新疆传统发酵乳制品中产γ-氨基丁酸乳酸菌的筛选与益生特性研究
2020-09-16王腾斌李宝坤卢士玲王庆玲董娟
王腾斌,李宝坤,卢士玲,王庆玲,董娟
(石河子大学 食品学院,新疆 石河子 832003)
γ-氨基丁酸(γ-aminobutyric acid,GABA)是一种四碳的非蛋白质氨基酸,广泛存在于微生物、植物、脊椎动物以及昆虫中。在植物和细菌中,它在三羧酸循环中起到关键的代谢作用,在脊椎动物大脑中起到神经信号传递的作用[1]。GABA具有突出的生理功能,例如:降血压、治疗肝损伤、治疗抑郁和癫痫等神经性疾病、预防糖尿病等[2-5]。
现今GABA合成的方法主要有4种:化学合成法、植物富集法、酶合成法与微生物合成法。对这4种方法对比发现:化学合成法制备的GABA无法运用于食品工业与饲料中;植物富集法通过富集后,GABA含量仍然较低;酶合成法由于多以固定化酶的方式生产,成本相对过高;而微生物合成法则具有成本低、含量高、可用于食品等特点[6],因此高产GABA菌株的筛选也变得更加重要。
新疆拥有丰富的乳品资源与千古流传的传统制作工艺,各民族凭借这种优势,制作出各种传统发酵乳制品。由于在制作过程中,经天然的乳酸菌与酵母菌参与,使得这些传统乳制品中具有丰富的微生物资源,这为功能性乳酸菌、酵母菌的筛选提供了宝贵的微生物资源[7]。并且乳酸菌一般被视为GRAS级微生物,其能生产食品级GABA已被普遍共识。因此,本研究旨在从新疆传统发酵乳制品奶酪、酥油、酸奶中筛选出GABA合成量较高的乳酸菌,并对其益生特性进行初步分析,为今后富含GABA食品的研发提供优良的乳酸菌资源。
1 材料与方法
1.1 材料与试剂
材料:新疆传统奶酪、酥油、酸奶,分别采自新疆阿勒泰市和伊宁县的不同牧民家庭和市场。
试剂:乙腈、三乙胺、苯异硫氰酸酯、γ-氨基丁酸均为色谱级,美国Sigma公司;谷氨酸钠、DPPH、邻菲罗啉、过氧化氢、铁氰化钾、三氯乙酸、正丁醇、冰乙酸、硫代乙醇酸钠,上海麦克林生化科技有限公司;薄层色谱硅胶,上海盛亚化工有限公司;PCRMix试剂,天根生化科技有限公司;MRS培养基,青岛高科园海博生物技术有限公司。
1.2 培养基
MSG-MRS培养基:向MRS液体培养基中加入1%谷氨酸钠,pH调至6.5。
MRS-THIO培养基:向MRS液体培养基中加入0.2%硫代乙醇酸钠。
1.3 实验菌株
指示菌株:表皮葡萄球菌CICC10294、大肠杆菌CICC10662、金黄色葡萄球菌CICC10348、肠炎沙门氏菌CICC21482,由中国工业微生物菌种保藏管理中心保藏。
1.4 仪器与设备
LDZX-30KBS立式压力蒸汽灭菌器 上海申安医疗器械厂;5417R型高速冷冻离心机 德国Eppendorf公司;多功能酶标仪 美国BioTek仪器有限公司;SPX-150B-Z型生化培养箱 苏州江东精密仪器有限公司;梯度PCR仪 美国BIO-RAD公司;BenchTop Pro冷冻干燥机 美国VirTis公司;LC-20A高效液相色谱仪 日本岛津公司。
1.5 乳酸菌的筛选
1.5.1 菌株的分离纯化
参考Xiong等[8]的方法,利用梯度稀释涂布法与平板划线法对新疆传统奶酪、酥油、酸奶中的菌株进行分离与纯化,将分离的菌株于-80 ℃甘油管保藏。
1.5.2 16S rDNA分子学鉴定
利用细菌基因组DNA提取试剂盒进行分离菌株DNA的提取。PCR扩增引物:27f(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492r(5′-GGTTACCTTGTTACGACTT-3′);PCR扩增体系:12.5 μL PCRMix、9.5 μL ddH2O、27f与1492r引物各1 μL、模板DNA 1 μL;PCR扩增程序:95 ℃ 5 min,94 ℃ 30 s,48 ℃ 30 s,72 ℃ 1 min,30个循环,72 ℃ 8 min。将扩增后的PCR产物进行1%琼脂糖凝胶电泳,若在1450 bp处有单一条带且无非特异性条带存在,则送至上海生工生物工程公司进行测序,将测序结果于NCBI上进行BLAST比对,利用MEGA 7建立系统发育树。
1.6 产γ-氨基丁酸乳酸菌的筛选
1.6.1 菌株的准备
取-80 ℃保藏的乳酸菌甘油管,按4%的接种量接种于MRS液体培养基中,沾取菌液划线纯化,挑取单菌落于5 mL MRS液体培养基中,连续活化2次,最后一次培养至OD600=1备用。
1.6.2 薄层色谱法定性
参考Kwon等[9]的方法,稍作修改:将1.6.1中菌液按4%接种于5 mL MSG-MRS培养基中,培养48 h后,10000 r/min离心10 min,获得上清液。各取1 μL 1 mg/mL MSG标准溶液、1 mg/mL与2 mg/mL GABA标准溶液,菌株上清液点样于薄层色谱硅胶板上,在含有0.4 g茚三酮的正丁醇∶冰醋酸∶蒸馏水为3∶1∶1的100 mL展开剂中层析90 min,105 ℃显色15 min。
1.6.3 Berthelot法初步定量
将1.6.2中方法获得的上清液按照梁慧等[10]的方法进行定量。标准曲线的绘制稍作修改:取0.1 mg/mL(0.9 mg/mL MSG)、0.2 mg/mL(0.8 mg/mL MSG)、0.4 mg/mL(0.6 mg/mL MSG)、0.6 mg/mL(0.4 mg/mL MSG)、0.8 mg/mL(0.2 mg/mL MSG)、1 mg/mL(0 mg/mL MSG)的混合标准溶液绘制GABA标准曲线。
1.6.4 HPLC精确定量
参考Kantachote等[11]的方法,部分优化。
柱前衍生:按照1.6.2中方法获得上清液,取1 mL GABA标准溶液或适当稀释的上清液冷冻干燥,将残余物中加入1 mL乙醇-水-三乙胺(4∶4∶2)溶液和80 μL乙醇-水-三乙胺-苯异硫氰酸酯(6∶1∶1∶1),在室温下避光反应20 min,形成PITC-GABA,经0.22 μm滤膜过滤,上样分析。
色谱条件:色谱柱:Shimazu C18色谱柱(4.6 mm×250 mm),流动相A:1.4 mmol/L乙酸钠缓冲溶液,并加入0.1%三乙胺和6%乙腈, pH调至6.1,流动相B:60%乙腈;洗脱程序:0%~100%流动相B;梯度洗脱50 min;0%流动相B,保持10 min,流速:1 mL/min,进样量:10 μL。选取0.01,0.1,0.25,0.4,0.5 mg/mL的GABA标准溶液,绘制标准曲线。
1.7 耐酸能力分析与胆盐耐受性评价
1.7.1 耐酸能力评价与添加谷氨酸钠的影响
参考Leite等[12]的方法,稍作修改:将1.6.1中菌液在12000 r/min下离心10 min,去除上清液,并重悬于0.02 mol/L pH 6.5的PBS缓冲液中,稀释至OD600=0.6,并以10倍稀释接种于pH 2.5,3.0,4的MRS-THIO液体培养基中,以pH 6.5作为对照,在37 ℃培养0,1,2,3,4 h,分别进行活菌计数。存活率(%)=[人工胃液/肠液处理后活菌数(log CFU/mL)]/最初活菌数(log CFU/mL)。
基于对食品安全与风味考虑,食品中谷氨酸钠推荐添加量为0.1%~0.8%[13],因此向MRS-THIO液体培养基中加入0.5%谷氨酸钠,其他操作相同,进行活菌计数,确定谷氨酸钠对菌株耐酸能力的影响。
1.7.2 胆盐耐受性评价
参考Delgado等[14]的方法,稍作修改:将1.6.1中菌液按照4%接种量接入含有0.15%、0.3%、0.4%牛胆盐的MRS-THIO液体培养基中,在37 ℃下培养,以不添加牛胆盐MRS液体培养基作为对照,培养至OD620上升0.3个单位。对比添加牛胆盐组与对照组,观察生长情况与滞后时间。
1.8 益生特性评价
1.8.1 胃肠道模拟与添加谷氨酸钠的影响
参考Guo等[15]的方法进行人工胃液与肠液的配制。食物在胃里的停留时间一般为3~4 h,流体食物一般为1~3 h。人体胃中pH由于饮食结构的不同波动较大,一般在pH 3.0左右,而肠道内pH值约为8.0,食物在肠道内停留的时间一般为2~6 h[16],因此本文选择在pH 3的人工胃液中处理3 h进行活菌计数,之后连续接种于pH 8的人工肠液中处理6 h进行计数。另外,向人工胃液中加入0.5%谷氨酸钠,其他操作相同,确定谷氨酸钠对胃肠道模拟实验的影响。存活率计算同1.7.1。
1.8.2 抗氧化能力
1.8.2.1 完整细胞组与无细胞提取物的制备
按照1.6.1中方法获得菌液,8000 r/min离心15 min,去除培养基,用0.02 mol/L pH 7.2的PBS缓冲溶液洗去培养基,重复3次,最后重悬于PBS缓冲溶液,使OD600=1。一组作为完整细胞组进行测定,另一组进行超声波破碎(总功率750 W,破碎效率75%,温度4 ℃,破碎时间5 min,破碎6 s间歇3 s),破碎后10000 r/min离心15 min,取上清液于离心管中,在显微镜下镜检,至无完整细胞后立刻进行抗氧化实验。
1.8.2.2 DPPH自由基清除能力
参考He等[17]的方法进行DPPH清除率实验与计算。
1.8.2.3 羟自由基清除能力
参考Zhang等[18]的方法进行羟自由基清除率实验与计算。
1.8.2.4 还原能力测定
参考杨静秋[19]的方法进行乳酸菌还原能力实验与计算。
1.8.3 抑菌能力
参考杨靖鹏等[20]的方法,以表皮葡萄球菌CICC10294、大肠杆菌CICC10662、金黄色葡萄球菌CICC10348、肠炎沙门氏菌CICC21482作为指示菌,利用双层琼脂牛津杯法进行实验。
1.9 数据处理
本文利用Edraw进行细胞图绘制;OriginPro 2016进行图表的制作;利用SPSS进行显著性分析;实验均为3次平行3次重复,以平均值±标准偏差表示。
2 结果与分析
2.1 乳酸菌筛选结果
从新疆传统乳制品奶酪、酥油、酸奶中共分离出163株菌株,其中有4株革兰氏染色结果为阴性,9株过氧化氢酶实验结果为阳性,则共有150株菌株疑似为乳酸菌。经16S rDNA分子学鉴定主要涵盖4个属11个种,见图1。主要为乳杆菌属(Lactobacillus):短乳杆菌(Lactobacillusbrevis)63株、瑞士乳杆菌(Lactobacillushelveticus)26株、副干酪乳杆菌(Lactobacillusparacasei)12株、植物乳杆菌(Lactobacillusplantarum)10株、罗伊氏乳杆菌(Lactobacillusreuteri)5株、发酵乳杆菌 (Lactobacillusfermentum) 3株、鼠李糖乳杆菌(Lactobacillusrhamnosus)3株;肠球菌属(Enterococcus):屎肠球菌(Enterococcusfaecium)8株、鸟肠球菌(Enterococcusavium)4株;片球菌属(Pediococcus):戊糖片球菌(Pediococcuspentosaceus)13株;魏斯氏菌属(Weissella):融合魏斯氏菌(Weissellaconfusa)3株。

图1 部分乳酸菌的系统发育树Fig.1 Phylogenetic tree of some lactic acid bacteria
分析发现,新疆传统奶酪、酥油、酸奶中优势菌属主要为乳杆菌属,其中短乳杆菌、瑞士乳杆菌为主要的优势菌种。刘晓蓉[21]对新疆不同地区传统酸奶样品中的乳酸菌进行分析发现乳杆菌属为优势菌属,德式乳杆菌为主要的优势菌种,张兴吉[22]通过对青海、甘肃、新疆、内蒙古、西藏地区的乳酸菌多样性进行分析发现,乳杆菌属为主要的优势菌属,其中青海地区以瑞士乳杆菌、发酵乳杆菌与德式乳杆菌保加利亚亚种为主要的优势菌种,甘肃与新疆地区以瑞士乳杆菌为主要的优势菌种,内蒙古地区以开菲尔乳杆菌与瑞士乳杆菌为主要的优势菌种,西藏地区以副干酪乳杆菌、植物乳杆菌与德式乳杆菌保加利亚亚种为主要的优势菌种。与本文相比,在菌属方面与其他研究者得到相同的结论,而在优势菌种上具有一定的差异,这可能是由于不同地区因海拔、温度、气候、季节、传统工艺等不同,导致乳酸菌的分布具有一定的差异[23]。
2.2 产γ-氨基丁酸乳酸菌筛选结果
2.2.1 薄层色谱结果
从150株乳酸菌中共筛选得到57株乳酸菌具有GABA合成能力,主要为短乳杆菌52株、屎肠球菌3株、戊糖片球菌2株,发现短乳杆菌在新疆传统奶酪、酥油、酸奶中为主要的GABA合成菌种,并且Wu等[24]得到相同的结论,在筛选出的产GABA乳酸菌中,短乳杆菌为主要的关键菌种,这充分证明短乳杆菌在传统发酵乳制品中,与GABA的形成有着紧密的联系。并且发现并不是所有的短乳杆菌、戊糖片球菌、屎肠球菌均能够合成GABA,这充分证明同一菌种的菌株之间的变异性,菌株合成GABA的能力取决于菌株本身,而非同一菌种的菌株之间都具有合成GABA的能力[25]。

图2 部分乳酸菌发酵上清液薄层色谱图Fig.2 Thin layer chromatogram of some lactic acid bacteria fermentation supernatant
2.2.2 Berthelot法初步定量




图3 Berthelot法定量结果Fig.3 Quantitative results by Berthelot method
通过Berthelot法绘制GABA标准曲线,见图3中A。短乳杆菌NL8 GABA合成能力最强,达到(4.25±0.1) mg/mL。通过分析发现,短乳杆菌GABA合成量均高于1 mg/mL,相比其他两种菌种,在合成GABA方面具有较大的优势,Barrett等[26]通过对人肠道内青春双歧杆菌、婴儿双歧杆菌、齿双歧杆菌以及短乳杆菌的GABA合成能力进行对比,发现短乳杆菌的合成量均高于其他菌种;Renes等[27]从巴西干酪中分离出产GABA的短乳杆菌以及乳酸乳球菌,发现短乳杆菌的合成量整体高于乳酸乳球菌;吕常江发现基于数据库中基因组或功能基因注释的分析显示短乳杆菌通常具有相对较高的GABA合成能力。充分证明短乳杆菌在合成GABA方面是一种很有价值的菌种。
2.2.3 HPLC精确定量

图4 HPLC峰图Fig.4 HPLC peak figure
由于Berthelot法定量可能会受培养基中其他游离氨基酸的影响[28],因此利用HPLC法对短乳杆菌NL8进行精确测量,经统计分析得回归方程为y=140210996.8x-1323464.582,相关系数R=0.9993,证明GABA含量在0~0.5 mg/mL与峰面积具有良好的线性关系。
经HPLC精确测定,短乳杆菌NL8 的GABA合成量为(4.17±0.05) mg/mL,相比通过紫外诱变提高GABA合成量的植物乳杆菌CICC6238(1.196 mg/mL)、从韩国泡菜中分离出的短乳杆菌OPY-1(0.825 mg/mL)和短乳杆菌OPK-3(2.023 mg/mL)[29]等具有相对较高的合成量。
2.3 耐酸能力分析与胆盐耐受性评价
2.3.1 耐酸能力
以2.2.1中筛选得到的不同菌种具有GABA合成能力的戊糖片球菌NL56(产量为(1.51±0.08) mg/mL,记作NL56)与不具备GABA合成能力的瑞士乳杆菌NL13(记作NL13)作为短乳杆菌NL8(记作NL8)的参照菌株,见图5。在未加谷氨酸钠(MSG)时,NL8、NL56、NL13均具有相对较好的耐酸能力,在pH 3条件下处理4 h后,存活率分别为(80.7±1.25)%、(73.39±0.905)%、(76.47±1.05)%,在pH 2.5条件下处理4 h后,存活率分别约为(69.27±0.658)%、(59.31±1.15)%、(62.8±0.675)%。通过对比,NL8具有更好的耐酸能力(p<0.05),Wu等[30]的研究表明能够合成GABA的乳酸菌一般具有较强的耐酸潜力,并建立一种新的方法,通过pH 6.5,1.5,4.0的连续酸化液体培养,结合后期产气现象对产GABA乳酸菌进行筛选,这表明产GABA乳酸菌一般具有较好的耐酸能力,但是可以发现NL13的耐酸能力强于NL56,这可能是由于乳酸菌具有多种耐酸机制且与谷氨酸脱羧酶的活性有一定的关系。



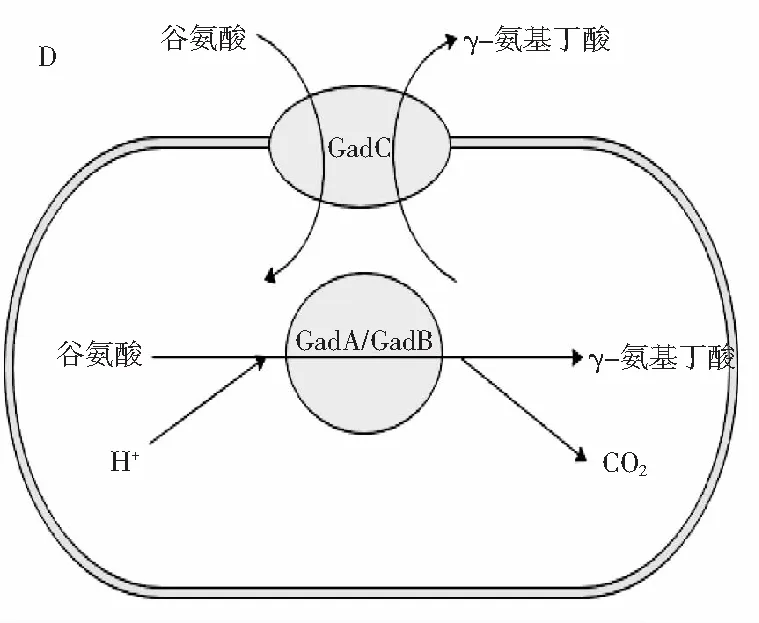
图5 0.5%谷氨酸钠对耐酸能力的影响Fig.5 Effect of 0.5% sodium glutamate on acid resistance
由图5可知,当加入MSG后,NL8与NL56的存活率在pH 3或pH 2.5的条件下处理4 h后,均有显著增加(p<0.05)。NL8在pH 3或pH 2.5处理4 h后,存活率分别增加了约8.02%、6.04%,NL56存活率分别增加了约4.43%、3.75%,而不具备GABA合成能力的NL13存活率未发生改变。这种现象主要是由于产GABA乳酸菌所特有的谷氨酸脱羧酶(GAD)系统所导致的,见图5中D。细胞在受到外界环境酸胁迫的条件下,细胞膜上的Glu/GABA转运蛋白被激活,使得外界的谷氨酸或其盐类进入细胞,在胞质中谷氨酸脱羧酶的作用下消耗胞内H+并生成GABA,使得胞内pH值回升,GABA由Glu/GABA转运蛋白转移至细胞外,由于GABA偏碱,导致胞外pH回升,从而使细胞内外pH平衡,提高细胞在酸胁迫环境中的生存能力[31]。可以发现,NL8的存活率相比NL64增加的更多,这可能是由于NL8具有更强的GABA合成能力,其谷氨酸脱羧酶活性更强,在酸胁迫条件下,使胞内外pH平衡的效率更高,从而有更多细胞存活。
2.3.2 胆盐耐受性


图6 胆盐耐受性评价Fig.6 Evaluation of bile salt tolerance
由图6可知,NL8的胆盐耐受能力相比NL56较弱,NL56在含有0.15%、0.3%、0.4%胆盐浓度的环境中,生长滞后时间分别约为1,2.5,4 h,而NL8在0.15%胆盐浓度下滞后时间约为4 h,胆盐浓度为0.3%以上时,生长受到强烈抑制,10 h内OD620值仍未上升0.3,这主要是由于胆盐可以通过破坏乳酸菌细胞膜上的脂质和脂肪酸从而引起细菌衰亡,而乳酸菌耐受胆盐主要有两种方式:通过分泌胆盐水解酶降解胆汁盐,使其正常生长以及胞外多糖包裹细胞,防止胆盐的破坏[32]。NL8胆盐耐受能力较差,因此如何增强细胞对胆盐的耐受能力也变得尤为重要。
2.4 益生特性评价
2.4.1 胃肠道模拟耐受性
在2.3.1中通过对两株菌的耐酸能力进行分析发现,两株菌均具有较好的耐酸能力,且通过添加0.5% MSG,在pH 3或pH 2.5处理3 h或4 h后,存活率均有显著的提高,因此有必要确定MSG是否可以在胃肠道模拟过程中起到积极作用。


图7 0.5%谷氨酸钠对模拟胃肠道耐受能力的影响Fig.7 Effect of 0.5% sodium glutamate on simulated gastrointestinal tolerance
由图7可知,在未加0.5%MSG的环境中,NL8经人工胃液、肠液处理后存活率分别为(86.36±1.25)%、(80.5±1.05)%,NL56分别为(81.48±0.758)%、(71.08±0.865)%。史晓萌等通过对植物乳杆菌 CICC 6238以及从怡能菌粉中分离出的嗜酸乳杆菌NCFM和鼠李糖乳杆菌HN001进行胃肠道模拟试验,最终存活率分别约为61.5%、79.4%、77.4%,对比发现,NL8和NL56均具有较好的胃肠道耐受能力且NL8的耐受能力更强(p<0.05)。
在加入0.5% MSG后,经人工胃液处理3 h、人工肠液处理6 h后,NL8和NL56的存活率相比未加MSG均显著升高(p<0.05),这主要是由于添加MSG后增强了NL8与NL56的耐酸能力(如2.3.1所述),使得在人工胃液中处理3 h后细胞数增多,而人工肠液对乳酸菌的抑制作用是一定的,从而使得最终存活率显著增加。因此,通过添加0.5% MSG可以有效改善NL8与NL56在人体胃肠道中的耐受情况。
2.4.2 抗氧化能力评价
通过对NL8与NL56的抗氧化能力进行分析发现,NL8具有较强的DPPH自由基清除能力,其完整细胞与无细胞提取物分别达到约(25.59±0.54)%、(42.29±0.87)%。李默[33]从发酵肉制品中分离出30株乳酸菌并对完整细胞和无细胞提取物进行DPPH自由基清除能力的测定,发现清除范围在0%~34.8%,而NL8的无细胞提取物高于这个范围,并且相比嗜酸乳杆菌874(32.9%)、商业菌种嗜酸乳杆菌NCFM(22.6%)、植物乳杆菌ATCC14917(38.0%)[34]均高,则说明NL8具有较强的DPPH清除能力。



图8 抗氧化能力评价Fig.8 Evaluation of antioxidant capacity
通过对比李默、刘珊春[35]对发酵肉制品、传统发酵酸乳中乳酸菌羟自由基清除能力和还原能力的测定结果,NL8与NL56清除羟自由基能力和还原能力适中,在清除羟自由基方面并无明显差异(p>0.05),在还原能力方面,NL56相比NL8具有更强的能力(p>0.05)。
2.4.3 抑菌能力评价

图9 抑菌能力评价Fig.9 Evaluation of antibacterial ability
由图9可知,NL8对大肠杆菌CICC10662与肠炎沙门氏菌CICC21482具有显著的抑制作用(p<0.05),抑菌圈直径分别为(18.62±0.58) mm和(18.13±0.56) mm,而对金黄色葡萄球菌CICC10348与表皮葡萄球菌CICC10294的抑菌作用显著较弱(p<0.05),仅为(12.29±0.48) mm和(11.62±0.33) mm。Kim等[36]发现,通过将短乳杆菌DF01的粗细菌素与大肠杆菌、鼠伤寒沙门氏菌共同孵育,能够减少大肠杆菌与鼠伤寒沙门氏菌的生物膜形成,从而起到抑制生长的作用,证明大肠杆菌与沙门氏菌对短乳杆菌产生的细菌素较为敏感,因此抑制作用较强。
3 结论
从新疆传统奶酪、酥油、酸奶各6份样品中共分离出150株乳酸菌,主要涵盖4个属11个种,其中乳杆菌属为优势菌属,短乳杆菌与瑞士乳杆菌为主要的优势菌种;通过利用薄层色谱法对产GABA乳酸菌进行筛选,150株乳酸菌中共有57株乳酸菌具有合成GABA的能力,其中包括52株短乳杆菌、3株屎肠球菌、2株戊糖片球菌。利用Berthelot法与HPLC法进行定量发现,短乳杆菌在合成GABA方面较有优势,且筛选出一株产量较高的菌株短乳杆菌NL8,产量达到(4.17±0.05) mg/mL。因此,选择短乳杆菌NL8与具有GABA合成能力的戊糖片球菌NL56[(1.51±0.08) mg/mL]进行后续益生特性的评价。
短乳杆菌NL8相比戊糖片球菌NL56具有更好的耐酸能力(p<0.05),且与不具备GABA合成能力的瑞士乳杆菌NL13进行对比,通过添加0.5% MSG能够使两株菌在低pH的环境下耐受能力显著增强(p<0.05);短乳杆菌NL8相比戊糖片球菌NL56在耐胆盐能力上较弱,短乳杆菌NL8在0.3%胆盐环境中生长开始受到强烈抑制,而戊糖片球菌NL56能够较好地耐受胆盐进行生长;经人工胃肠液处理,短乳杆菌NL8表现出更强的耐受性,且通过添加0.5% MSG可使两株菌在人工胃液环境中耐受性显著增强,从而提高最终存活率(p<0.05);短乳杆菌NL8相比戊糖片球菌NL56具有更强的DPPH自由基清除能力(p<0.05),且两株菌均具有一定的羟自由基清除能力与还原能力;短乳杆菌NL8能够显著抑制大肠杆菌与肠炎沙门氏菌的生长(p<0.05),而对金黄色葡萄球菌与表皮葡萄球菌的抑制作用较弱,戊糖片球菌NL56对指示菌均具有一定的抑制作用。
综上所述,短乳杆菌NL8具有较强的GABA合成能力与优良的益生特性,戊糖片球菌NL56虽然GABA合成量相对较低,但也具有较好的益生特性,且在今后的富含GABA发酵产品中,一般会残留一定量的MSG,而本文已证明通过添加MSG可以增强两株菌在胃肠道中的耐受能力,因此可作为优良发酵剂用于后续功能性产品的研发。
