广西红椎树干径向生长研究
2020-08-05刘士玲杨保国姚建峰庞圣江廖树寿邹位锡
刘士玲,杨保国,姚建峰,郑 路,张 培,庞圣江,廖树寿,邹位锡
(1 中国林业科学研究院 热带林业实验中心,广西 凭祥 532600;2 广西友谊关森林生态系统国家定位观测研究站,广西 凭祥 532600;3 中国林业科学研究院 资源信息研究所,北京 100091)
森林生态系统是陆地生物圈的最大组成部分,在调节土壤−植物−大气连续体的碳和水交换方面发挥着关键作用[1]。树木作为森林生态系统的基本单元,对其径向生长与气候变化的关系研究有助于了解森林生态系统对气候变化的响应机制[2]。树干径向变化记录仪可以提供长期和连续的高时间分辨率的径向变化测量,已被广泛应用于树木径向生长监测[3-6]。树木气候学研究表明,中国南方亚热带树木生长对气候变化的反应不一,如暖春树木生长增加,但夏季由于干旱而树木生长下降[7-10]。这些对比鲜明的树木生长反应清楚地表明,亚热带森林存在复杂的气候−生长关系。因此,迫切需要有关生长−气候关系的信息,以提高我们对亚热带森林应对未来气候变化的认识。然而,以往对环境因素影响树木生长的研究主要是基于优势木[11],且缺少精细分辨率生长−气候数据;另外,一般木和被压木在林分中占很大比例,在木材生产中有重要的贡献。许多功能过程与树的大小密切相关,因此随着树的年龄和大小的增加而发生变化[12-13]。在一个林分中,不同大小和生态位的树木竞争光、水和其他资源,因此可能会对环境压力有不同的反应[14-15]。在气候变化背景下,理解气候−生长关系尤为重要。但国内基于精细径向生长观测的气候−生长关系研究并不多见。
红椎Castanopsis hystrix是我国亚热带地区重要的乡土阔叶珍贵用材和高效多用途树种,具有生长快、材质优、适应广、效益高等优良特性[16]。本研究以南亚热带19年生红椎人工林为对象,依托广西友谊关森林生态系统国家定位观测研究站,通过对气象环境因子和树干径向生长的连续观测,试图探究以下问题:1)评估不同分化等级红椎树干径向生长的日变化和季节变化;2)确定红椎各分级木的主要生长时期;3)探讨径向生长的影响因子及作用机制,找到主导因子并确定数量关系。这对于丰富南亚热带红椎树干径向生长动态的观测研究,了解全球气候变化对红椎生长的影响,应对气候变化的适应性森林经营等都有一定的理论和实践意义。
1 研究区域概况
本研究在广西友谊关森林生态系统国家定位观测研究站实验区范围内 (21°57′~22°16′ N,106°41′~106°59′E)的红椎人工林内进行。区域内日照充足,雨、热充沛,有明显的干湿季。属南亚热带半湿润−湿润季风气候,年均气温20.5~21.7 ℃,极端高温40.3 ℃,极端低温−1.5 ℃。年均降雨量 1 200~1 600 mm,78%降雨量发生在5—10月(1965—2015年),年均蒸发量1 261~1 388 mm。土壤是花岗岩风化形成的红壤,质地为砂质壤土。红椎为2001年种植,林下天然更新红椎幼苗较多,灌木主要有九节Psychotria rubra、粗叶榕Ficus hirta、鲫鱼胆Maesa perlarius等,草本主要有扇叶铁线蕨Adiantum flabellulatum、弓果黍Cyrtococcum patens等。
2 研究方法
2.1 径向生长数据采集
2018年 1月选择生长良好的红椎人工林,建立20 m×20 m的试验样地,在对林分进行全面调查的基础上,采取定性和定量相结合的方法,在林分内选取优势木、一般木和被压木共3株[17-18](表1),在树干1.3 m高处刮去部分翘裂及干枯树皮,安装树干径向变化记录仪(DC3,Ecomatik,德国),连续记录径向生长变化过程。数据采集器型号为DL18(Ecomatik,德国),采集时间间隔为 15 min。

表1 红椎样树的基本特征Table 1 Characters ofCastanopsis hystrix sample trees
2.2 气象环境因子测定
采用广西友谊关森林生态系统国家定位观测研究站的自动气象站的气象数据(直线距离<1.5 km),气象环境因子包括空气温度(θ/℃)、降雨(P/mm)、太阳日均辐射 [SR/(μmol·s−1·m−2)]、空气相对湿度(RH/%)和深度为20和40 cm土壤的体积含水率(φs20/%、φs40/%)、土壤温度 (θs/℃)等因子。该气象站采用的是CR1000数据采集器(Campbell,美国),各要素数据采集步长均为1 min,数据分析时所有气象数据均换算为15 min的平均值,与径向生长数据一一对应。饱和水汽压差(VPD/kPa)和大气水势(Pa/Pa)分别基于空气相对湿度、空气温度和大气绝对温度(Ta/K)的数据计算得到[19-20],计算公式如下:
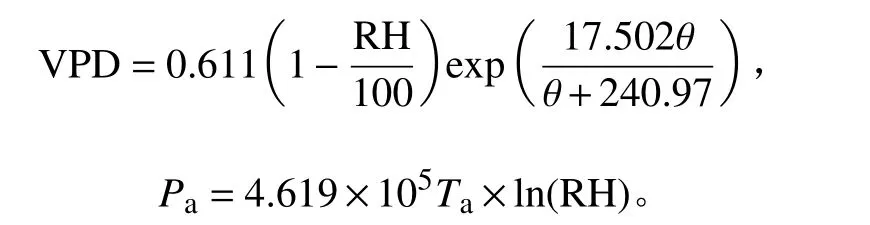
2.3 数据处理及分析
本研究所用的是 2018年观测的树干径向生长数据和气象环境因子数据。
采用最大值法[21]获得径向生长信息,即提取每日径向数据中的最大值,计算连续两日的最大值差值,求得日变化(ΔR)。树干径向累积变化量为逐日ΔR的累加之和。
树干径向生长拟合采用Gompertz生长模型[22],对拟合函数求一阶导数得到树木生长的速度曲线。生长速度大于测量仪测量精度(4 μm/d)的时期为树木主要生长时期,并用于之后的生长环境相关分析研究[23]。Gompertz生长模型表达公式如下:

其中:x为天数,y为树干径向累积变化量,a为y的上渐进值,e是自然对数的底数,b为与y初始值有关的参数,c为内秉生长率,方程的拐点为b/c,此时y的变化速度最大。
利用Excel2010软件对数据进行汇总和初步分析;利用SPSS19.0进行Pearson相关分析、偏相关分析和主成分分析;利用Origin9.0、Sigmaplot10.0软件进行径向生长拟合和做图等。
3 结果与分析
3.1 树干径向生长日变化特征
国内外有关树干径向变化的研究都发现其具有日周期模式,其中,夏季模式与水分相关,夜间温度较低,根系吸水强于蒸腾耗水,径向膨胀,早晨温度上升后,蒸腾耗水不断增强,径向失水收缩,直到下午温度降低后,径向才再次吸水膨胀[24],日径向变化波动呈正弦曲线;冬季模式(即冰冻−解冻周期)与温度密切相关,夜间径向冷冻收缩,白天温度上升后解冻膨胀,表现出与夏季模式相反的变化动态[25]。年尺度上的红椎树干径向生长月均日变化周期如图1所示,观察到红椎树干径向生长的循环模式只有1种类型,即典型的夏季模式,树干径向变化呈正弦曲线,每天上午出现峰值,下午出现谷值。12个月的月平均日变化的振幅有所不同,日平均气温的增加和日照的增加使日振幅增强,1—2月份径向变化的振幅较小,3—10月份振幅较大,11—12月份振幅又减小,即温度较高时径向变化比较剧烈,气温较低时径向变化较平缓。从不同分化等级样树的径向日变化周期来看,优势木的径向变化峰值、谷值和振幅明显大于一般木和被压木,优势木的径向变化峰值大多出现在09:00或10:00,谷值出现在16:00或17:00,一般木和被压木的径向变化峰值和谷值比优势木推后1~2 h。
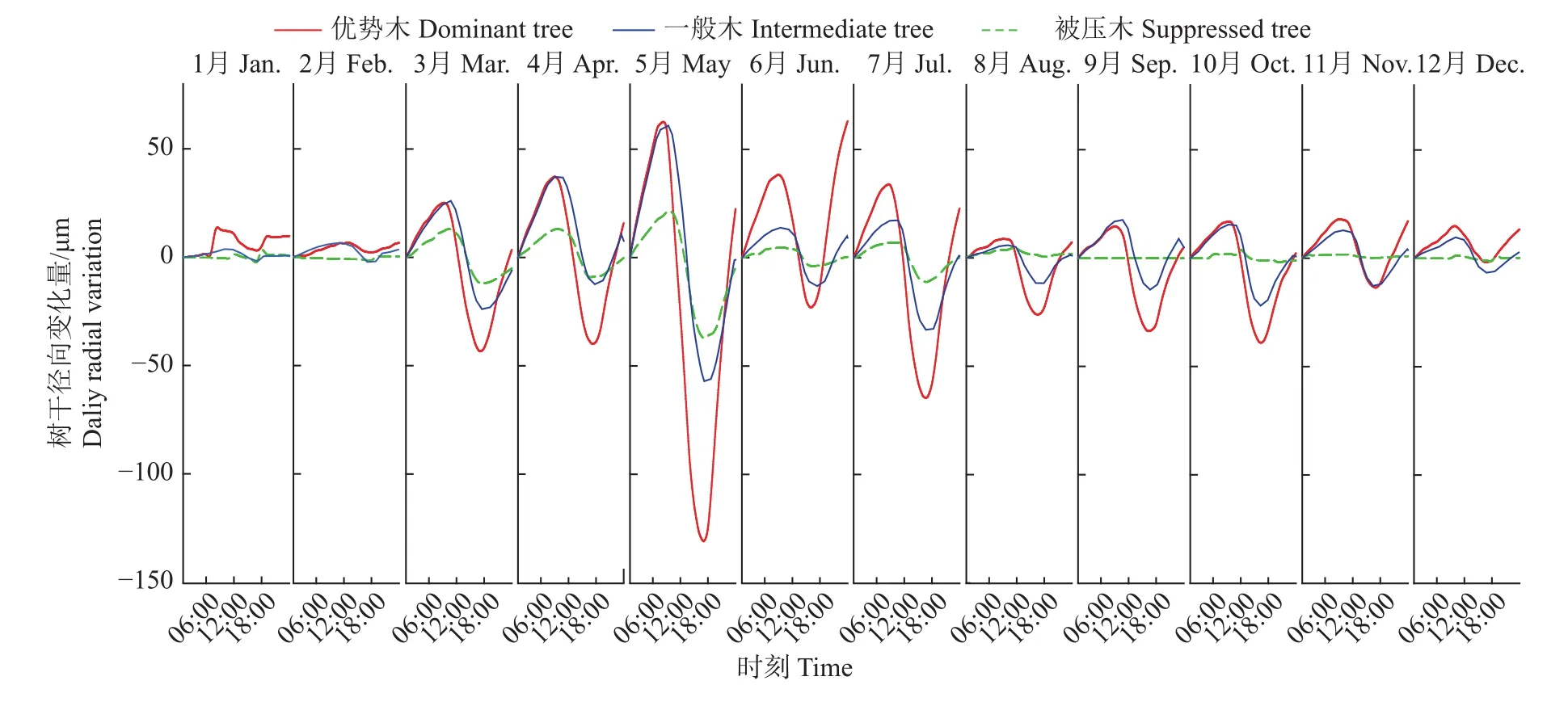
图1 红椎树干径向生长的各月平均日变化Fig.1 Monthly mean diurnal changes of stem radial growth ofCastanopsis hystrix
3.2 树干径向生长季节变化特征
利用Gompertz生长模型对红椎树干径向累积生长进行拟合(图2a),该模型解释了57%~99%的红椎树干径向累积生长变化,其径向累积生长表现为“S”型曲线,树干径向累积生长量上限值表现为优势木最大 (7138 μm),一般木次之 (2466 μm),被压木最小(267 μm)。从红椎树干径向累积生长的季节动态变化来看,除被压木径向累积生长的动态变化不明显外,样树树干径向累积生长的变化趋势基本一致,季节性径向变化的特点是春季(3月至5月上旬)径向逐渐生长,随后进入平台期(优势木)或径向生长明显减小(一般木和被压木),春末和初夏(5月下旬至7月中旬)径向快速生长,随后再次进入平台期(一般木和被压木)或径向逐渐生长(优势木),秋季(9月至11月)径向累积生长明显增大,之后开始稳定,除降雨引起了几次较大的波动,未观察到径向收缩现象。
红椎树干径向生长速度拟合曲线呈单峰型曲线(图2b),径向最大生长速度及其出现时间在不同样树间均明显不同,优势木、一般木和被压木树干径向生长的最大速度分别为 46.14、12.21 和 1.70 μm·d−1, 分别出现在一年中的第146、163和190天。3棵样树生长状况表明,优势木主要生长时期为一年中的第59 —331天,一般木为第73 — 317天,被压木没有主要生长时期,即优势木的主要生长时期开始时间比一般木早 14 d,结束时间比一般木晚 14 d。
3.3 树干径向生长变化对环境因子的响应
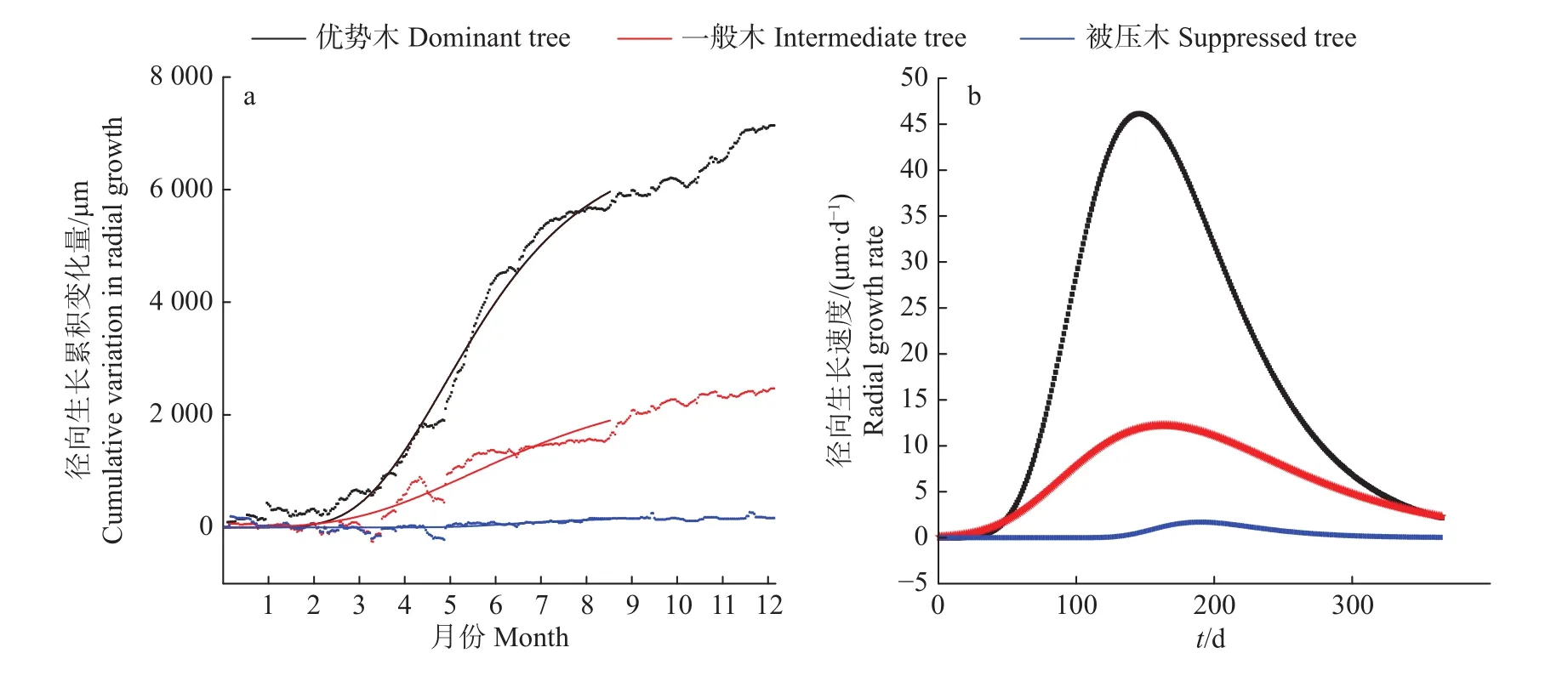
图2 红椎树干径向生长的累积变化及生长速度拟合Fig.2 Curve fittings of cumulative variation in stem radial growth and radial growth rate forCastanopsis hystrix
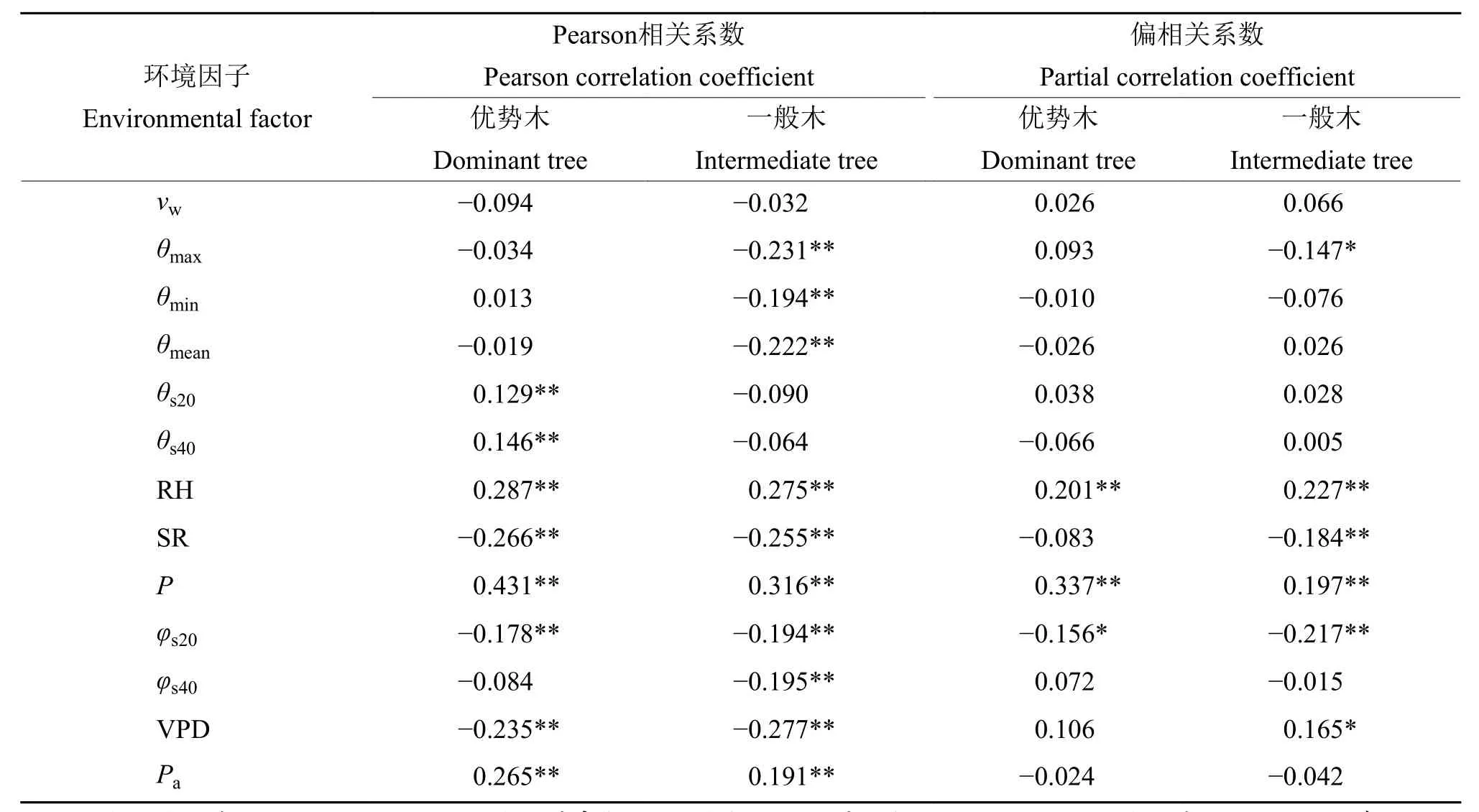
表2 红椎树干径向生长与环境因子的相关性分析1)Table 2 Correlation analyses of stem radial growth and environmental factors forCastanopsis hystrix
对红椎优势木和一般木主要生长时期的径向生长与气象环境因子的关系进行分析(表2),从Pearson相关系数来看,优势木树干径向生长与深度为20 cm土壤的日均温度、深度为40 cm土壤的日均温度、日降雨量、日均相对空气湿度和日均大气水势呈极显著正相关,与日均辐射、深度为20 cm土壤的日均含水率和日均饱和水汽压差呈极显著负相关,与日最高气温、日均气温、日最低气温、日均风速和深度为40 cm土壤的日均含水率呈正相关或负相关关系,但相关性不显著;一般木树干径向生长与日降雨量、日均相对空气湿度和日均大气水势呈极显著正相关,与日最高气温、日均气温、日最低气温、日均辐射、深度为20和40 cm土壤的日均含水率以及日均饱和水汽压差呈显著负相关;与日均风速、深度为20和40 cm土壤的日均温度呈负相关关系,但相关性不显著。
红椎优势木和一般木主要生长时期的气象环境因子的主成分分析结果(表3)显示,有3个特征值大于1的主成分,其累计贡献率分别为78.045%、77.991%,这3个主成分能够反映出其树干径向生长对气象环境因子的响应。从表3主成分分析中可以看出:第1主成分中,日空气温度、日均土壤温度的载荷较大,其次是日均辐射和日均饱和水气压差,它们反映了该地区的热量条件,与树木光合物质的形成、积累和代谢等生理过程相关,是影响树干径向生长的直接因子;第2主成分中载荷较大的是日均大气水势、空气相对湿度和饱和水汽压差;第3主成分中载荷较大的是日均土壤含水率和日降雨量,它们反映了该地区的水分条件,与树干形成层细胞的水分利用过程相关,是影响树干径向生长的关键因子。

表3 红椎主要径向生长时期的气象环境因子的主成分载荷及特征值和贡献率1)Table 3 Load eigenvalue and contribution rate for environmental factor during the major growth period ofCastanopsis hystrix
为进一步量化树干径向生长对各气象环境因子的响应关系,进行了偏相关分析(表2),从径向生长与环境因子的偏相关系数可知,影响不同分化等级样树树干径向生长变化的主导因子不同。优势木树干径向生长与日降雨量、日均空气相对湿度呈极显著正相关,与深度为20 cm土壤的日均含水率呈显著负相关,而一般木除与日降雨量、日均空气相对湿度、深度为20 cm土壤的日均含水率和日均辐射呈极显著正或负相关外,还与日最高气温、日均饱和水汽压差呈显著正或负相关。
综合考虑Pearson相关系数、主成分分析和偏相关分析结果,影响优势木树干径向生长的主要气象环境因子选择日均空气湿度、日均降雨量和深度为20 cm土壤的日均含水率3个因子,影响一般木树干径向生长的主要气象环境因子选择日均空气湿度、日均降雨量、日均辐射和深度为20 cm土壤的日均含水率4个因子,采用多元线性回归分析建立了影响红椎树干径向生长的主要气象环境因子模型,所有因子均与树干径向日生长量(SRI,μm)显著相关。优势木的树干径向生长模型表示为:

一般木的树干径向生长模型表示为:

4 讨论与结论
4.1 树干径向生长日变化特征
蒸腾作用、土壤水分状况、空气温度随时间的变化而变化,树干径向生长日变化的振幅和相位整合了对环境变化的生理反应的信号。本研究中,红椎树干径向生长月均日变化只有1种夏季模式,这与以往研究的落叶松Larix decidua[26]、云杉Picea crassifolia[27]、青杄Picea wilsonii和圆柏Sabina chinensis[23]的树干径向生长日周期模式有夏季和冬季2种模式的研究结果不同,其原因主要与研究区域有关,以往研究区域的年均气温为0.6~7.4 ℃、年均降雨量为419.0~825.0 mm,远低于本研究区域的年均气温 (20.5~21.7 ℃)和降雨量 (1 200~1 600 mm)。一般来说,夏季模式受水分条件控制,反映了水分在土壤、根系及茎干吸水和光合及蒸腾失水这一水分运输过程中的平衡状态[28],而冬季模式与空气温度密切相关。由此可见,水分是影响本研究区域红椎树干径向生长日周期变化的主要因素。
4.2 树干径向生长季节变化特征
在亚热带气候条件下,树干径向生长受暖冬和酷暑双重气候胁迫的制约[29-30]。红椎树干径向生长在过渡季节(春季−初夏和夏末−秋季)出现了2次生长高峰,在夏季生长速度下降。在高温环境条件下,叶片气孔导度较差,呼吸速率较高,降低了树干径向生长所需的碳水化合物供应,导致径向生长速度降低,夏季生长抑制是树木应对夏季高温及干旱等恶劣环境的一种生存策略[19]。秋季降雨可能会激活形成层细胞的生理活动,从而出现了秋季树干径向增加的第2个高峰[31-32]。
4.3 不同分化等级树木的生长反应
在相同的气候条件下,同一树种、同一年龄、生长在同一林分中的树木,主要生长时期因分化等级的不同而不同。本研究中,优势木的主要生长时期开始早于一般木,结束晚于一般木,被压木没有主要生长时期。Rathgeber等[33]对法国东北部不同径级冷杉Abies alba树干形成层细胞生长的研究表明,优势木细胞开始生长的时间比一般木早8 d,比被压木早21 d;优势木细胞停止生长的时间比一般木晚 11 d,比被压木晚 49 d,支持了本研究的结果。优势木主要生长时期开始较早的原因可能在于,春季较高树的树冠顶部在早晨较早和下午较晚被阳光直射而变暖,从而在本季节早些时候达到触发快速生长的临界温度水平。树冠较大的树木受益于更多的资源和更强的光合作用能力,使韧皮部含有较高浓度的同化物质,并向形成层输送更多的碳水化合物,使其达到较高的径向生长水平,红椎树干径向累积生长量表现为优势木最大,一般木次之,被压木最小。Kozlowski等[34]对美国威斯康辛州中部的红松Pinus koraiensis形成层生长的研究表明,优势木、一般木和被压木形成层生长的季节速率不同,优势木生长最快,被压木生长缓慢,这与本研究中的优势木树干径向生长最大速度可达46.14 μm·d−1,一般木为 12.21 μm·d−1,而被压木仅为 1.70 μm·d−1的结果不谋而合。此外,本研究表明,优势木、一般木和被压木树干径向生长最大速度分别出现在5月末、6月中旬和7月上旬。一些学者在研究中也有类似的发现[35-36],这主要是因为光周期对树木生长和植物的生理过程的影响[37-38],为抵御冬季低温,树木会在光周期相对较长的生长季前期快速生长以产生大量的生长激素[39],促进形成层完成大部分细胞的分裂和伸长生长,从而在生长季后期有足够的时间完成新生细胞的细胞壁木质化。
4.4 树干径向生长变化对主要环境因子的响应
以往研究结果表明,树干径向生长量主要与日降雨量、日均相对空气湿度显著正相关,与日均土壤含水率、日均辐射显著负相关[27,40-41],这与本文研究结果基本一致。降雨导致叶片水势降低,水通过渗透作用进入形成层细胞促进细胞伸长生长[42],且降雨提高了土壤含水量,利于根系吸水使形成层细胞水分含量增加[36]。空气相对湿度、辐射通过影响树木蒸腾作用影响径向生长变化[43]。空气湿度大,蒸腾作用减弱,降低了水运系统的负压,促进形成层细胞膨胀;辐射越大,蒸腾作用越强,从而抑制形成层细胞膨胀[42]。此外,后者可导致气孔关闭,影响光合作用,进而抑制径向生长[44]。
在生长−气象模型中包含日常数据可能有助于提高潜在的树木生长对气候的反应的预测,为此许多学者进行了研究[3,19,27,41]。然而,不同分化等级树木的主要生长时期差异会导致树木在生命、功能和生长发育能力等许多方面产生差异,可能会有不同的气候−生长关系,现有研究中,鲜有建立不同分化等级林木生长−气象生长模型。本研究通过对13个环境因子进行主成分分析和偏相关分析,分别建立了影响优势木、一般木径向日生长量的多元线性回归模型,这是探索径向生长与气象因子之间关系的新尝试。
致谢:感谢中国林业科学研究院资源信息所雷相东研究员对论文修改过程的指导!
