黔中地区马尾松-杜鹃植物群落多样性研究
2020-11-09于瀚欧静漆倩张智叶
于瀚 欧静 漆倩 张智叶


摘要:为研究黔中地区马尾松-杜鹃群落的物种组成及结构特点,并为人工营造松马尾松-杜鹃植物群落景观提供理论依据,采用典型样地调查法对群落进行调查。结果表明:研究地共记录植物50科78属91种,乔木层的建群种为马尾松,灌木层建群种为杜鹃,除此之外,分布较广的为禾本科、菊科、忍冬科、薔薇科植物。灌木层与草本层Simpson指数无显著差异,灌木层、草本层在Pielou均匀度指数、Margalef丰富度指数、Shannon-Wiener指数高于乔木层且差异显著,马尾松与杜鹃在乔灌层次中优势明显。马尾松-杜鹃植物群落乔木层中乔木种类较少,灌木层中物种较为丰富,整体呈现草本层>灌木层>乔木层趋势,样地之间存在明显差异。不同样地的马尾松-杜鹃群落的伴生植物存在差异,植物多样性在不同植物群落空间层次上有差异,草本层植物在数量和种类上占绝对优势,其次是灌木层。
关键词:马尾松;杜鹃;生物多样性;植物群落;黔中地区
中图分类号:S56723+6
文献标识码:A
文章编号:1008-0457(2020)02-0014-07国际DOI编码:10.15958/j.cnki.sdnyswxb.2020.02.002
Study on the Plant Community Diversity of Pinus massoniana-Azalea in Central Region of Guizhou
YU Han,OU Jing*,QI Qian,ZHANG Zhiye
(College of Forestry,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:The study aims to investigate the species composition and structural characteristics of Pinus massoniana-Azalea in central region of Guizhou Province by a typical plot survey method, which would provide the theoretical basis for artificially cultivated Pine-Azalea community landscape. The results showed that there area total of 91 species of 78 genera and 50 families of plants were recorded in the study area.The colony species in the arbor layer is P. massoniana and the shrub layer is constructed by Azalea. In addition, the plantswith wide distribution were Gramineae, Asteraceae, Lonicerae and Rosaceae plants.There was no significant difference in the Simpson index between the tree layer and the shrub layer.The shrub layer and herb layer in Pielou evenness index, Margalef richness index, Shannon-Wiener index were higher than the tree layer with significant difference. Pine.massoniana and Rhododendron had obvious advantages in the arbor and shrub hierarchy.There were fewer species in the trees layer of P.massoniana-
Rhododendron community, but more species in the shrub layer.The overall trend was herbaceous layer>shrub layer>tree layer.There were obvious differences between the plots.There were significant differences in the associated plants of P. massoniana-Azalea community,and the plant diversity varied at different spatial levels of plant communities.Herbal layer plants took an absolute advantage in terms of number and species,followed by shrub layer.
Keywords:Pinus massoniana;Azalea;biodiversity;plant community;central region of Guizhou
马尾松(Pinus massoniana Lamb.)是贵州省山林林分布最广泛的上层树种,普遍分布在贵州山地原生及次生林中,具有适应性强、速生丰产等优点[1]。杜鹃(Rhododendron simsii Planch.)为常见的马尾松下层伴生树种,多生于马尾松林下或林缘及山坡灌木丛[2],杜鹃对维持林木生产力具有重要意义[3]。相对稳定的复层混交植物群落物种多样性较高[46],而以马尾松为主的单优种群落多样性指数则较低[7],松纯林在乔木层中竞争力较差,同时,其群落结构也导致下层植物生长较差[89]。杜鹃群落对于灌木层植物的物种多样性和功能多样性呈显著相关性[10],在群落调查中,松林下多见杜鹃生长,并在下层灌木空间占有重要地位[1112]。同时包含松和杜鹃的植物群落,两者比一般的纯林长势更好[2],林下植物多样性更为丰富,林间生态系统更为稳定[13]。
本文以贵州省马尾松-杜鹃群落为研究对象,探究其物种多样性、群落物种组成及其种间关系、林下层级结构等内容,以马尾松-杜鹃为主的植物群落典型样地的群落类型、生态结构、物种多样性开展调查和系统研究分析,对营造生态、景观、经济多效益多功能的馬尾松-杜鹃植物群落具有重要意义。
1材料与方法
1.1研究区概况
选取贵州省黔中地区的马尾松-杜鹃植物群落进行调查,位于N26°26′~27°36′;E105°15′~107°57′。该区域的气候类型属中国亚热带高原季风湿润气候。年平均气温14~16 ℃,年平均降水量在1100~1300 mm之间,光照条件较差,年平均日照1200~1600 h左右。境内隶属云贵高原的喀斯特丘陵地貌,土壤多为微酸性红壤、黄壤,部分地区处于石漠化敏感地区,生态系统较为脆弱,需要保护部分山区,维护水源涵养、土壤保持、生物多样性等生态功能。黔中地区得益于气候
、地理等多方条件,植物种类繁多,植被群落组成与演替多样[14],具有明显的山地垂直自然带分布的植被类型,为植物群落发展提供了良好条件。
1.2样地概况及调查方法
1.2.1样地设置
2018年4-6月采样,基于群落外貌、地形地貌、群落伴生物种差异等三方面特征,调查样地为黔中地区7处马尾松-杜鹃典型群落,样地概况见表1。乔木群落样地设3处20 m×20 m样方,起测径阶1.5 cm,高度1.5 m2,对乔木采用每木调查法,记录种类、株数、高度、胸径、冠幅;采取系统抽样法在每个样方中选取3个5 m×5 m和2 m×2 m的小样方调查灌木及草本植物,记录灌木种类、株数、平均高度、盖度,草本植物种类、株数或丛数、平均高度和盖度;调查区内存在有间层植物,功能与灌木相似,故纳入灌木进行计算。同时记录经纬度、海拔、坡度、坡向。
1.2.2数据分析
重要值:表示某个种在群落中的地位与作用的综合数量指标,某一物种的重要值与该物种在群落中的优势程度成正相关[15],公式如下:
IV乔木层 =(相对多度+相对频度+相对显著度)×100% /3(1)
IV灌木层 =(相对多度+相对频度+相对显著度)×100% /3(2)
IV草本层 =(相对密度+相对频度+相对盖度)×100% /3(3)
物种多样性:根据α多样性的测度方法[17]分析,研究植物群落配置构成其公式如下:
丰富度指数:
R=S/lnA(4)
Shannon-Wiener指数:H'=-∑Pi ln Pi(5)
Simpson指数:D=1-∑(ni/N)^2(6)
均匀度指数:J=H'/ln S(7)
优势度指数:C=Nmax/N(8)
式中,重要值的取值范围为0~100;
A为单位面积;Pi是物种i的重要值,Pi=ni/N,N为所有物种的个体数之和,ni为第i个物种的个体数;S为物种总数;Nmax为优势种的个体数。
数据处理:采用Excel 2010对数据进行统计分析;利用SPSS 17.0进行群落物种多样性指数的相关性分析;运用Origin 7.5绘图。
2结果与分析
2.1群落植物组成及重要值分析
根据统计,黔中地区马尾松-杜鹃群落中共调查到植物91种,分属50科78属,其中蕨类植物6科6属8种,裸子植物2科2属2种,被子植物42科70属81种,被子植物中双子叶植物39科62属73种,单子叶植物3科8属8种。统计乔灌木共计25科35属45种,草本蕨类植物28科43属46种。
调查点单科多属种植物依次为蔷薇科(Rosaceae)(8属10种)和菊科(Compositae)(6属7种),其次为禾本科(Gramineae),壳斗科(Fagaceae),忍冬科(
Caprifoliaceae),杜鹃科(Ericaceae),百合科(Liliaceae)等,表明在马尾松-杜鹃植物群落中除建群种外,禾本科、菊科、忍冬科、蔷薇科植物为优势种,占各分布点总科属数的62.64%。藤黄科(Guttiferae)、玄参科(Scrophulariaceae)、楝科(Meliaceae)、杉科(Taxodiaceae Warming)、榆科(Ulmaceae)、五加科(Araliaceae)、胡颓子科(Elaeagnaceae)、葡萄科(Vitaceae)、紫草科(Boraginaceae)、唇形科(Labiatae)、兰科(Orchidaceae)、小檗科(Berberidaceae)木兰科(Magnoliaceae)、省沽油科(Staphyleaceae)等为单属单种植物,分布状况因样地海拔、气候、林间成熟度等因素而显示差异,占各分布点总科属数的37.36%。
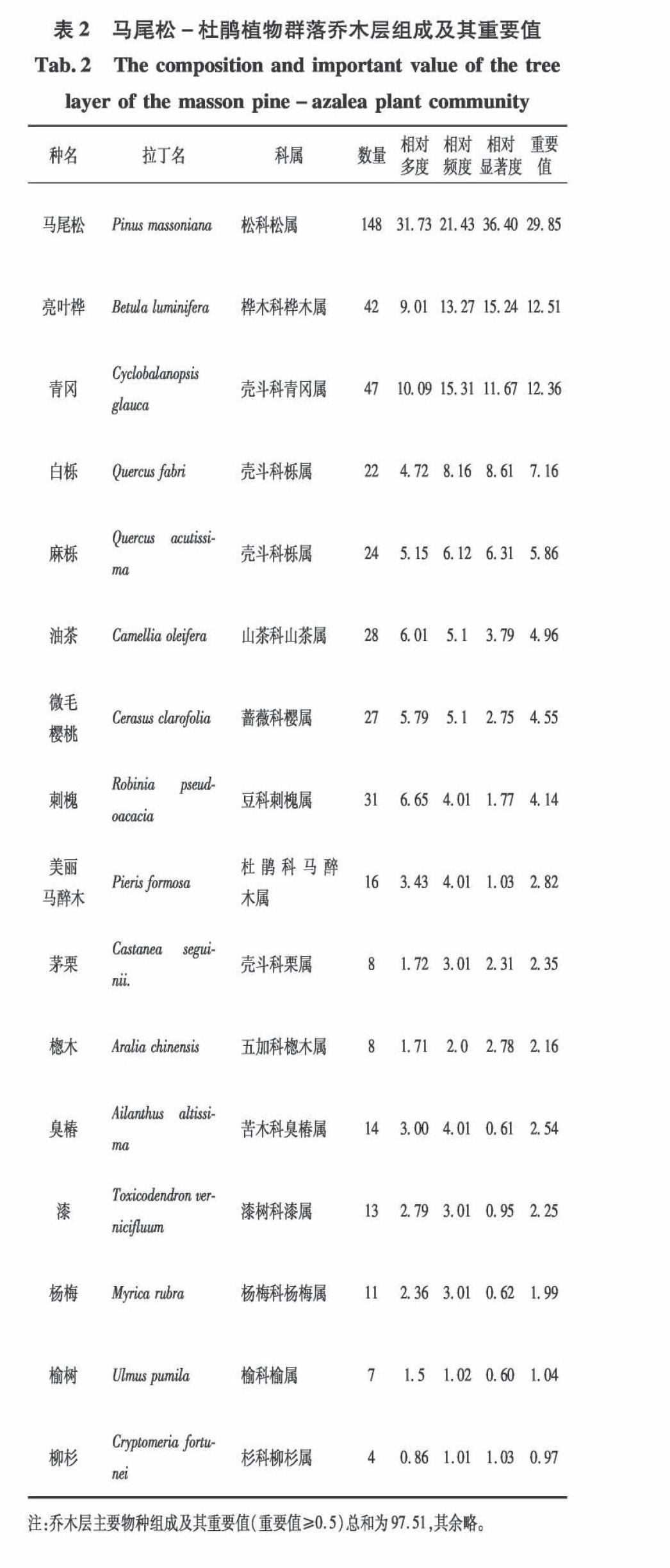
根据调查共统计乔木层21种,其中马尾松148株,因植株高大,生长竞争力强,为上层乔木建群种,其重要值为29.85
。剩余有较高权重的乔木有亮叶桦、油茶、青冈、白栎等,其重要值介于4.96~12.51之间,在不同地区样地出现频率较高
。其余种类如毛樱桃、杨梅、漆、杉树等在部分样地存在较广泛,区域差异较大,重要值介于2.25~4.55之间。马尾松-杜鹃林下灌木层共计25种,其中杜鹃127株,重要值为16.24,山茶、火棘、山莓、荚蒾类植物重要值分别在2.66~8.88之间,占较高权重。草本层中,五节芒、荩草、狗脊、白茅、紫箕占比重较大,重要值在4.65~13.25之间,为下层主要地被植物
,蕨类植物以西南凤尾蕨最为丰富,重要值9.06(表2~4)。
2.2植物多样性分析
多样性指数表示群落内种类多样性的程度,用来判断群落或生态系统的稳定性指标。群落多样性的测试值依赖样地的大小[1819]。黔中地区内不同样地的马尾松-杜鹃植物群落在植物多样性、丰富度上均存在差异,Shannon-Wiener指数和Margalef丰富度指数能够较好地表现出植物群落的差异[20]。结果表明,乔木层Margalef指数、Shannon-Wiener指数分别为2.453和0.763,低于灌木层的6.175和2.018,整体呈现草本层>灌木层>乔木层趋势,
LSD分析差异显著;Simpson指数乔灌草三层分别为0.541、0.740和0.751,群落间无明显差异;Pielou指数分别为0.551、0.639和0.817,草本层Pielou指数差异较大,且明显高于乔木层(表3)。
贵州黔中地区马尾松-杜鹃群落21处样地的植物多样性分析发现,除部分样地外,Margalef指数、Shannon-Wiener及Simpson指数在各地大多显示出相同趋势,即草本层>灌木层>乔木层。灌木层、草本层Pielou指数相近,乔木层Pielou指数普遍低于灌木层、草本层。同时,不同样地之间的差异性明显,同地区不同样方之间存在相似性。如乔木层Margalef指数在样地DF07、DF08中较低,而在样地MG15、MG17中较高;灌木层的Margalef指数在样地ZZ10、ZZ12中较低,而在样地QZ13、QZ14中较高;各层中Simpson指数在17号样地中最低,Pielou指数却最高;LL19、LL20、LL21与其他样地差异明显。由图1可以看出,马尾松-杜鹃7个分布群落丰富度指数与Shannon-Wiener指数趋势基本一致。
马尾松-杜鹃7个分布点马尾松、杜鹃的Simpson优势度指数均大于0.133,其中LL马尾松、杜鹃林间优势明显,为0.348和0.366;AS以马尾松优势度最低,为0.133;DF群落杜鹃优势度最低,为0.145。乔木层优势度普遍大于灌木层,马尾松既是建群种又是优势种;杜鹃优势种地位较马尾松低,下层群落多样性更加丰富。
3结论与讨论
根据各个样地植物调查研究结果可知,马尾松-杜鹃植物群落共有21种乔木,25种灌木,草本及蕨类46种,马尾松、杜鹃既是建群种又是优势种。群落物种多样性由于考虑了群落中不同物种的重要值,因而物种多样性能较好地反映群落的组成、结构和动态[21]。多样性指数除受物种数的影响外,还决定于物种个体数及其分布情况[22]。在本研究中,灌木层的Margalef物种丰富度指数、Shannon-Wiener指数和Simpson指数均明显高于乔木层,而灌木层与草本层Pielou均匀度指数相似并存在一定相关性,究其原因,可能是马尾松和杜鹃同时存在的群落环境中既有喜阴植物,又有喜阳植物,马尾松的林缘是一个过渡环境,种间关系复杂,其生长又极易受到外界环境的影响,所以造成Margalef丰富度指数、Shannon-Wiener指数和Simpson指数等较马尾松成熟林分高并呈现上述规律。
贵州喀斯特山地环境条件特殊,林分受环境及人为活动的影响较大[23],马尾松林缘的生境异质性大,因此与成熟马尾松群落的下层林分有所不同。马尾松-杜鹃群落在调查中多分布在阳坡,且马尾松林龄较小并未对下层空间造成直接影响,在没有明显影响层间郁闭度和产生严重的枯落物堆集的情况下,其下层植物表现出更强的竞争力;但是在修文县扎佐镇(ZZ)样地中成熟马尾松林的杜鹃仍长势良好,导致这一现象的原因可能是受群落自身生境因素的影响,马尾松与杜鹃地下部分着生不同生态功能的真菌形成特殊生境,增加地下层根系真菌类型多样性,同时影响森林生态系统的生物多样性[24],马尾松-杜鹃的菌根共生体提高了其对恶劣环境中汲取养分水分的能力[25],因而杜鹃在灌木层的特殊生态环境中对多样性与群落稳定和整個林分方面有促进作用。
马尾松-杜鹃林下群落空间与森林成熟形及林间郁闭度有相关关系,林分郁闭度过小和过大都对马尾
松天然更新产生显著影响[21,2627],从而影响植物群落稳定性与群落演替[28]。比较多地马尾松-杜鹃植物群落大致分为:幼年期,马尾松林中灌木层占主体,随着马尾松生长,郁闭度降低,喜光植物逐渐退至林缘,此时草本层灌木层较为丰富,杜鹃促进了马尾松的生长;到达群落成熟后期,林下光照不足、马尾松枯落物堆集,造成种子传播限制,凋落物、土壤理化性质等导致灌草层生存空间受限[29],中下层物种比例减少,
杜鹃和马尾
松对环境有相似的生态需求,有助于维持生态平衡与群落更替。
马尾松-杜鹃林不仅能够维持小区域生态稳定,还能够提供经济、社会等资源。为了更好的保护林间生态效应,提升景观效果,根据不同林龄和结构的植物群落,推荐补种喜阴阔叶植物并进行间伐处理[30],并拟建立永久样地,长期追踪马尾松-杜鹃植物群落动态发展,以便维持群落稳定,为后期继续探讨马尾松-杜鹃植物群落综合分析、配置模式以及群落景观评价等方面提供实践与理论基础。
参考文献:
[1]葛晓改,周本智,肖文发. 马尾松人工林凋落物产量、养分含量及养分归还量特性[J].长江流域资源与环境, 2014,23(7):9961003.
[2]林石狮,沈如江,郭微,等. 江西三清山台湾松+白豆杉-猴头杜鹃植物群落研究[J].生态环境, 2007,16(3): 912919.
[3]杨旭,丁炳扬,胡仁勇. 浙江杜鹃花属植物资源开发及利用[J].中国野生植物资源, 2005,24(2): 2225.
[4]陈昌雄,陈平留,肖才生,等. 人工马尾松复层混交林林分结构规律的研究[J].林业科学, 2001,37(S1): 205207.
[5]翁燕萍,魏绪英,陆金森,等. 南昌市象湖湿地花卉公园植物多样性调查与群落分析[J].江西农业学报, 2017, 29(5):
2227.
[6]朱建琴. 武夷山风景名胜区马尾松次生林群落数量特征变化研究[J].西南林业大学学报, 2014, 34(5): 1419.
[7]吴毅,周国英,祁承经,等. 湖南紫金山马尾松的群落特征及物种多样性分析[J].中南林业科技大学学报, 2011, 31(11): 120124.
[8]谢春平,南程慧,伊贤贵,等. 浙江安吉金钱松群落特征研究[J].植物资源与环境学报, 2018, 27(1): 9199.
[9]项小燕,吴甘霖,王志高,等. 大别山五针松群落物种多样性及区系分析[J].植物科学学报, 2017, 35(2): 243251.
[10]向琳,陈芳清,官守鹏,等. 井冈山鹿角杜鹃群落灌木层功能多样性及其随海拔梯度的变化[J].生态学报,
2019,39(21): 112.
[11]祝小科,黄艳斌,禹丹云,等. 黔东南国省干道公路边坡自然恢复植物群落物种多样性研究[J].山地农业生物学报, 2019, 38(4): 2934, 38.
[12]王厚麟,黄林生,缪绅裕,等. 广东石门台保护区猴头杜鹃群落特征[J].广西植物, 2008,14(1):7377.
[13]樊金会. 山东野生杜鹃花群落特性的调查[J].山东林业科技, 2000(2): 2527.
[14]李援越,朱守谦,祝小科. 黔中退化喀斯特森林植物群落的数量分类[J].山地农业生物学报, 2000,19(2): 9498.
[15]黄康有,廖文波,金建华,等. 海南岛吊罗山植物群落特征和物种多样性分析[J].生态环境, 2007,16(3): 900905.
[16]张钦弟,段晓梅,白玉芳,等. 山西太岳山脱皮榆群落的功能多样性[J].植物学报, 2016,51(2): 218225.
[17]马克平,刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J].生物多样性,1994,2(4): 231239.
[18]董冬,周志翔,何云核,等. 安徽省九华山风景区古树群落景观美学评价[J].生态学杂志, 2011, 30(8): 17861792.
[19]董安强,梁耀,胡明锋,等. 广东南岭国家级自然保护区南华杜鹃群落学研究[J].安徽农业科学, 2012, 40(21):
1092910933.
[20]胡理乐,闫伯前,江明喜,等. 毛柄小勾儿茶伴生群落种类组成及多样性研究[J].西北植物学报,2007,27(3): 594600.
[21]严兆贤. 马尾松天然更新的特点及制约成林因素[J].现代农业科技, 2019(10): 123.
[22]易好,邓湘雯,项文化,等. 湘中丘陵区南酸枣阔叶林群落特征及群落更新[J].生态学报, 2014, 34(12): 34633471.
[23]盘金文,郭其强,余大龙,等. 不同林龄马尾松人工林优势种径级结构及物种多样性分析[J].西北植物学报, 2019, 39(4): 722728.
[24]熊丹,欧静,李林盼,等. 黔中地区马尾松林下杜鹃根部内生真菌群落组成及其生态功能分析[J].生态学报, 2020(4): 112.
[25]欧静,韦小丽,何跃军,等. 接种ERM真菌对桃叶杜鹃幼苗的促生效应及生理生化影响[J].林业科学, 2013, 49(7): 4856.
[26]崔宁洁,刘洋,张健,等. 林窗对马尾松人工林植物多样性的影响[J].应用与环境生物学报, 2014, 20(1): 814.
[27]吴庆锥. 福建明溪丝栗栲优势群落的結构稳定性[J].亚热带农业研究, 2019, 15(1): 16.
[28]秦随涛,龙翠玲,吴邦利. 茂兰喀斯特森林不同地形部位优势乔木种群的生态位研究[J].广西植物, 2019, 39(5):
681689.
[29]陈小红,赵安玖,张健,等. 不同林龄马尾松林下植物多样性与环境特征[J].四川农业大学学报, 2017, 35(2): 186
192, 233.
[30]殷沙,赵芳,欧阳勋志. 马尾松木荷不同比例混交林枯落物和土壤持水性能比较分析[J].江西农业大学学报, 2015, 37(3): 454460.
