转入木聚糖酶突变基因重组酵母的产酶能力及酶学性质分析
2020-08-15王国强李淑丽尹清强
王国强,李淑丽,魏 沛,尹清强
(1.南阳农业职业学院,河南 南阳 473000;2.河南农业大学牧医工程学院,河南 郑州 450002)
1 试验材料
1.1 试验时间与地点
试验于2017—2018年在河南农业大学动物营养与饲料生物技术实验室进行。
1.2 试验仪器
BCM-1000生物超净工作台,恒温培养箱(上海跃进医疗器械厂),双层气浴振荡器(金坛市杰瑞尔电器有限公司),离心机(上海安亭仪器厂),高速冷冻离心机(湘南湘仪实验室仪器开发有限公司),涡旋混合仪,PCR仪器,稳流稳压电泳仪(北京六一仪器厂),凝胶成像装置设备(北京君意电泳设备有限公司),紫外-可见分光光度测定计(上海菁华科技仪器有限公司),PHS-2C酸度测定计(天津市得利斯实验分析仪器厂),磁力搅拌器(金坛市华峰仪器有限公司),台式封闭电炉(天津市泰斯特仪器有限公司),电子分析天平,具塞试管,Dionex ICS-3000离子色谱仪等。
1.3 试验菌种
挑选的重组毕赤酵母转化子。
1.4 主要试剂及培养基
1.4.1 试剂 木聚糖及D-木糖购自Sigma公司,小麦麸由河南德林生物股份有限公司提供,胰蛋白胨、酵母浸粉、葡萄糖等均为分析纯。
1.4.2 培养基与溶液
1.4.2.1 培养基的配制
(1)YPD培养基:分别称取胰蛋白胨2 g、酵母浸出物1 g,溶解到80 mL去离子水中,搅拌使其完全溶解;称取无水葡萄糖2 g,溶解于20 mL蒸馏水中,充分搅拌使其溶解。在121℃、1.034 MPa条件下高压灭菌20 min,待冷却后将两者混合均匀。
在胰蛋白胨和酵母浸提物的混合液中加入1.5 g琼脂粉,与葡萄糖溶液高压蒸汽灭菌20 min,两者混匀后,待其温度冷却至50℃以下,可加入抗生素Zecoin摇匀后,将培养基倾倒于玻璃培养皿中,待其完全凝固后,封口膜密封完全后,将平板倒放在4℃冰箱,保存以备使用。
(2)含0.2%的吐温80的YPD液体培养基:取2 g胰蛋白胨、1 g酵母浸粉,溶解于约80 mL去离子水中,搅拌使其完全溶解后,称取2 g小麦麸,再加入0.2%的吐温80,最后定容至100 mL。121℃、1.034 MPa条件下高压蒸汽灭菌20 min备用。
1.4.2.2 溶液的配制
(1)氯化钠溶液:浓度是1 mol/L。称量29.22 g氯化钠,加入含400 mL去离子水的烧杯中,充分搅拌待其完全溶解后,定容到500 mL,混合均匀后常温保存。
(2)氢氧化钠溶液:浓度是200 g/L。称量20 g氢氧化钾(NaOH),放入烧杯内,加水溶解,待其冷却后,定容至100 mL。
(3)木聚糖溶液:浓度为100 mg/mL。称取1.00 g木聚糖(Sigma)、0.32 g氢氧化钠,加 90 mL 水,磁力加热搅拌约30 min直至木聚糖完全溶解,之后停止加热继续搅拌30 min,加入0.5 mL冰乙酸,测定其pH,若其偏离5.50,用乙酸或乙酸钠溶液进行调节,之后用乙酸-乙酸钠溶液定容至100 mL,摇匀后,4℃冰箱避光保存,其有效期为12 h。
2 试验方法
2.1 不同重组酵母菌产酶活力的测定方法
2.1.1 粗酶液的制备
(1)量取100 μL培养好的毕赤酵母培养物,接种于含100 mL YPD液体培养基的250 mL三角瓶中,30℃摇床培养36 h。
(2)取上述培养物,按2%的接种比例接种到含2%麸皮的发酵培养基内,30℃振荡培养,每隔12 h收集1次发酵培养液,3 000 r/min离心3~5 min,收集上清液作为粗酶液。
2.1.2 木聚糖酶活力的测定 将酶液进行适当稀释,取2.00 mL经过37℃平衡的酶液,加入到具塞试管中,再添加DNS试剂5 mL,混合均匀后,加入2.00 mL木聚糖溶液,37℃保温30 min后,沸水浴5 min。用水冷却到常温,添加水定容到25 mL,涡旋混匀后,测定吸光度标记为空白样吸光度。
将酶液进行适当稀释,取2.00 mL经过37℃平衡的酶液,加入到具塞试管中,再添加2.00 mL木聚糖溶液,混合均匀后,37℃保温30 min后,再加入5 mL DNS试剂,沸水浴5 min。用水冷却到常温,添加水定容到25 mL,涡旋混匀后,测定吸光度,标记为酶反应液吸光度。
木聚糖酶活力的计算:稀释酶液的木聚糖酶活力=[(酶反应液吸光度-空白样吸光度)×标准曲线斜率+标准曲线截距]/[木糖的摩尔质量/酶解反应时间]×1 000。
试样木聚糖酶活力=稀释酶液中的木聚糖酶活力×试样的稀释倍数。
2.2 培养时间对重组酵母产酶活力影响
选取木聚糖酶活力高的菌株,每隔12 h进行取样,测定不同时间木聚糖酶活力的变化。
2.3 木聚糖酶酶学性质的分析
2.3.1 不同温度处理对木聚糖酶活力的影响 分别在30℃、40℃、50℃、60℃、70℃、80℃、90℃处理酶液15 min,立即用冰水冷却至室温,测定不同温度对木聚糖酶活力的影响。
2.3.2 不同pH处理对木聚糖酶活力的影响 配制不同pH的缓冲液和底物溶液,将酶液pH分别调至3.50、4.50、5.50、6.50、7.50、8.50、9.50,测定不同 pH对木聚糖酶活力的影响。
2.4 添加0.2%的吐温80对木聚糖酶活力的影响
在含2%麸皮的YPD培养基内,加入0.2%的吐温80,以未加0.2%的吐温80的发酵液作对照,测定木聚糖酶活力。
2.5 木聚糖酶对小麦中非淀粉多糖的降解
2.5.1 色谱条件 采用Dionex ICS-3000离子色谱仪,CarboPac PA10分离柱(4×250 mm)与 CarboPac PA10保护柱(4×50 mm)进行测定。其淋洗液为50 mmol/L NaOH,设定流速为0.8 mL/min,设定柱箱的温度为30℃,进样体积是25 μL。采用电化学检测器检测方式、脉冲安培检测模式进行检测。
2.5.2 小麦的处理 将小麦进行粉碎,留取40~60目之间的样品备用。
2.5.3 粗酶液的制备
2.5.3.1 液体粗酶液的制备
(1)取100 μL培养好的含空 pGAPZαA质粒的毕赤酵母、重组毕赤酵母、突变体毕赤酵母培养物,接种于含100 mL YPD液体培养基的250 mL锥形瓶内30℃,振荡培养36 h。
(2)取上述培养物,按2%的接种比例接种到含2%麸皮的发酵培养基内,30℃振荡培养12 h,取发酵培养液,1 000 r/m离心3~5 min,留取上清液作为粗酶液。
(3)将其进行适当稀释,并将粗酶液的pH调整至4.50。
2.5.3.2 固体粗酶液的制备
(1)用无菌接种环挑取黑曲霉孢子,划线接种于PDA固体培养基上,30℃恒温静置培养约72 h。
(2)用无菌接种环将培养好的孢子转移至0.9%的生理盐水中,配制孢子悬液,并测定其孢子浓度约为 1×109个/mL。
(3)按10%的比例,将孢子悬液接种于固体发酵培养基内,混合均匀后,30℃恒温静置培养72 h。
(4)将固体发酵产物和0.9%的生理盐水按1∶10的比例进行混合,搅拌30 min后,室温静置,过滤,取滤液作为粗酶液。
(5)将其进行适当稀释,并将粗酶液的pH调整至4.50。
2.5.3.3 试验方法 将处理过的小麦和经稀释调整的粗酶液,按固体(g)∶液体(mL)=1∶10 的比例进行混合,40℃,恒温发酵培养2 h。3 000 r/min离心3~5 min,留取上清液作为检测液。将检测液经适当处理后上样检测。
3 结果与分析
3.1 不同突变株产酶活力的测定结果
由图1可知,突变株5、突变株7、突变株8等重组毕赤酵母的木聚糖酶活力并不是很高,只有突变株1-2、突变株4-5及突变株6-3分泌的木聚糖酶活力较高,并且突变株1-2的木聚糖酶活力显著高于其他各组(P<0.05)。这可能是由于氨基酸的变化引起了蛋白质变化,从而影响了酶活性的变化。同时,在同一个重组酵母中也可能由于拷贝数的不同而造成酶活力的改变。因此,选择突变株1-2、突变株4-5及突变株6-3进行下面的试验。
3.2 培养时间对重组酵母产酶活力的影响
由表1可知,突变株和原始基因分泌的木聚糖酶活力均在12 h时达到了最大值,显著高于其他时间点的木聚糖酶活力(P<0.05)。培养时间延长后,木聚糖酶活力不断下降。其中突变株1-2在12 h的木聚糖酶活力达到0.148 0 U/mL,显著高于对照组及原始基因组(P<0.05);同时突变株1-2在24 h、36 h的木聚糖酶活力均显著高于其他各组(P<0.05)。由此选择突变株1-2进行下面的酶学性质分析试验。

表1 培养时间对重组酵母菌木聚糖酶活力的影响 U/mL
3.3 不同温度处理对木聚糖酶活力的影响
突变株在40℃木聚糖酶的活力显著高于其他温度(P<0.05),并显著高于原始基因重组毕赤酵母在40℃的木聚糖酶活力(P<0.05)。由此可见,突变株和原始基因重组酵母的最适温度是40℃。在60℃时,原始基因重组酵母和突变株受温度的影响水平是一致的,两者差异不显著,但经过30℃、50℃、70℃、80℃和90℃的处理,原始基因重组酵母的木聚糖酶活力显著高于突变株(P<0.05)。图2给出不同温度对突变株分泌的木聚糖酶活性,以及不同温度对含原始基因重组酵母分泌的木聚糖酶活性的影响。
以最高木聚糖酶酶活力为100%,研究不同温度对不同菌株分泌的木聚糖酶的相对酶活的影响。由图3可知,突变株受温度影响较大,70℃以后,其相对酶活均小于60%。随着温度的上升,酶活力逐渐下降;而原始基因重组酵母的木聚糖酶活力虽然在50℃以后随着温度的上升而逐渐下降,但其相对酶活仍能保持较高水平。
3.4 不同pH处理对木聚糖酶活力的影响
突变株和含有原始基因重组酵母的木聚糖酶的最适pH均为5.5,显著高于其他各pH点(P<0.05)。pH在3.5、4.5时,含原始基因的重组酵母的木聚糖酶活力均显著高于突变株的木聚糖酶活力(P<0.05),随着pH的上升,突变株和含有原始基因重组毕赤酵母的木聚糖酶的酶活力升到最高,之后都急剧下降,由此可见,这两个酶在中性及碱性条件下,酶和底物的催化作用较低。图4给出不同pH对突变株分泌的木聚糖酶活性,以及不同pH对含原始基因重组酶母分泌的木聚糖酶活性的影响。图5为不同pH对重组酵母木聚糖酶相对酶活的影响。
3.5 添加0.2%的吐温80对木聚糖酶活力的影响
以未添加吐温80的培养基为对照组,结合图6可知,添加0.2%的吐温80均能显著提高突变株和含有原始基因重组酵母的木聚糖酶活力(P<0.05)。同时突变株的木聚糖酶活力提高了约30%,而含有原始基因重组酵母的木聚糖酶活力提高了35%。
3.6 木聚糖酶对小麦中非淀粉多糖的降解结果
由表2可知,黑曲霉分泌的木聚糖酶可显著提高葡萄糖和阿拉伯糖的产量,但其木糖的产量低于突变株(P<0.05)。原始基因重组毕赤酵母及突变株分泌的木聚糖酶与对照组相比,可显著提高木糖的产量(P<0.05),抑制葡萄糖的产量,但阿拉伯糖的产量低于对照组。阿拉伯糖的含量并不是很高,这可能是因为原始基因的毕赤酵母及突变株分泌的木聚糖酶单一,真菌分泌的木聚糖酶多样,从而使真菌中阿拉伯糖和木糖的含量均较高;也可能是因为氨基酸的改变导致酶作用位点发生变化,从而使原始基因重组酵母和突变株对木聚糖中阿拉伯糖残基的降解不显著。
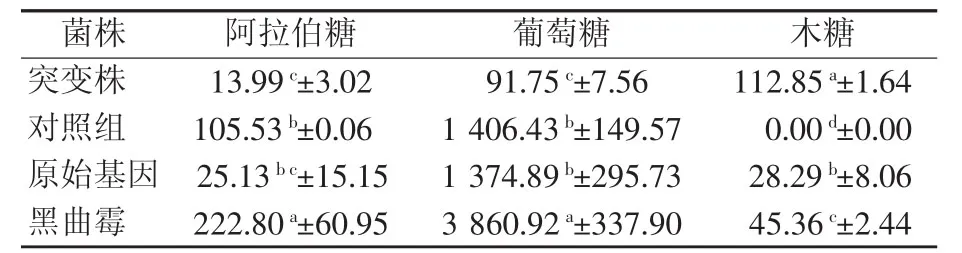
表2 不同来源的木聚糖酶对小麦非淀粉多糖的降解效果 μg/mL
4 讨论
在发酵培养基中加入0.2%的吐温80后,突变株1-2和含有原始基因重组酵母的木聚糖酶活力均显著提高30%左右。吐温80又称聚山梨酯-80,是油酸酯,也是一类非离子型表面活性剂,因能增加细胞膜的通透性,促使多种酶分泌到胞外,从而促进营养物质的摄入及代谢产物的排出[1]。
氨基酸的不同可能导致蛋白质的性质发生变化,这可能是由于蛋白质之间的连接作用和空间结构等的变化所致。本试验表明,突变株4-5的木聚糖酶活力显著高于突变株6-3(P<0.05),同时突变株1-2的木聚糖酶活力显著高于其余突变体(P<0.05)。这表明第116位、第43位、第34位、第179位等氨基酸的变化可能影响了蛋白质的分子结构,从而改变了木聚糖酶的活性。突变体1-2分泌的木聚糖酶活力在12 h显著高于含有原始基因重组酵母(P<0.05),并在其他的时间点也维持较高的木聚糖酶活力,这表明第117位天冬氨酸变为天冬酰胺可能对木聚糖酶的催化基团有积极的影响。
本试验表明,突变株1-2分泌的木聚糖酶最适温度和最适pH与含有原始基因重组酵母一致,这表明氨基酸的细微变化并不能使木聚糖酶的性质发生剧烈的改变。由试验可知,两种基因的重组酵母分泌的木聚糖酶都是常温较耐酸的蛋白质。然而,由于此酶在50℃以后酶活力在持续下降,故而不具有耐高温的特性,因此,该酶不适合制粒[2]和挤压[3]。同时该酶的这种耐酸性决定着它在饲料中的应用效果,因为饲料中的酶只有通过消化道经过胃这种酸性环境到达小肠后才能发挥其作用[4]。这与周晨妍等[5]的研究结果一致,他认为基因突变并未改变原始基因的最适作用温度和热稳定性。Chen等[6]发现,通过易错PCR能使木聚糖酶在pH为8.0时表现活力,而野生酶是在这种条件下是没有木聚糖酶活力的。邓萍等[7]认为重组蛋白糖基化程度的差异会影响其最适温度,但不影响其最适pH的变化。
通过研究发现含有原始基因重组酵母和突变株1-2及黑曲霉所分泌的木聚糖酶可显著提高木糖的产量(P<0.05),黑曲霉分泌的木聚糖酶以提高葡萄糖和阿拉伯糖产量为主,突变株分泌的木聚糖酶有助于木糖产量的提高。这与雷丽[8]的研究有所差异,他认为添加木聚糖酶能提高阿拉伯糖、木糖残基和葡萄糖残基在嗉囊和肌胃中的消化率(P<0.05),使水溶性非淀粉多糖在嗉囊中的消化率显著提升(P<0.05)。两者有差异可能是由于氨基酸的些微变化致使酶作用位点发生了变化,但其具体原因还需进一步研究才能解决。
5 小结
本研究从6个突变体中分别挑选5个重组酵母,通过对30个突变株的木聚糖酶活力进行检测,最终发现突变株1-2在12 h的木聚糖酶活力显著高于其他各突变株,并显著高于含有原始基因重组酵母的木聚糖酶活力。通过对突变株1-2木聚糖酶和原始酶的酶学性质分析发现,突变株1-2的最适温度和最适pH与原始木聚糖酶的酶学性质基本一致。同时也证实了加入0.2%的吐温80可使突变株1-2和原始木聚糖酶活力均提高30%左右。进一步研究发现黑曲霉分泌的木聚糖酶可显著提高葡萄糖和阿拉伯糖的产量,但其木糖的产量低于突变株(P<0.05)。原始基因重组毕赤酵母及突变株分泌的木聚糖酶与对照组相比,可显著提高木糖的产量(P<0.05),抑制葡萄糖的产量,但阿拉伯糖的产量低于对照组(P<0.05)。
