粳稻蔗糖转运蛋白基因OsSUTs在灌浆期的表达特点
2020-12-14吕东李丹丹徐汝聪
吕东 李丹丹 徐汝聪
摘要:蔗糖转运蛋白编码基因(SUTs)的表达在植物生长发育过程中的蔗糖转运与分配途径中起着至关重要的作用。由粳米生产的稻米具有良好的适口性,其市场需求量也越来越大,在稻米生产过程中,灌浆期是决定水稻产量和品质的重要时期。由此可见,研究粳稻灌浆期蔗糖转运蛋白编码基因(OsSUTs)的表达可为高产优质粳稻品种的选育和栽培提供有价值的参考。以云南农业大学稻作研究所保育的粳稻品系为材料,研究粳稻灌浆期叶片、穗部中OsSUTs基因的表达特点。结果表明,在粳稻叶片中,OsSUT2基因的表达量最高,而其余4个基因的表达量由高到低排序依次为OsSUT4、OsSUT1、OsSUT5、OsSUT3。穗部不同基因表达量的排序基本与叶片中的相同,由高到低为OsSUT2、OsSUT4、OsSUT5、OsSUT1、OsSUT3,不同的是OsSUT5与OsSUT1表达量的排序位置发生了对调。对比叶片、穗部中OsSUTs基因的表达量发现,除OsSUT1基因在穗部中的表达量低于在叶片中的表达量外,其他4个基因(OsSUT2、OsSUT3、OsSUT4、OsSUT5)的表达量均是穗中高于叶片中。在叶片中,OsSUT3、OsSUT4、OsSUT5这3个基因间的相关性较高,呈极显著正相关关系;但在穗中,OsSUTs基因间的相关性都较高,而且都达到了极显著正相关水平。研究结果可为人们深入认识水稻OsSUTs基因的表达特点及指导水稻高产优质育种生产提供理论参考。
关键词:蔗糖转运蛋白;基因表达;灌浆期;叶;穗;粳稻
中图分类号:S511.01 文献标志码: A 文章编号:1002-1302(2020)20-0056-06
水稻(Oryza sativa)是全球重要的糧食作物,世界上有一半的人口以水稻为主食[1-2]。随着国家稻米市场的开放及人民生活水平的提高,近年来人们对粳米的需求量与日俱增,国内粳米供需矛盾突出[3]。2018年,全球粳米消费量较2010年增长了约6900万t,增幅约为19%,全球粳米消费量占大米总消费量的比例由2010年的13%上升至2018年的14%。目前,我国仍为第一大粳米消费国,2018年的粳米消费量约为4200万t,较2010年增加了约400万t。因此,要保证粳米供给,必须扩大粳稻生产能力,突破粳稻品种选育滞后的问题,培育高产、抗逆的粳稻新品种[4]。高产优质粳稻品种的选育和生产离不开对粳稻遗传和生理代谢等各方面的深入研究。
蔗糖转运蛋白(sucrose transporter,简称SUT)是协调植物中碳素分配的重要分子,在植物各个生长阶段的能量分配中起着至关重要的作用[5]。多个研究发现,SUTs基因之间通过相互协作或相互抑制来共同调节植物的生理代谢及生长发育[6]。研究发现,沉默马铃薯(Solanum tuberosum)StSUT4基因,StSUT1基因呈现出过表达现象,甚至导致马铃薯花期提前[7];对拟南芥(Arabidopsis thaliana)的AtSUT1基因进行过表达发现,AtSUT2、AtSUT4基因的表达量相对下降[8]。此外,SUTs基因与植物的抗逆性也紧密相关。在对拟南芥AtSUT9基因进行沉默处理后,发现拟南芥植株对盐胁迫、渗透胁迫和低温胁迫的抵抗能力显著提高[9];对苹果(Malus domestica)的MaSUT2基因进行过表达后,发现植株对盐胁迫的抵抗力也明显增强[10]。
目前,研究者在水稻中共鉴定出5个蔗糖转运蛋白编码基因(OsSUT1、OsSUT2、OsSUT3、OsSUT4、OsSUT5)[11],其中对OsSUT1基因功能的研究较多,发现其在花粉发育、籽粒淀粉积累、灌浆速率、千粒质量等多个方面都起着非常重要的作用[12-13]。Miyazaki等研究发现,水稻耐热品种Genkitsukushi(暂无中文名称)在水稻抽穗期的茎、成熟期的种子中的OsSUT1基因表达量要明显高于其他热敏品种,说明此基因表达量的上调可以提高水稻对高温胁迫的抗性[14]。OsSUT2基因在水稻籽粒灌浆的过程中对有机物的转运起着重要作用,该基因突变会导致水稻农艺性状发生变化,如分蘖数减少,株高、千粒质量及根部干质量明显下降等,而其过表达会抑制OsSUT4基因的表达。此外,OsSUT2基因表达量的上调还可明显增强植株的抗旱和抗盐能力[15]。OsSUT3、OsSUT4和OsSUT5基因对颖果早期发育过程中的蔗糖转运起着一定的作用[11]。从表达模式上看,OsSUT3优先在水稻花粉中表达,因此推测其可能影响花粉发育过程中的淀粉积累[12]。OsSUT5基因在库组织中的表达量较高,与作物产量增加有关,抑制其表达会导致OsSUT1基因的表达量下调[16]。将OsSUT5基因转入马铃薯中发现,马铃薯产量提高了1.9倍多,说明此基因还可以调节植物果实的生长发育[17]。
灌浆期是水稻生殖生长的一个重要环节,然而目前关于粳稻灌浆期OsSUTs基因表达特点的报道较少。本研究以云南农业大学稻作所保育的部分粳稻品种(系)为材料,通过分析粳稻品种(系)灌浆期OsSUTs基因在叶片、穗中的表达模式,为选育高优、稳产的粳稻品种及水稻碳水化合物在源库流分配中的分子调控提供重要的参考价值。
1 材料与方法
1.1 试验材料
粳稻材料共30份,均由云南农业大学稻作所保育。试验材料种植于云南省宜良市云南农业大学稻作所试验田,采用大田常规管理,同时取样灌浆期的穗和绿色叶片,用液氮速冻后于-80 ℃保存备用。
1.2 试验方法
1.2.1 总RNA的提取 参照天根生化科技(北京)有限公司的RNA提取试剂盒说明书,用液氮预冷的镊子取100 mg水稻材料放入1.5 mL离心管中,并在离心管中加入液氮,用灭菌的研磨棒将材料充分磨碎。研磨完毕后立即加入1 mL TRIzol,并振荡混匀。将匀浆样品在室温下放置5 min,使核酸蛋白复合物完全分离,TRIzol、三氯甲烷的添加比例为1 mL ∶ 0.2 mL,盖好管盖后剧烈振荡15 s,室温放置3 min,然后于4 ℃、12 000 r/min离心 15 min;将上层无色液体转移到新的离心管中,加入1 mL 75%乙醇洗涤沉淀,然后于4 ℃、5 000 r/min离心 3 min。倒出上部液体,将剩余的少量液体于 5 000 r/min 离心10 s,然后用枪头吸出液体,将带有沉淀的离心管室温放置约2~3 min,再加入30~60 μL RNas-Free ddH2O,以充分溶解RNA。
1.2.2 cDNA的合成 根据TaKaRa逆转录试剂盒进行cDNA的合成。第1步,在RNase-free的无菌PCR管中加入4 μL H2O(RNase free)、2 μL 5×gDNA Eraser Buffer、1 μL gRNA Eraser和提取的 3 μL 总RNA,4 ℃、5 000 r/min离心5 s,于42 ℃恒温 2 min,然后立即轉移到冰上降温处理1 min;第2步,在新的RNase-free的PCR管中加入4 μL H2O(RNase free)、4 μL Prime Script Buffer、1 μL RT Prime Mix、1 μL Prime Script RT Enzyme Mix和10 μL 第1步的RNA混合液,然后将其置于37 ℃恒温处理15 min,随后于85 ℃处理5 s,取出后保存在-80 ℃,备用。
1.2.3 qRT-PCR 使用笔者所在课题组前期研究设计的5对OsSUTs基因的定量引物进行qRT-PCR试验,OsSUTs基因和内参β-actin基因引物均由生工生物工程(上海)股份有限公司合成。用SuperReal PreMix Plus[SYBR Green,天根生化科技(北京)有限公司]试剂盒及CFX96 Optical Reaction Module(美国伯乐公司)PCR仪进行实时定量PCR扩增。反应体系:12.5 μL 2×SuperReal PreMix Plus,1 μL cDNA,正、反向引物各1 μL,用RNase-free ddH2O补充总体积至25 μL。反应程序:95 ℃ 10 min;95 ℃ 15 s,61 ℃ 1 min,40个循环。每个样品独立重复3次。
1.3 数据处理
数据在Excel软件中采用2-△△CT法[18]进行分析,表达量数据图的绘制及相关性分析采用GraphPadPrism 8.0软件进行。
2 结果与分析
2.1 叶片和穗中OsSUTs基因在灌浆期的相对表达量
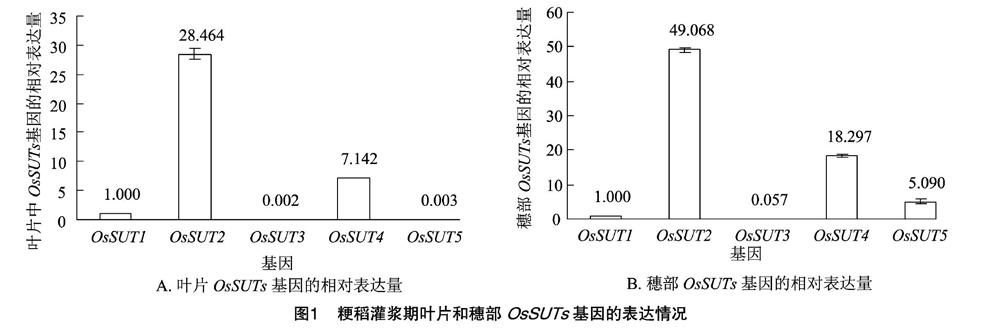
叶片的qRT-PCR结果表明,OsSUT2的相对表达量最高,达到28.464,其次为OsSUT4、OsSUT1、OsSUT5,OsSUT3的相对表达量最低,为0.002;与其他4个基因相比,OsSUT2的相对表达量是OsSUT1的28.464倍,OsSUT3的14 232倍,OsSUT4的3985倍,OsSUT5的9 488倍;OsSUT4的相对表达量虽居第2,但却明显高于OsSUT1、OsSUT3、OsSUT5,分别是它们的7.142、3 571、2 381倍;OsSUT1的相对表达量位列第3,是OsSUT3的500.0倍,OsSUT5的333.3倍;在5个基因中OsSUT5与OsSUT3的相对表达量最接近,OsSUT5的相对表达量仅为OsSUT3的1.5倍(图1-A)。
穗部OsSUTs的相对表达量最高的也为OsSUT2,达49.068,其次为OsSUT4,最低的是OsSUT3,相对表达量为0.057;与其他4个基因相比,OsSUT2的相对表达量是OsSUT1的49.068倍,OsSUT3的860.842倍,OsSUT4的2.682倍,OsSUT5的9.640倍;穗部OsSUT4的相对表达量也是排在第2位,为OsSUT1的18.297倍,OsSUT5的3595倍;OsSUT5的相对表达量也较高,为OsSUT1的5090倍,OsSUT3的89.298倍,OsSUT4的相对表达量仅为OsSUT5的3595倍;与叶片中相似,OsSUT3的相对表达量最低,且与其他4个基因之间的差异极大(图1-B)。
综上可见,OsSUT2的相对表达量在叶片、穗部均为最高,OsSUT1、OsSUT4的相对表达量在叶片、穗部也都相对较高,并且排序也较一致,均为OsSUT2 > OsSUT4 > OsSUT1。在检测的2个组织中,OsSUT3的相对表达量均为最低。有趣的是,OsSUT5在叶片中的相对表达量很低,但在穗部其相对表达量却较叶片中有所提高,接近OsSUT2的10%,变化最明显。
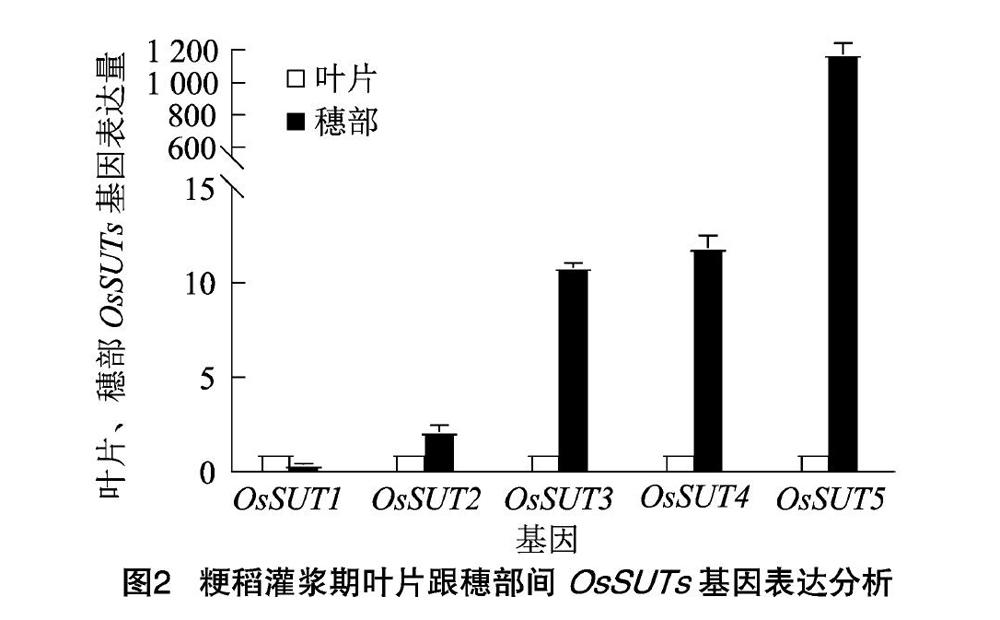
将叶片与穗部OsSUTs的相对表达量进行对比发现,由图2可知,只有OsSUT1的相对表达量表现为叶片中高于穗部,前者约比后者高2.4倍,叶片中其余4个基因的相对表达量均低于穗部;OsSUT5的相对表达量在叶片与穗部中的差异最大,穗部中的相对表达量是叶片中的1 176.44倍;OsSUT3和OsSUT4的相对表达量在叶片中与穗中的差异也极明显,穗部中OsSUT3的相对表达量是叶片中的108倍,穗部中OsSUT4的相对表达量是叶片中的11.9倍;OsSUT2在叶片中和穗部中的相对表达量相对稳定,穗部中OsSUT2的相对表达量较叶片中仅增加了1.15倍。
以上结果表明,在5个OsSUTs中,OsSUT1~4的表达量在叶片中和穗中差异明显,OsSUT5的相对表达量在叶片和穗部中的差异很大。
2.2 OsSUTs相对表达量的相关性分析
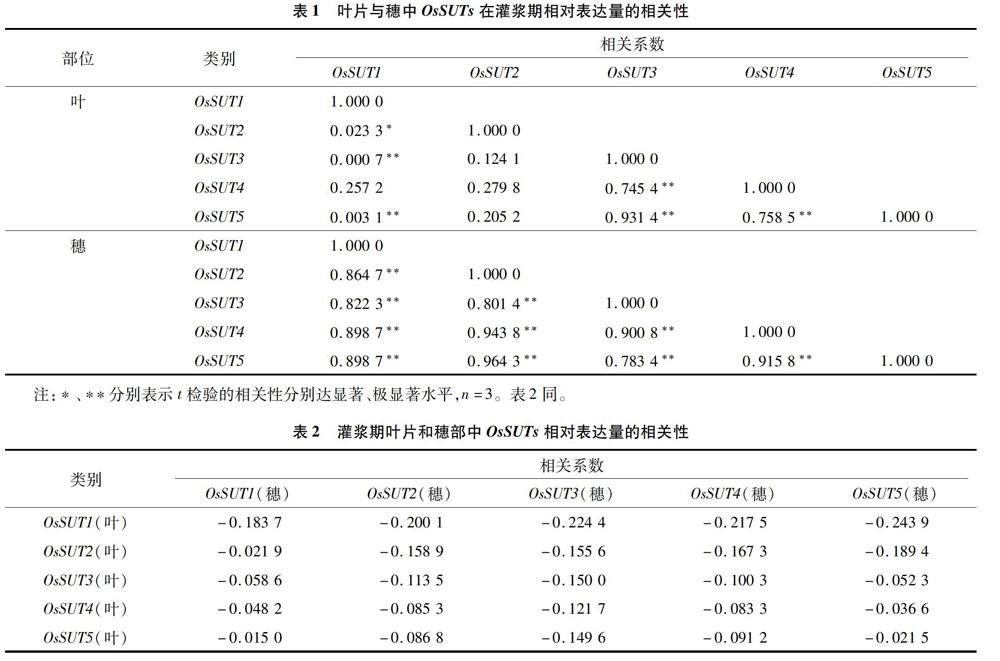
由表1可以看出,在叶片中,OsSUT1的相对表达量与OsSUT2的相对表达量呈显著正相关关系,与OsSUT3、OsSUT5的相对表达量呈极显著正相关关系,但与OsSUT4的相对表达量无显著相关性;
OsSUT2 的相对表达量只与 OsSUT1 的相对表达量呈显著正相关关系,与其他3个基因的相对表达量无显著相关性;OsSUT3的相对表达量与OsSUT1、OsSUT4、OsSUT5的相对表达量呈极显著相关关系,且与OsSUT5相对表达量的相关性最高,与OsSUT2的相对表达量无显著相关性;OsSUT4的相对表达量与OsSUT3、OsSUT5的相对表达量呈极显著正相关关系,与OsSUT1、OsSUT2的相对表达量无显著相关性;OsSUT5的相对表达量与OsSUT1、OsSUT3、OsSUT4的相对表达量呈极显著相关关系,但与OsSUT2的相对表达量无显著相关性。
由表1还可以看出,穗部OsSUT1的相对表达量与OsSUT2、OsSUT3、OsSUT4、OsSUT5的相对表达量呈极显著正相关关系;OsSUT2的相对表达量与OsSUT1、OsSUT3、OsSUT4、OsSUT5的相对表达量呈极显著正相关关系,且与OsSUT5相对表达量的相关性最高;OsSUT3的相对表达量与OsSUT1、OsSUT2、OsSUT4、OsSUT5的相對表达量呈极显著正相关关系;OsSUT4的相对表达量与OsSUT1、OsSUT2、OsSUT3、OsSUT5的相对表达量呈极显著正相关关系;OsSUT5的相对表达量与OsSUT1、OsSUT2、OsSUT3、OsSUT4的相对表达量呈极显著正相关关系。
综上可见,这5个OsSUTs在穗中的表达量相互间具有极显著的相关性,但是OsSUTs的相对表达量在叶片和穗部间的相关性不显著(表2)。
3 讨论与结论
3.1 灌浆期OsSUTs基因在叶片中的表达特点
植物体中蔗糖转运蛋白是具有蔗糖转运活性的跨膜结构蛋白,在植物生长发育、生理代谢、蔗糖的运输和分配过程中起着关键作用,还可以通过识别蔗糖信号影响其他代谢途径[19]。目前,人们从水稻中共鉴定出5个OsSUT基因,其中OsSUT1主要在韧皮部装载及同化物运输过程中起关键作用[20]。OsSUT1是负责运输蔗糖的关键基因,在籽粒灌浆期间,叶片产生的同化产物需要以蔗糖的形式通过韧皮部进行长距离运输才能到达穗部,并且长距离运输途径并不集中在叶部,而是在整个植株中,因此OsSUT1在叶片中的表达量不会太高。本研究中,OsSUT1在灌浆期叶片中的表达量在5个OsSUT基因中处于中间水平,也能说明OsSUT1在灌浆期的叶片中并没有较高表达量这一特点。OsSUT2主要在液泡膜上表达,并且在糖从源器官叶片输出到库器官的过程中扮演着重要角色,而在水稻灌浆期,叶片是光合作用的主要场所,其在灌浆期合成蔗糖的作用明显增大,使得叶肉液泡中的蔗糖含量显著增加[15]。本研究结果显示,OsSUT2在灌浆期叶片中的表达量最高,说明OsSUT2在光合同化物的运输和分配过程中起着举足轻重的作用。有研究发现,OsSUT3在开花后20 d会降低到几乎不能检测的水平[11]。本研究发现,OsSUT3在灌浆期叶片中的表达量极低,说明OsSUT3在水稻灌浆阶段的作用不明显。在水稻的库—源转化过程中,OsSUT4在叶鞘中有较高的表达量[21]。本研究中,OsSUT4在叶片中的表达量较高,也可能说明OsSUT4对于叶片中的蔗糖转运起着很重要的作用。有研究认为,OsSUT5主要参与蔗糖在组织和颖果中的卸载过程[22],而本研究发现,OsSUT5在叶片中相对表达量较低,说明OsSUT5发挥作用的主要场所不在叶片中。
3.2 灌浆期OsSUTs基因在穗中的表达特点
籽粒充实是水稻高产的重要保障。Aoki等研究OsSUT1发现,在水稻库组织中OsSUT1表达量很小[11],这与Hirose等研究发现的在开花后的穗中几乎检测不到OsSUT1的表达[21]相似,这与本研究发现的OsSUT1在穗中表达量相对较低的结果也一致。研究发现,OsSUT2在水稻各组织中呈表达水平稳定趋势[11],并通过其在花后籽粒的表达模式,推断出OsSUT2在籽粒的生长发育中起着主导作用,此外通过抑制OsSUT2的表达,从而造成源端光合作用合成的蔗糖不能及时有效地输送到库细胞中,从而造成源端大量积累蔗糖,籽粒蔗糖供应不充足,最终造成其生长迟缓,分蘖数、株高、千粒质量和根干质量显著降低,结实率低,结实不饱满及品质差等问题[15]。反之将OsSUT2进行过表达,发现其在抽穗后的表达量显著高于普通植株OsSUT2的表达量,并且在灌浆期中有效地提高了光合产物的转运和转换,进而显著地提高了水稻的单株粒质量及籽粒充实度[23]。这与本研究发现的叶与穗中OsSUT2的表达量都是稳定最高的结果相符,同时也表明了它对水稻产量确实有着重要的影响。他人研究发现,OsSUT3表达量有一定的波动性,在开花后1~2 d时达到最大,但在3 d时明显下降,5~7 d又恢复,然后逐渐下降[24-25]。本研究中,灌浆期穗中的OsSUT3表达量极低,这可能说明OsSUT3在灌浆期行使的功能场所并不在穗中;OsSUT4在颖果等组织中有较强的表达[11],这与本研究发现的OsSUT4在穗部的表达量较高的结果吻合。OsSUT5在花序及发育早期的颖果中能特异表达,其功能与籽粒的充实度和灌浆相关[26],本研究中OsSUT5的表达量显著增加与前人研究结果相同,这说明OsSUT5对水稻灌浆起着一定的作用。
3.3 叶穗间OsSUTs相对表达量的相关性
水稻叶片是蔗糖合成的重要产所,蔗糖从源到库的转运效率也尤为重要[27]。在水稻灌浆期间,叶片能为稻穗籽粒的充实提供合成有机物的源物质。因此,研究灌浆期叶和穗器官中OsSUTs基因表达的相关性可为探讨光合同化物运输、分配的代谢调控和水稻高产优质栽培提供理论参考。而在植物体的生长发育过程中,蔗糖转运蛋白基因会在植物的不同组织、不同时期中协同调控植物体的生长过程,因此蔗糖转运蛋白基因可能是单个特异的也可能是部分重叠的。本研究发现,多数OsSUTs基因的表达量在叶中不相关,这很可能跟OsSUTs特异性表达有关。比如OsSUT1与OsSUT5都是定位在质膜上的蔗糖转运蛋白,但OsSUT5的特异性又比OsSUT1差[28],与本研究相同的是OsSUT1与OsSUT5之间存在极显著正相关关系,但相关性并不明显。OsSUT2是定位在液泡中[15],而液泡作为贮存和积累能量的细胞器,其主要功能是调节细胞渗透压,因此OsSUT2与细胞膜上的OsSUT1、OsSUT5几乎是无相关性。OsSUT3与OsSUT4功能尚不明确,但本研究发现,OsSUT3、OsSUT4与OsSUT5都呈极显著相关关系且相关性较大,因此猜测OsSUT3与OsSUT4在叶片当中也起着一定作用,具体的机制有待下一步研究。
穗是水稻利用或貯存同化物的库,有研究认为库能刺激叶片中的光合活性,调节光合产物的分配[29]。基于OsSUTs基因在花后籽粒中的表达模式,可将籽粒发育分为2个阶段,第1个阶段为花后的1~4 d,为籽粒的生长及细胞分化阶段;第2阶段为花后5~15 d,为籽粒达到最长并开始增重阶段[30]。由此可推测出OsSUT2、OsSUT4和OsSUT5在第1阶段起主要作用,OsSUT1则在第2阶段起作用,而OsSUT3则起着衔接这2个阶段的作用[24]。与本研究的结果类似,在穗中,OsSUTs之间都呈极显著正相关关系,证明这5个基因共同协调灌浆期的籽粒发育过程。此外,也有人认为,在开花期0~14 d,在水稻中过表达OsSUT2基因,OsSUT4的表达量明显降低,为正常植株表达量的一半,表明OsSUT2基因的过表达可能会抑制OsSUT4的表达量[26]。而在本研究中,发现水稻灌浆期中的OsSUT2与OsSUT4基因呈正相关关系,这与上述结果相悖,猜测可能是植株本身会有一定的临界值,在过表达植株上,OsSUT2大量表达可能会适得其反,造成其他基因表达量降低以及影响植株生长发育。OsSUT3、OsSUT4与OsSUT5在穗中呈极显著正相关关系,且OsSUT5抑制表达会导致千粒质量降低,结实率降低[23]。由此猜测这几个基因都与水稻的经济产量有着紧密的关系。这也为OsSUT3与OsSUT4这2个基因功能的进一步研究提供了一定的参考价值。
叶片与穗部是水稻最主要的源与库器官。而流则是源和库之间的桥梁,流运行通畅会间接或者直接影响到源和库的生理活性。前人研究发现,转OsSUT2与OsSUT5会加快光合转运速率,理论上其最终的垩白度应该高于对照,但结果是垩白度低于对照[23]。从这里可以猜测,从源中将光合产物转运进库中不仅仅依靠蔗糖转运蛋白就能完成,在转运的过程中,还需要细胞壁转化酶[31-33]、单糖转运蛋白[34]等共同配合完成,并且在灌浆期中籽粒淀粉的生物合成过程中还有大量的酶与蛋白质参与。而本研究只涉及蔗糖转运蛋白基因,这也就说明了叶穗间OsSUTs并无相关性的原因。
综上,在本研究中,OsSUT2无论在叶和穗中表达量都显著高于其他4个基因,说明OsSUT2在灌浆期对蔗糖的转运分配起着重要的作用。由于5个OsSUTs基因在穗中的表达量呈极显著正相关关系,表明在颖果成熟阶段这5个基因是协同表达的。本研究可为蔗糖转运基因表达关系的进一步研究提供参考。
参考文献:
[1]程式华,胡培松. 中国水稻科技发展战略[J]. 中国水稻科学,2008,22(3):223-226.
[2]胡 锋. 保障我国粮食安全的水稻品种创新与应用研究[J]. 种子,2009,28(2):106-107,110.
[3]陈温福,潘文博,徐正进. 我国粳稻生产现状及发展趋势[J]. 沈阳农业大学学报,2006,37(6):801-805.
[4]胡兰香. 南方粳稻育种分析[J]. 南方农业,2018,12(18):180-181.
[5]Wang L,Lu Q T,Wen X G,et al. Enhanced sucrose loading improves rice yield by increasing grain size[J]. Plant Physiology,2015,169(4):2848-2862.
[6]Jung D L,Jung-Ⅱ C,Youn-Ⅱ P,et al. Sucrose transport from source to sink seeds in rice[J]. Physiologia Plantarum,2006,126(4):517-584.
[7]Izabela A,Chincinska,Johannes L,et al. Sucrose transporter StSUT4 from potato affects flowering,tuberization,and shade avoidance response[J]. Plant Physiology,2008,146(2):515-528.
[8]He H,Chincinska I,Hackel A et al. Phloem mobility and stability of sucrose transporter transcripts[J]. The Open Plant Science Journal,2008,2:1-14.
[9]Jia W Q,Zhang L J,Wu D,et al. Sucrose transporter AtSUC9 mediated by a low sucrose level is involved in arabidopsis abiotic stress resistance by regulating sucrose distribution and ABA accumulation[J]. Plant Physiology,2015,56(8):1574-1587.
[10]Ma Q J,Sun M H,Lu J,et al. Transcription factor AREB2 is involved in soluble sugar accumulation by activating sugar transporter and amylase genes[J]. Plant Physiology,2017,174(4):2348-2362.
[11]Aoki N,Hirose T,Scofield G N,et al. The sucrose transporter gene family in rice[J]. Plant & Cell Physiology,2003,44(3):223-232.
[12]Hirose T,Zhang Z J,Miyao A,et al. Disruption of a gene for rice sucrose transporter,OsSUT1,impairs pollen function but pollen maturation is unaffected[J]. J Exp Bot,2010,61(13):3639-3646.
[13]孙学武. 生育后期过表达OsSUT1基因对水稻碳水化合物与粒重的影响[D]. 长沙:中南大学隆平分院,2013:3-4.
[14]Miyazaki M,Araki M,Okamura K,et al. Assimilate translocation and expression of sucrose transporter,OsSUT1,contribute to high-performance ripening under heat stress in the heat-tolerant rice cultivar Genkitsukushi[J]. Journal of Plant Physiology,2013,170(18):1579-1584.
[15]Eom J S,Cho J I,Reinders A,et al. Impaired function of the tonoplast-localized sucrose transporter in rice,OsSUT2,limits the transport of vacuolar reserve sucrose and affects plant growth[J]. Plant Physiol,2011,157(1):109-119.
[16]张武君,管其龙,付艳萍,等. 反义抑制水稻蔗糖转运蛋白基因(OsSUT5)的表达降低其愈伤组织诱导和植株再生频率[J]. 农业生物技术学报,2014,22(7):825-831.
[17]Sun A J,Dai Y,Zhang X S,et al. A transgenic study on affecting potato tuber yield by expressing the rice sucrose transporter genes OsSUT5Z and OsSUT2M.[J]. Journal of Integrative Plant Biology,2011,53(7):586-595.
[18]Willems E,Leyns L,Vandesompele J. Standardization of real-time PCR gene expression data from independent biological replicates[J]. Analytical Biochemistry,2008,379(1):127-129.
[19]Schulze W X,Reinders A,Ward J,et al. Interactions between co-expressed Arabidopsis sucrose transporters in the split-ubiquitin system.[J]. BMC Biochemistry,2003,4:3.
[20]Barker L,Kühn C,Weise A et al. SUT2,a putative sucrose sensor in sieve elements[J]. Plant Cell,2000,12(7):1153-1164.
[21]Hirose T,Imaizumi N,Scofield G N et al. cDNA cloning and tissue specific expression of a gene for sucrose transporter from rice (Oryza sativa L.) [J]. Plant & Cell Physiology,1997,38(12):1389-1396.
[22]孫爱君. 水稻蔗糖转运蛋白基因的鉴定、表达分析及遗传转化[D]. 北京:中国科学院遗传与发育生物学研究所,2005:56-58.
[23]洪海强. 过表达OsSUT2和OsSUT5籼稻的灌浆生理[D]. 福州:福建农林大学,2008.
[24]Bush D R. Inhibitors of the proton-sucrose symport[J]. Bush,1993,307(2):355-360.
[25]Murata Y. Rice[C]//Evans L T. Crop physiolog. Cambridge:Cambridge University Press,1977.
