尺蠖的发生与气象条件的关系及其防治[J]. 现代农业科技, 2009(3): 146, 149. Xue D Y, Wu Y C, Cao L X, et al. Relationships betweenand meteorological condition and its prevention and control [J]. Modern Agricultural Science and Technology, 2009(3): 146, 149.
[9] 张辉, 李慧玲, 李良德, 等. 高温及覆土逆境对茶尺蠖蛹羽化的影响[J]. 茶叶学报, 2016, 57(2): 100-103. Zhang H, Li H L, Li L D, et al. Effect of temperature and soil coverage on pupae emergence ofProut [J]. Acta Tea Sinica, 2016, 57(2): 100-103.
[10] 董道青, 陈建明. 茶尺蠖蛹的耐冷藏性研究[J]. 中国计量学院学报, 2008, 19(2): 178-182. Dong D Q, Chen J M. Effects of cold storage on pupae of tea geometridae (Prout) [J]. Journal of China Jiliang University, 2008, 19(2): 178-182.
[11] 葛超美, 殷坤山, 唐美君, 等. 灰茶尺蠖发育起点温度和有效积温的研究[J]. 植物保护, 2016, 42(6): 110-112. Ge C M, Yin K S, Tang M J, et al. Developmental threshold temperature and effective accumulated temperature of[J]. Plant Protection, 2016, 42(6): 110-112.
[12] 楼云芬. 茶尺蠖发育历期的温度效应[J]. 茶叶科学, 1993, 13(2): 127-133. Lou Y F. Effect of temperature on the developmental duration of tea geometrid (Prout) [J]. Journal of Tea Science, 1993, 13(2): 127-133.
[13] 张汉鹄. 茶尺蠖发育起点温度与有效积温研究[J]. 安徽农业科学, 1989(1): 62-66. Zhang H H. Threshold for development and effective thermal summation ofWehrli (Lepidoptera: Lymantriidae) [J]. Journal of Anhui Agricultural Sciences, 1989(1): 62-66.
[14] 徐秀秀, 蔡晓明, 边磊, 等. 茶尺蠖潜在飞行能力的研究[J]. 茶叶学报, 2015, 56(4): 249-253. Xu X X, Cai X M, Bian L, et al. Flight capability of the tea geometrid,Prout (Lepidoptera: Geometridae) [J]. Tea Science and Technology, 2015, 56(4): 249-253.
[15] da Silva R S, Kumar L, Shabani F, et al. An analysis of sensitivity of CLIMEX parameters in mapping species potential distribution and the broad-scale changes observed with minor variations in parameters values: an investigation using open-fieldandas an example [J]. Theoretical and Applied Climatology, 2018, 132(1): 135-144.
[16] Chen Y T, Vasseur L, You M S. Potential distribution of the invasive loblolly pine mealybug,(Hemiptera: Pseudococcidae), in Asia under future climate change scenarios [J]. Climatic Change, 2017, 141(4): 719-732.
[17] Kumar S, Neven L G, Yee W L. Assessing the potential for establishment of western cherry fruit fly using ecological niche modeling [J]. Journal of Economic Entomology, 2014, 107(3): 1032-1044.
[18] Khormi H M, Kumar L. Climate change and the potential global distribution of: spatial modelling using geographical information system and CLIMEX [J]. Geospatial Health, 2014, 8(2): 405-415.
[19] Ramos R S, Kumar L, Shabani F, et al. Climate model for seasonal variation inusing CLIMEX in tomato crops [J]. International Journal of Biometeorology, 2019, 63(3): 281-291.
[20] Sutherst R W, Maywald G F, Bottomley W, et al. CLIMEX version 2. User’s guide. Hearne Scientific Software [M]. Melbourne: Hearne Scientific Software Pty Ltd., 2004.
[21] 白家赫, 王志博, 肖强. 浙江茶区茶尺蠖两近缘种的遗传分化及分布[J]. 昆虫学报, 2018, 61(6): 741-748. Bai J H, Wang Z B, Xiao Q. Genetic differentiation and distribution of two sibling species of tea geometrids in tea-growing areas in Zhejiang, eastern China [J]. Acta Entomologica Sinica, 2018, 61(6): 741-748.
[22] 夏英三. 茶尺蠖的生活习性与防治措施[J]. 福建茶叶, 1999(4): 12. Xia Y S. Living habits and prevention measures of[J]. Tea in Fujian, 1999(4): 12.
[23] 乔利, 洪枫, 张权, 等. 信阳市茶园灰茶尺蠖的抗药性研究[J]. 种业导刊, 2018(8): 12-14. Qiao L, Hong F, Zhang Q, et al. Resistance ofin tea plantations of Xinyang city [J]. Seed Industry Guide, 2018(8): 12-14.
[24] 赵丰华, 吕立哲, 党永超, 等. 茶尺蠖和茶细蛾性诱剂在豫南茶园的应用研究[J]. 天津农业科学, 2017, 23(1): 91-94.Zhao F H, Lyu L Z, Dang Y C, et al. Study on application ofandsex pheromone in southern Henan tea garden [J]. Tianjin Agricultural Sciences, 2017, 23(1): 91-94.
[25] 朱祚亮, 江书春, 张卜芬, 等. 茶尺蠖性信息素诱杀茶尺蠖试验[J]. 现代农业科技, 2018(9): 142, 144. Zhu Z L, Jiang S C, Zhang B F, et al. Control effects of sexual pheromone trap of[J]. Modern Agricultural Science and Technology, 2018(9): 142, 144.
[26] 谭荣荣, 刘明炎, 龚自明, 等. 湖北省茶区主要病虫害的种类及发生规律分析[J]. 茶叶通讯, 2013, 40(4): 36-38.Tan R R, Liu M Y, Gong Z M, et al. Investigation and analysis of diseases and insect pests in Hubei Province [J]. Tea Communication, 2013, 40(4): 36-38.
[27] 阮英东, 朱丽, 何涛. 房县茶园茶尺蠖危害特点及综合防控技术[J]. 湖北植保, 2018(3): 45-46. Ruan Y D, Zhu L, He T. Harmful characteristics and comprehensive control technology ofin tea plantations in Fang County [J]. Hubei Plant Protection, 2018(3): 45-46.
[28] 雷该翔, 王友平, 李新华, 等. 茶尺蠖发生特点及其绿色防控对策[J]. 湖北农业科学, 2018, 57(12): 51-53. Lei G X, Wang Y P, Li X H, et al. Occurrence characteristics and green prevention and control strategy of tea geometrid [J]. Hubei Agricultural Sciences, 2018, 57(12): 51-53.
[29] 张书平, 余燕, 李尚, 等. 天敌对白毫早茶园四种害虫空间跟随关系密切程度的年度间差异及其原因[J]. 应用昆虫学报, 2018, 55(4): 725-747. Zhang S P, Yu Y, Li S, et al. Spatial relationships among natural enemies and four insect pests over two consecutive years [J]. Chinese Journal of Applied Entomology, 2018, 55(4): 725-747.
[30] 孙钦玉, 张家侠, 罗仲兴, 等. 皖南茶区茶尺蠖发生情况及防治技术措施[J]. 中国植保导刊, 2014, 34(2): 38-40. Sun Q Y, Zhang J X, Luo Z X, et al. Occurrence and control measures ofin tea plantations in south Anhui province [J]. China Plant Protection, 2014, 34(2): 38-40.
[31] 涂海华, 邝先飞, 毛宇, 等. 双波段LED太阳能杀虫灯对茶园害虫的控制作用研究[J]. 中国植保导刊, 2018, 38(10): 53-57. Tu H H, Kuang X F, Mao Y, et al. Effect of solar insecticidal lamp with dual-band LED on insect pests in tea plantations [J]. China Plant Protection, 2018, 38(10): 53-57.
[32] 谢小群, 黎小萍, 杨普香, 等. 不同类型茶园主要益害生物的群落结构和消长规律分析[J]. 江西农业学报, 2018, 30(5): 65-67, 72. Xie X Q, Li X P, Yang P X, et al. Community structure and fluctuant law of main insect pests and their natural enemies in different types of tea plantations [J]. Acta Agriculturae Jiangxi, 2018, 30(5): 65-67, 72.
[33] 熊金龙. 杀虫灯在解决茶叶农药残留中的应用效果[J]. 江西农业, 2017(21): 23. Xiong J L. Application effect of insecticidal lamp in solving pesticide residues in tea [J]. Jiangxi Agriculture, 2017(21): 23.
[34] 李桂友, 王美兰. 博罗县茶叶种植过程中常见病虫害及防治[J]. 植物医生, 2018(6): 58-59. Li G Y, Wang M L. Common diseases and insect pests and control in tea plantations in Boluo County [J]. Plant Doctor, 2018(6): 58-59.
[35] 杨月策, 侯渊, 杨澜, 等. 100亿孢子/mL短稳杆菌悬浮剂防治茶尺蠖田间药效试验[J]. 现代农业科技, 2018(6): 99, 102. Yang Y C, Hou Y, Yang L, et al. Efficacy of10 billion spores/mL SC againstin tea plantations [J]. Modern Agricultural Sciences and Technology, 2018(6): 99, 102.
[36] 刘初生. 桂林毛尖茶园主要病虫害发生规律及防治技术[J]. 时代农机, 2018, 45(6): 26-27. Liu C S. Occurrence regularity and control technology of main diseases and insect pests of Guilin maojian tea garden [J]. Times Agricultural Machinery, 2018, 45(6): 26-27.
[37] 张方梅, 乔利, 潘鹏亮, 等. 灰茶尺蠖对三种非寄主植物精油的触角电位和行为反应[J]. 昆虫学报, 2018, 61(5): 565-573. Zhang F M, Qiao L, Pan P L, et al. Electroantennogram and behavioral responses of(Lepidoptera: Geometridae) to essential oils from three non-host plants [J]. Acta Entomologica Sinica, 2018, 61(5): 565-573.
[38] 姜星. 石阡苔茶主要病虫害绿色防控技术[J]. 植物医生, 2018(3): 47-48. Jiang X. Green control techniques of main diseases and insect pests of Shiqian Taicha [J]. Plant Doctor, 2018(3): 47-48.
[39] 段小凤, 徐小茜, 田景涛, 等. 松桃县灰茶尺蠖发生动态研究[J]. 南方农机, 2018, 49(6): 1-2, 5. Duan X F, Xu X Q, Tian J T, et al. Study on the population dynamics ofin Songtao County [J]. China Southern Agricultural Machinery, 2018, 49(6): 1-2, 5.
[40] 彭玉萍, 吴明耀, 罗宗秀, 等. 灰茶尺蠖性信息素诱杀效果试验[J]. 中国茶叶, 2018, 40(8): 30-31. Peng Y P, Wu M Y, Luo Z X, et al. Control effects of sexual pheromone trap of[J]. Chinese Tea, 2018, 40(8): 30-31.
[41] 罗学平. 绿色茶园病虫害系统控制[J]. 植物医生, 2017, 30(9): 51-53. Luo X P. System control on disease and insect pests of green ecological tea plantations [J]. Plant Doctor, 2017, 30(9): 51-53.
[42] 蒲德强, 毛建辉, 陈宇, 等. 2018年四川茶叶夏秋季病虫害防治技术要点[J]. 四川农业科技, 2018(6): 31-32. Pu D Q, Mao J H, Chen Y, et al. Key points of pest control techniques of Sichuan tea in the summer and autumn of 2018 [J]. Sichuan Agricultural Science and Technology, 2018(6): 31-32.
[43] 盛忠雷. 性信息素对茶园主要鳞翅目害虫控制的研究[D]. 重庆: 西南大学, 2012.
Sheng Z L. Research of the control for sex pheromones on main Lepidoptera pests in the tea garden [D]. Chongqing: Southwest University, 2012.
[44] 田忠正, 姜军侠, 李帅, 等. 陕南茶树病虫害发生现状与防治对策[J]. 陕西农业科学, 2016, 62(6): 71-74, 86. Tian Z Z, Jiang J X, Li S, et al. Damage situation and countermeasures of diseases and insect pests in tea plantations in southern Shaanxi [J]. Shaanxi Journal of Agricultural Sciences, 2016, 62(6): 71-74, 86.
[45] IPCC. Climate Change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change [R]. Cambridge: Cambridge University Press, 2007: 2-18.
[46] 张晓玲, 李亦超, 王芸芸, 等. 未来气候变化对不同国家茶适宜分布区的影响[J]. 生物多样性, 2019, 27(6): 595-606. Zhang X L, Li Y C, Wang Y Y, et al. Influence of future climate change in suitable habitats of tea in different countries [J]. Biodiversity Science, 2019, 27(6): 595-606.
[47] 杨海滨, 盛忠雷, 谢堃, 等. 不同栽培模式对山地茶园生态环境和茶叶品质的季节调控[J]. 西南农业学报, 2015, 28(4): 1559-1563. Yang H B, Sheng Z L, Xie K, et al. Seasonal regulation of different cultivation mode on ecological environment and tea quality in hilly tea plantation [J]. Southwest China Journal of Agricultural Sciences, 2015, 28(4): 1559-1563.
[48] 沈洁. 茶草复合生态系统的生态生理特性及草产量研究[D]. 合肥: 安徽农业大学, 2005. Shen J. Studies on ecological and biophysical traits and herby yields of tea-grass ecosystems [D]. Hefei: Anhui Agricultural University, 2005.
[49] 郭萧, 王晓庆, 彭萍, 等. 茶树不同成熟度叶片对茶尺蠖发育适合度的影响[J]. 茶叶科学, 2012, 32(3): 229-235. Guo X, Wang X Q, Peng P, et al. Developmental fitness ofProut feeding on tea leaves with different maturity [J]. Journal of Tea Science, 2012, 32(3): 229-235.
[50] 姚惠明, 周孝贵. 2016年秋季茶尺蠖暴发成因分析及防治启示[J]. 中国茶叶, 2016, 38(12): 21-22. Yao H M, Zhou X G. Causes analysis and control enlightenment ofoutbreak in autumn 2016 [J]. China Tea, 2016, 38(12): 21-22.
[51] 简文双. 南靖县茶尺蠖大发生原因初探及综合防治技术[J]. 东南园艺, 2015, 3(2): 66-68. Jian W S. A primary investigation of serious occurrence causes and comprehensive control techniques of tea geometrid in Nanjing County [J]. Southeast Horticulture, 2015, 3(2): 66-68.
[52] 高宇. 茶尺蠖天敌生态学研究进展[J]. 北方园艺, 2014(9): 203-206. Gao Y. Research progress on ecology of natural enemies of tea geometrid [J]. Northern Horticulture, 2014(9): 203-206.
[53] 张春蓓, 黄延政, 屈家新, 等. 夷陵西北山区茶园病虫害发生规律及绿色防控对策[J]. 中国茶叶, 2017, 39(6): 42-43.Zhang C B, Huang Y Z, Qu J X, et al. The rules of outbreak and green prevention and control strategies against the diseases and pests in tea plantations in southern regions of Yiling [J]. China Tea, 2017, 39(6): 42-43.
[54] Zhang G H, Yuan Z J, Zhang C X, et al. Detecting deep divergence in seventeen populations of tea geometrid (Prout) in China by COI mtDNA and cross-breeding [J]. Plos One, 2014, 9(6): e99373. doi: 10.1371/journal.pone.0099373.
[55] 席羽. 茶尺蠖地理种群对茶尺蠖核型多角体病毒的敏感性差异及遗传变异研究[D]. 北京: 中国农业科学院, 2011. Xi Y. Susceptibility variation againstnucleopolyhedrovirus and genetic variation in geographic populations of tea geometrid,Prout [D]. Beijing: Chinese Academy of Agricultural Sciences, 2011.
[56] 席羽, 殷坤山, 唐美君, 等. 浙江茶尺蠖地理种群已分化成为不同种[J]. 昆虫学报, 2014, 57(9): 1117-1122.Xi Y, Yin K S, Tang M J, et al. Geographic populations of the tea geometrid,(Lepidoptera: Geometridae) in Zhejiang, eastern China have differentiated into different species [J]. Acta Entomologica Sinica, 2014, 57(9): 1117-1122.
[57] 姜楠, 刘淑仙, 薛大勇, 等. 我国华东地区两种茶尺蛾的形态和分子鉴定[J]. 应用昆虫学报, 2014, 51(4): 987-1002. Jiang N, Liu S X, Xue D Y, et al. External morphology and molecular identification of two tea geometrid moth from southern China [J]. Chinese Journal of Applied Entomology, 2014, 51(4): 987-1002.
Potential Climate-suitable Distribution ofin China Based on the CLIMEX and ArcGIS Prediction
CHEN Lilin1,2, ZHOU Hao1,2, ZHAO Jie3
1. State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 2. Anxi College of Tea Science, Fujian Agriculture and Forestry University, Anxi 362406, China; 3. College of Natural Resources and Environment, Northwest Agriculture and Forestry University, Yangling 712100, China
Based on the data of 820 meteorological stations in China, known geographic distribution data and biological data, the current and potentialgeographic distribution of a major pestWarren in tea plantations were predicted using CLIMEX models and ArcGIS software. The potential effects of climate change on the future distribution ofwere also evaluated. The results suggest that the potential geographic distribution area was between 3°51′N and 40°6′N, which accounts for 34.27% of the total area of the country. The climatic conditions of most provinces and regions in China were suitable for the survival of. Due to climate change, the increasing rate of potential suitable area forgrew slowly, but its composition changed greatly. By 2050, the predicted proportion of highly suitable area reached a maximum of 22.23%. Compared
tea geometrid, climate change, suitable areas, potential distribution, prediction
S571.1;S435.711
A
1000-369X(2020)06-817-13
with A1B, the A2 scenario would accelerateextension in Shaanxi-Gansu-Ningxia region. Asare widely distributed in China, we suggested that monitoring measures should be improved and pest control should be taken as early as possible to ensure the safe production and quality of tea.
2019-09-03
2020-09-20
国家重点研发计划(2016YFD0200900)、国家自然科学基金项目(31501650)、福建农林大学茶产业链科技创新与服务体系建设项目(K1520005 A03)、福建省发改委农业“五新”工程项目(闽发改农业[2017]410号)、福建省区域发展项目(2017N3012)
陈李林,女,讲师,主要从事茶树病虫害生态控制和茶叶质量安全研究,llchen@fafu.edu.cn
