新疆13种荒漠植物根内生放线菌多样性及其抗菌活性筛选
2020-10-29雷艳娟夏占峰张利莉
雷艳娟 夏占峰 曾 红 张利莉
(塔里木大学生命科学学院/新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,新疆 阿拉尔 843300)
植物内生菌是指在其生活史的一定阶段或全部阶段存活于健康植物组织内部或细胞间隙,而不使宿主植物表现出明显感染症状的微生物,包括细菌、真菌和放线菌[1]。受温度、湿度、土壤情况等生长环境因素的影响,即便同种植物也会因地理位置的不同,表现出内生菌多样性分布的差异。云南大学微生物研究所陈华红等[2]从云南不同地点采集药用植物分离内生放线菌,其种类、数量有很大差异,表明了宿主的地域多样性在一定程度上决定了内生放线菌的多样性。来自植物内生菌的化合物约51%是新化合物[3],而来自土壤微生物的只有38%是新化合物,因此植物内生菌的研究备受关注。不同地域和种类的植物提供了内生菌生境的多样性,拓宽了筛选活性物质的途径。目前,在真菌、细菌、放线菌等植物内生菌中,因放线菌生长缓慢、分离困难而研究相对较少。植物内生放线菌研究也主要集中于药用植物、雨林植物等陆地植物和红树林等海洋植物,荒漠植物内生放线菌是尚未充分开发的新领域,值得探索研究。
塔里木盆地处于天山、昆仑山和阿尔金山之间,边缘是与山地相连的砾石戈壁,中心是塔克拉玛干沙漠,受地形气候影响,盆地边缘和沙漠之间的土壤干旱、盐渍化严重;孕育了很多具有防风固沙作用的荒漠盐生植物,如盐生灌木柽柳、盐穗木、骆驼刺等,它们适应干旱高盐生境,具有较强的耐旱抗盐能力[4]。干旱生境下的微生物为了能在持续缺水和高盐等严酷条件下生长,就不得不在生理和分子方面进化[5],在这样极端环境下生长的植物,其内生放线菌,伴随宿主植物长期适应性协同进化,除了产生新的基因型,还可能产生新型次级代谢产物[6]。感染性疾病是目前临床上最常见的疾病,在人类疾病方面,金黄色葡萄球菌所致的皮肤软组织感染,大肠杆菌所致的泌尿生殖道感染及白色念珠菌所致的念珠菌性阴道炎较常见;此外,金黄色葡萄球菌和大肠杆菌对家畜、家禽及其他经济动物也是易感染的,如奶牛乳房炎和肉鸡大肠杆菌病[7-9]。金黄色葡萄球菌、大肠杆菌和白色念珠菌这三种病原菌分别作为革兰阳性菌、革兰阴性菌和真菌的模式菌株研究具有代表意义。在植物病害方面,棉花枯萎病和黄萎病以及红枣黑斑病已成为新疆两大经济作物的最大病害。黄萎病导致棉花经济损失15%以上,红枣黑斑病严重时减产50%[10-11],因此迫切需要寻找能够抵抗人类或植物病害的新型活性天然产物。为了更好保护和利用塔里木盆地微生物资源,本研究以13种荒漠植物为材料,分离其内生放线菌,探索新疆典型荒漠植物内生放线菌的群落结构;以人体病原菌金黄色葡萄球菌、大肠杆菌、白色念珠菌,红枣黑斑病致病菌细极链格孢菌、棉花黄萎病致病菌大丽轮枝菌、棉花枯萎病致病菌尖孢镰刀菌为靶标菌,筛选活性菌株,以期寻找新型抗菌活性化合物的生产菌株,为生物医药和生物肥料提供菌株来源。
1 材料与方法
1.1 供试材料
2018年7月至8月在新疆天山山脉戈壁和盐碱地采集柽柳(Tamarix chinensisLour.)、碱蓬(Suaeda glauca(Bunge)Bunge)、河西菊(Launaea polydichotoma(Ostenf.)Amin ex N.Kilian)、盐穗木(Halostachys caspica(Bieb.)C.A.Mey.)、盐爪爪(Kalidium foliatum(Pall.) Moq.)、盐 节 木(Halocnemum strobilaceum(Pall.)M.Bieb.)、盐生草(Halogeton glomeratus(M.Bieb.)Ledeb.)、铃铛刺(Halimodendron halodendron(Pall.)Voss)、骆驼刺(Alhagi sparsifoliaShap.)、耳叶补血草(Limonium otolepis(Schrenk)Ktze.)、老鹳草(Geranium wilfordiiMaxim.)、西伯利亚铁线莲(Clematis sibirica(L.)Mill.)和新疆猪毛菜(Salsola sinkiangensisA.J.Li)13种植物样品,样品装于自封袋中,带回实验室,置于4℃冰箱保存备用。
1.2 供试靶标菌
以革兰阳性细菌代表种金黄色葡萄球菌(Staphyloccocus aureus,S.a)敏感株 ATCC 25923、革兰阴性细菌代表种大肠杆菌(Escherichia coli,E.c)敏感株ATCC 25922、酵母菌代表种白色念珠菌(Candida albicans,C.a)ATCC 64550以及植物病原菌大丽轮枝菌(Verticillium dahliae,V.d)ACCC 36916、尖孢镰刀菌(Fusarium oxysporum,F.o)ACCC 372587和细极链格孢菌(Alternaria tenuissima,A.t)为靶标菌进行活性放线菌的筛选。
1.3 培养基
(1)分离培养基:SCA培养基。
(2)纯化及斜面培养基:高氏一号培养基。
(3)活性筛选培养基:革兰阳性细菌和革兰阴性细菌培养选用美国临床和实验室标准协会(CLSI)中的MHA(MHB)培养基培养。白色念珠菌选用SDA(SDB)培养基培养;大丽轮枝菌、尖孢镰刀菌选用PDA(PDB)培养基培养;细极链格孢菌选用OA(OB)培养基培养。
1.4 化学试剂
(1)抑制剂:重铬酸钾、萘啶酮酸、放线菌酮、制霉菌素终浓度均为50 mg/L,待培养基温度降至40~50℃时加入。
(2)PBS缓冲液(g/L):NaCl 8.0,KCl 0.2,Na2HPO41.4,KH2PO40.27。
1.5 植物内生菌的分离、保藏和鉴定
参考李飞娜等[12]实验方法,将植物根进行表面消毒、研磨,磨碎的植物组织用PBS缓冲液混匀,涂布于SCA培养基,每种样品设置三组平行实验。最后一次清洗样品的无菌水取100 μL涂布于SCA培养基,28℃倒置培养1周,检测植物表面消毒效果。没有微生物生长,说明植物表面消毒彻底,分离到的放线菌为植物内生放线菌。对放线菌采用三区划线法进行纯化,直至得到纯菌株。以20%(体积比)甘油作为保护剂,将菌株接种于其中,置于-80℃保藏。
采用酶解法提取放线菌基因组DNA,扩增16S rRNA基因序列,送往北京睿博兴科测序公司测序。登录数据库EzBioCloud[13](http://www.ezbiocloud.net),对16S rRNA基因序列进行相似性比对,从中选取相似性较高的典型菌株16S rRNA基因序列作为参比对象,采用MEGA 7.0软件以邻接法(neighbor-joining)进行聚类分析,并构建系统进化树[14],根据序列比对结果和系统发育分析,确定菌株的种类。
1.6 抗菌活性筛选
将待检测的放线菌菌株接种于高氏固体培养基,28℃培养7天,制为直径7 mm的菌饼备用。对于细菌,采用琼脂扩散法,将靶标菌单菌落接种至10 mL MHB培养基,37℃,180 rpm摇床培养8 h,取100 μL涂布于MHA培养基,接种放线菌菌饼,置于培养箱中孵育18 h。对于真菌,靶标菌为白色念珠菌时,拮抗活性的检测方法与细菌相同,只需将培养基换为SDA;靶标菌为大丽轮枝菌、尖孢镰刀菌时,采用平板对峙法稍做修改,将PDA培养基上靶标菌接种至10 mL PDB培养基,20℃,180 rpm摇床培养72 h,取100 μL涂布于PDA培养基,接种放线菌菌饼,置于20℃培养箱中孵育48 h;靶标菌为细极链格孢菌时,将OA培养基上细极链格孢菌的孢子,用无菌棉签收集至无菌水中,取100 μL涂布于OA培养基,接种放线菌菌饼,置于28℃培养箱中孵育48 h。各设置平行实验3组,计算抑菌圈直径。
2 结果与分析
2.1 13种荒漠植物根内生放线菌多样性分析
通过SCA分离培养基从13种植物根分离出533株放线菌,根据菌落特征初步排除重复后,选取361株提取基因组DNA,进行16S rRNA基因序列扩增和测序,通过EZbiocloud进行比对分析。结果显示,361株内生放线菌,分属于放线菌纲的6个目7个科9个属77个种,这9个属分别是链霉菌属(Streptomyces)、拟诺卡氏菌属(Nocardiopsis)、拟无枝酸菌属(Amycolatopsis)、诺卡氏菌属(Nocardia)、冢村氏菌属(Tsukamurella)、糖霉菌属(Glycomyces)、链孢囊菌属(Streptosporangium)、伦茨菌属(Lentzea)、北里孢菌属(Kitasatospora)。
13种荒漠植物根内生放线菌群落结构结果见表1。由表1可以看出,不同荒漠植物内生放线菌多样性有差异。从分离到的菌株数量上看差异较大,老鹳草最多,依次为骆驼刺、铃铛刺、红柳、补血草、碱蓬、铁线莲、盐爪爪、河西菊、猪毛菜、盐节木、盐生草和盐穗木。从种属水平看差异也较大,骆驼刺内生放线菌最丰富,鉴定到6个属27个种,链霉菌属有20个种(图1);碱蓬次之,鉴定到4个属9个种,链霉菌属有6个种;S.griseoincarnatus为骆驼刺和碱蓬内生放线菌中分离频率最高的优势种;柽柳鉴定到3个属17个种,链霉菌属有15个种,S.plicatus为优势种;老鹳草分离到的菌株数量最多,归属于3个属,S.pactum为优势种;铃铛刺鉴定到2个属,其中的链霉菌属有28个种,S.ambofaciens优势种;猪毛菜、盐穗木等鉴定到链霉菌属。此外,16S rRNA基因序列比对结果显示,361株内生放线菌中有3株相似性小于98.65%,为潜在新种,与3个已有效发表的放线菌物种相似性最高,见表2。在骆驼刺、柽柳和盐爪爪各1株,均属于链霉菌的潜在新物种,具体分类地位正在进行多相分类学研究确认。
由表1和表3可以看出,本研究荒漠植物内生放线菌中,链霉菌为优势属,在13种植物中均有分布。此外,获得8个稀有放线菌属,拟诺卡氏菌属在8种植物中有分布,拟无枝酸菌属在5种植物中有分布,诺卡氏菌属在3种植物中有分布,糖霉菌属、链孢囊菌属和伦茨菌属仅在骆驼刺中分离得到,而北里孢菌属仅在老鹳草分离到,冢村氏菌属也仅在盐生草中分离到。表明放线菌的链霉菌可在植物中广泛分布,拟诺卡氏菌属、拟无枝酸菌属可能在植物中选择性分布,还有一些放线菌对宿主植物选择具有特异性。拟诺卡氏菌属是稀有放线菌中产新型天然活性化合物最多的,拟无枝酸菌属仅次于小单孢菌属位列第三[15],这类型菌株产生的化合物具有广泛的生物学活性,在医药方面作为抗菌、抗过敏、抗肿瘤、降血脂、抗肿瘤等药物有很好的疗效。本研究中,分离得到的拟诺卡氏菌属和拟无枝酸菌属,为后续抗菌活性物质的筛选提供了更大的可能性。经文献调研,冢村氏菌属与伦茨菌属为首次从植物中分离到。
2.2 荒漠植物内生放线菌抗菌活性筛选
361株荒漠植物根内生放线菌的抗菌活性筛选结果见表4。结果显示有80株活性放线菌,占鉴定总数的22%,其中链霉菌74株,拟无枝酸菌3株,拟诺卡氏菌、糖霉菌和链孢囊菌各1株。
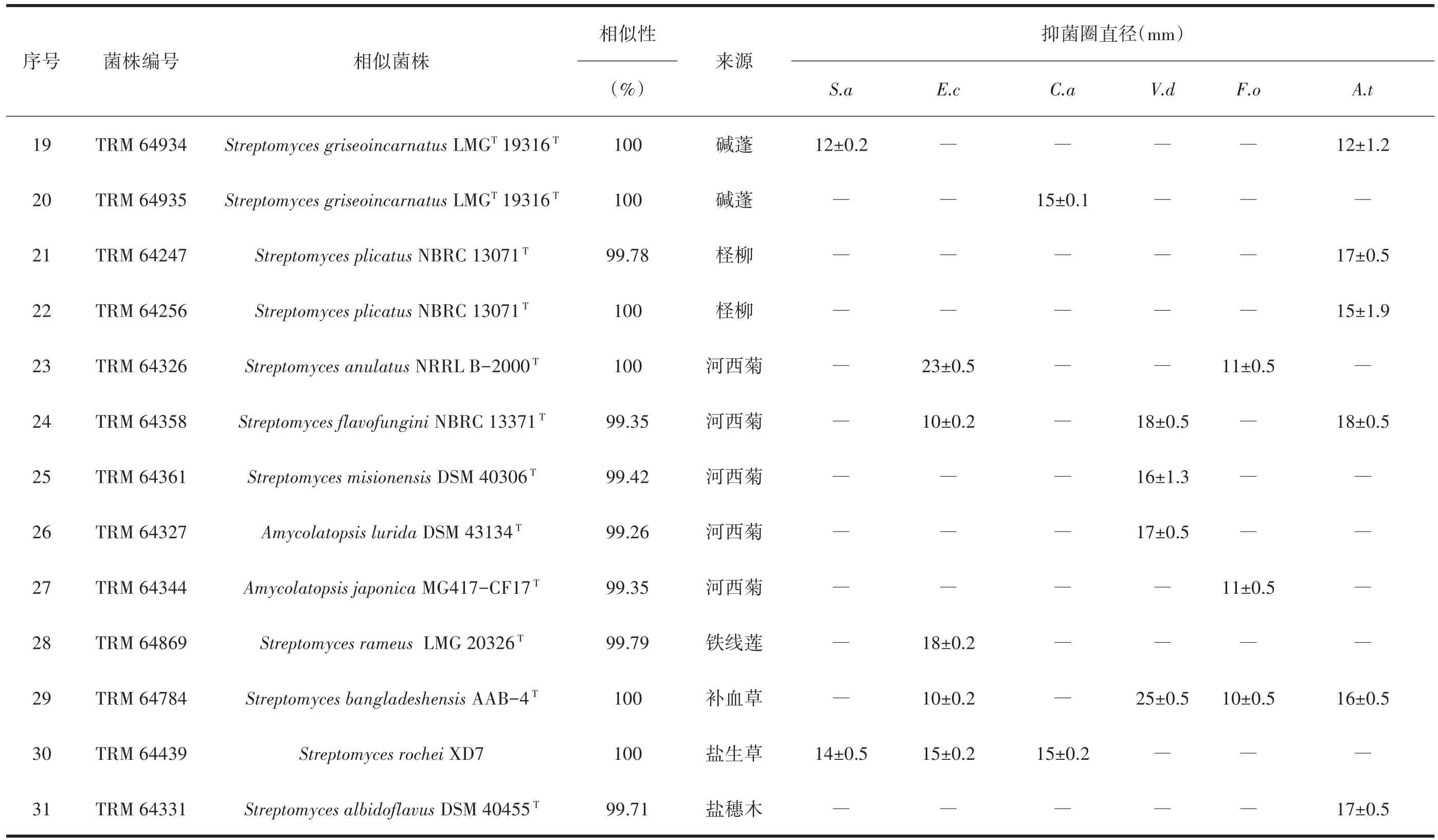
铃铛刺内生放线菌中,仅链霉菌属有活性菌株15株;骆驼刺内生放线菌中,活性菌株共有12株,链霉菌有9株;拟诺卡氏菌属、糖霉菌属、链孢囊菌属中的活性菌株各为一株,链孢囊菌和糖霉菌为稀有放线菌中少见的有抑菌活性的菌株,链孢囊菌Streptosporangiumamethystogenessubsp.amethystogenes(TRM 64968)对白色念珠菌有抑制作用,糖霉菌Glycomyces algeriensis(TRM 65112)对大肠杆菌和白色念珠菌有抑制;拟诺卡氏菌属中Nocardiopsis dassonvilleisubsp.dassonvillei(TRM 64374)对大丽轮枝菌有抑制。骆驼刺中,TRM 64368、TRM 64461、TRM 65026三株放线菌均与S.griseoincarnatus的16S rRNA基因序列相似性为100%,抗菌谱却不同;此外,碱蓬中TRM 64934和TRM 64935也与S.griseoincarnatus16S rRNA基因序列相似性为100%,不仅抗菌谱不同,抗菌强度也不一样,见表5。

图1 基于骆驼刺根内生放线菌16S rRNA基因序列构建的neighbour-joining系统进化树

表2 13种荒漠植物根内生放线菌中的潜在新物种

表3 361株内生放线菌的多样性
对于不同靶标病原菌,拮抗大肠杆菌的活性放线菌有39株,拮抗链格孢菌的活性放线菌有35株,拮抗金黄色葡萄球菌的活性放线菌有31株,拮抗白色念珠菌的活性放线菌有30株,拮抗尖孢镰刀菌的活性放线菌有17株,拮抗大丽轮枝菌的活性放线菌有6株。铃铛刺内生放线菌S.rectiviolaceus(TRM 64937)对大肠杆菌、尖孢镰刀菌、白色念珠菌和细极链格孢菌均有抑制作用,该菌株是13种植物中对尖孢镰刀菌抑制作用最强的菌株,抑菌圈直径为 15 mm;S.albidoflavus(TRM 64433)对白色念珠菌抑制作用最强,抑菌圈直径为16 mm。骆驼刺内生放线菌中S.variabilis(TRM 64369)对链格孢菌的抑制作用最强,抑菌圈直径达到30 mm;S.griseoincarnatus(TRM 65060)对金黄色葡萄球菌抑制作用最强,抑菌圈直径达到21 mm。补血草内生放线菌S.bangladeshensis(TRM 64784)对大丽轮枝菌抑菌活性最强,抑菌圈直径25 mm。河西菊内生放线菌S.anulatus(TRM 64326)对大肠杆菌抑菌活性最强,抑菌圈直径23 mm。
对3种病原菌均有抑菌作用的有S.rectiviolaceus(TRM 64937)、S.rochei(TRM 64439)、S.pactum(TRM 64989)、S.griseoincarnatus(TRM 65060)、S.griseorubens(TRM 64343),后三者是13种植物内生放线菌中链霉菌属分离频率最高的物种,S.griseoincarnatus和S.griseorubens在13种荒漠植物中的柽柳、猪毛菜、补血草、碱蓬、骆驼刺都有分布。自河西菊中筛选出的放线菌活性菌株比率最高(河西菊活性菌株数量/河西菊分离菌株数量),结合抑菌活性强度、活性菌株的数量与多样性,骆驼刺和河西菊是13种荒漠植物内生放线菌中适合抗菌活性筛选的植物。
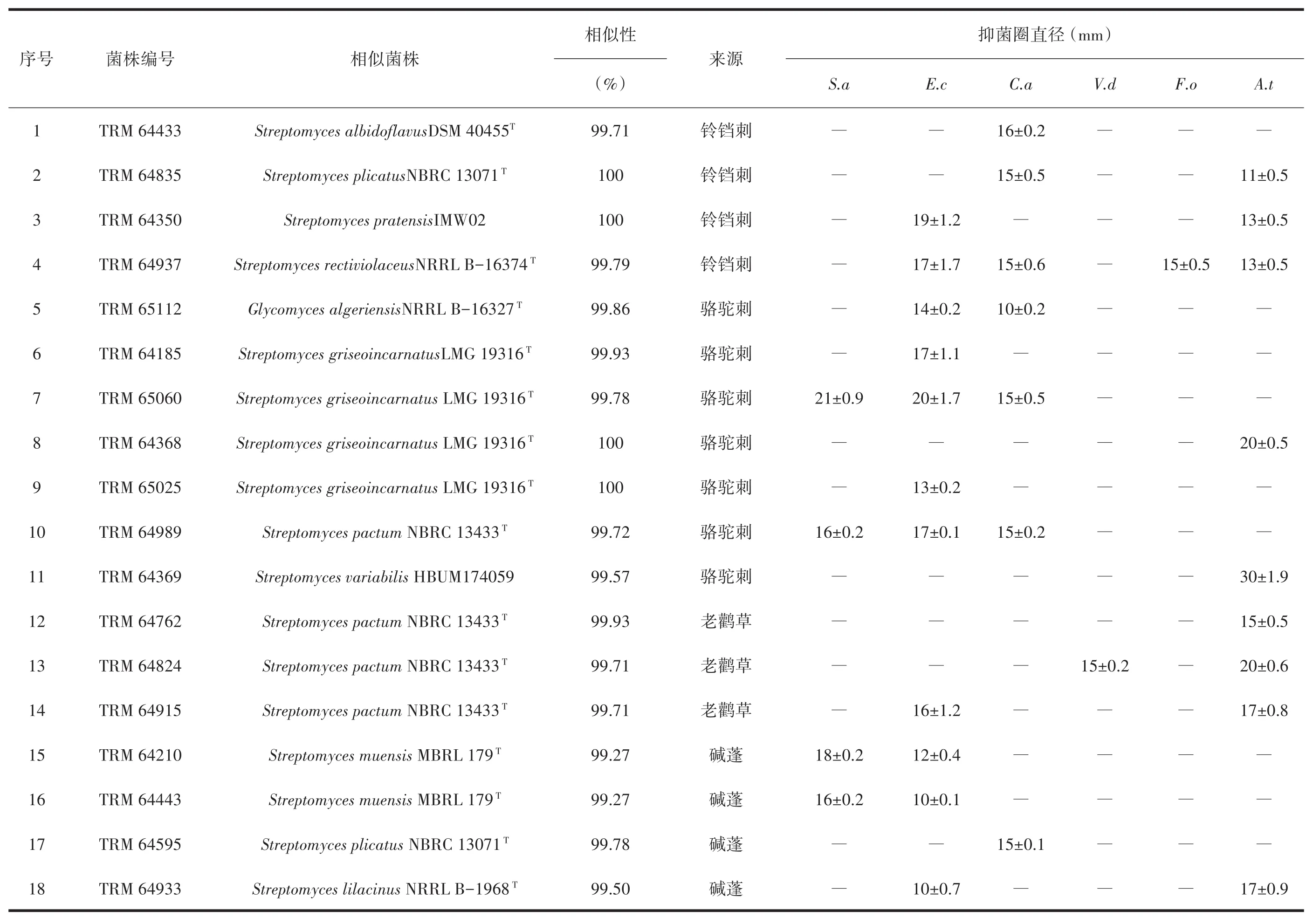
表5 对不同病原菌有较强活性的菌株
续表5 对不同病原菌有较强活性的菌株
3 结论与讨论
近年来,随着临床耐药菌和植物真菌病害的发展,热带雨林植物、药用植物、红树林植物及经济作物内生放线菌的研究与日俱增,荒漠植物内生放线菌鲜有研究。因此,本实验选用13种荒漠植物为材料,分析其内生放线菌的多样性,研究荒漠植物内生放线菌在抗菌方面的潜在应用价值。
13种荒漠植物共分离出533株内生放线菌,去除重复后对361株进行16S rRNA基因序列测定,结果显示共有9个属。研究表明类似土壤环境,链霉菌在植物样品中也广泛分布[16],是植物体内优势放线菌,本研究链霉菌也是优势属,占鉴定菌株总数的88%,在13种样品根部均有分布。这些植物样品中内生放线菌多样性最丰富的是骆驼刺,由6个属组成:链霉菌属(Streptomyces)、拟诺卡氏菌属(Nocardiopsis)、诺卡氏菌属(Nocardia)、伦茨菌属(Lentzea)、糖霉菌属(Glycomyces)、链孢囊菌属(Streptosporangium)。张晓波[17]等人,采集新疆北疆克拉玛依戈壁荒漠的骆驼刺进行内生细菌分离鉴定时,分离出放线菌门中的棒杆菌属(Corynebacterium)、考克氏菌属(Kocuria)、细杆菌属(Microbacterium)和微球菌属(Micrococcus)。同为骆驼刺样品,但多样性差异很大,主要原因可能是地理位置不同,南疆和北疆的气候差异明显。南疆被山脉环绕,受不到大气环流影响,降水量极少,不仅干旱,受纬度和沙漠影响辐射也更强,相同样品的内生放线菌群落结构组成会不同。这与前期学者研究结果一致:不同地区的同种植株内生放线菌,在数量与种类上分布有差异的规律[2]。
稀有放线菌是链霉菌以外其他放线菌的统称,拟诺卡氏菌属是本研究中植物内生稀有放线菌中优势属,这与祁鹤兴和祖丽皮亚木·木沙尔[6,18]等人研究结果相似。本研究值得注意的是,除拟诺卡氏菌属外,拟无枝酸菌属也是13种荒漠植物内生稀有放线菌中的优势属,这可能与植物所处的干旱盐碱生境有关,有研究表明拟无枝酸菌通常存在于干旱环境[19-20],拟诺卡氏菌属的分离也大多对盐分需求较高[21-22]。新疆为“三山夹两盆”地形,降水稀少,水源主要为山区的大小河流,这些河流溶解了古生代海相岩层中的盐分,流入盆地[23],在蒸发大、极端干旱气候条件下,将盐分积累在土壤中。为适应盐渍化土壤,植被多为泌盐盐生植物如胡杨、柽柳和茎叶肉质化的盐生植物如盐节木、新疆猪毛菜,这些盐生植物体内含有大量可溶性盐,为拟诺卡氏菌属和拟无枝酸菌属提供了适宜的生境。除老鹳草外,其余12种植物均为能适应新疆干旱盐渍生境的盐生植物,新疆的地理气候和植物生长的土壤类型可能与两类稀有放线菌定殖相关。
从抗菌活性筛选结果看,筛选到80株活性放线菌,占比22%,是活性放线菌的主要类群,这与先前学者研究结果链霉菌是拮抗菌株的重要来源[16]相符。本实验选用棉花枯黄萎病病原菌和红枣黑斑病病原菌为靶标菌,尝试运用当地微生物资源解决当地农作物生产过程中遇到的植物真菌感染。TRM 6478416S rRNA基因序列与S.bangladeshensis相似性100%,该菌对棉花黄萎病病原菌V.dahliae有很好的抑制作用,有研究表明S.bangladeshensis可产生真菌抑制剂邻苯二甲酸双-(2-乙基己基)酯(bis-(2-ethylhexyl)phthalate)[24]。TRM 6436916S rRNA基因序列与S.variabilis相似性99.57%,对红枣黑斑病病原菌A.tenuissima有很强的抑制作用,这与 Gherbawy[25]研究结果类似,其结果发现S.variabilisYH21具有几丁质酶活性,几丁质酶有降解病原真菌细胞壁的功能。此外,S.variabilis还有产肝素酶的能力,能抗血栓,以及抑制临床细菌生物膜的作用。说明TRM 64784和TRM 64369具有潜在的应用价值,尤其是在作为生物防治棉花黄萎病和红枣黑斑病方面,可作为生防菌株进行进一步的研究。
对不同靶标菌的抑制,本研究中荒漠植物内生放线菌对大肠杆菌有抑制作用的菌株最多,细极链格孢菌次之。这与祖丽皮亚木·木沙尔[18]研究的新疆药用植物百里香内生放线菌的抗菌活性结果有差异,他们的活性菌株中对植物病原真菌有活性的最多,对人类致病的革兰阳性和阴性菌有活性的菌株最少。S.anulatus对植物病原真菌具有广谱抑制性[26],TRM 64326的16S rRNA基因序列与其相似性100%却对大肠杆菌表现出很强的抑制作用,仅对供试的3种植物病原真菌中棉花枯萎病病原菌F.oxysporum有微弱抑制。说明菌株来源不同,对病原菌的抑制作用也会不同,荒漠植物内生放线菌的抗菌谱和抗菌强度也类似多样性,表现出地域和植物种类的差异。此外,本研究中同一骆驼刺植株中TRM 64368、TRM 64461和TRM 65025三株链霉菌与S.griseoincarnatus16S rRNA基因序列相似性均是100%,但它们的抑菌活性差异很大,见表5。表示在抗菌活性筛选时,同一植株中与典型菌株为同一物种的菌株,活性也会不同;即同一物种的菌株活性有差异,在筛选时对每个菌株都进行检测,可以尽可能避免遗漏活性菌株。
本次研究,活性放线菌菌株主要有链霉菌,其次是拟无枝酸菌。拟无枝酸菌属是一类重要的抗生素产生菌,以产生用于临床抗耐药菌感染治疗的万古霉素和抗结核的利福霉素备受关注,广泛应用于医药。近年来,陆续研究发现,拟无枝酸菌产生的新化合物占稀有放线菌产生的新天然产物11%[15],表明拟无枝酸菌属和拟诺卡氏菌属一样是寻找有用天然产物的重要来源。
从13种阿克苏地区荒漠植物鉴定出361株内生放线菌,进行了多样性和抗菌活性研究。荒漠植物内生放线菌丰富的物种多样性,为放线菌资源贮备和后续深入开展活性次级代谢产物挖掘打下基础,具有研究和潜在应用价值。
