皇竹草竞争抑制入侵植物假臭草的潜力
2020-08-05许秋园全国明章家恩李赛飞
许秋园,王 丹,全国明,章家恩,李赛飞
(1 华南农业大学 资源环境学院,广东 广州 510642;2 广东省生态循环农业重点实验室/广东省现代生态农业与循环农业工程技术研究中心/农业部华南热带农业环境重点实验室,广东 广州 510642;3 广州城市职业学院 城市建设工程系,广东 广州 510405)
替代控制是一种应用前景广阔的入侵杂草控制方法。该方法根据种间竞争和群落自然演替的规律,通过人为引入具有竞争力的植物,减少甚至清除群落中的有害入侵植物[1]。竞争力强的本地植物是用于替代控制外来入侵植物的理想选择,然而本地植物中竞争能力高于入侵植物的种十分有限,成功的实践更是鲜有报道。因此,研究者们开始尝试利用入侵潜力较低的外来物种替代控制入侵植物[2-4]。Kuebbing等[5]通过分析前人的研究发现,在外来植物之间的相互作用类型中,竞争性相互作用十分常见。种间竞争作用可以阻止入侵植物在邻近群落中定植,从而在一定的空间尺度内限制其扩散[6]。种间竞争在外来物种之间的普遍性及其竞争效果为利用外来植物控制入侵植物提供了理论基础。植物通过改变性状与相邻的植物竞争有限的自然资源(如光照、水分和土壤养分等)[6],方式包括通过吸收和利用公共资源的直接竞争和通过阻碍相邻植物获取资源的间接竞争[7-8]。因此,种间竞争的结果,同时取决于植物对相邻植物的抑制能力及其耐受抑制作用的能力2个方面[9]。由于物种的抑制能力和耐受能力并不必然具有负相关或正相关关系[6],而且入侵物种大多对相邻的植物具有化感作用,因此在选择用于替代控制的植物时应同时考察其对目标植物的抑制能力及耐受能力。
假臭草Praxelis clematidea是原产于南美洲的1年生菊科草本植物,于20世纪80年代首次出现在香港,近年来已大面积入侵华南地区,占据道路两旁的空地、荒草地、果园和农田[10],目前对其还没有生态高效的防治清除方法。该植物喜较湿润及阳光充足的环境。皇竹草Pennisetum hydridum是一种多年生直立丛生禾本科植物,由象草Pennisetum purpureum和美洲狼尾草Pennisetum typhoideum杂交选育而成[11],入侵风险极小[12]。皇竹草作为一种植株高大、生长快速的复合型经济作物,在替代控制该入侵杂草方面可能具有较大的潜力。因此,为了检验使用皇竹草替代控制假臭草是否具有可行性,同时为了检验皇竹草作为替代控制工具的竞争机制和优势,本研究以假臭草和皇竹草作为对象设置了不同比例的混种盆栽试验。
1 材料与方法
1.1 试验材料与试验设计
分别收集假臭草种子和皇竹草新鲜茎段进行育苗。苗床统一使用按3∶1的体积比混合均匀的有机水藓基质泥炭土和珍珠岩(丹麦品氏)作为基质。假臭草的种子采自广州市华南农业大学增城教学科研基地,经40 ℃温水浸泡24 h后均匀撒播于苗床中自然萌发;皇竹草茎段采集自华南农业大学资源环境学院教学科研基地附近,分节后扦插到苗床中。
育苗约3周后,选择生长健壮、株高18~30 cm的幼苗分别将皇竹草和假臭草按1∶3、2∶2和3∶1的数量比例移栽到塑料盆中,每盆共种植4株幼苗,每个处理5个重复(盆)。另设皇竹草和假臭草单种(即二者数量比例为4∶0和0∶4)作为对照。花盆规格为 23.0 cm(上口径)×15.0 cm(下口径)×21.0 cm(高),每盆装土5 kg。试验用土采自华南农业大学资源环境学院教学科研基地附近未被外来植物入侵的荒坡地表层土。采集的土壤经充分混匀并过5 mm筛后再填装入盆,土壤全氮质量分数0.86 g·kg−1、全磷质量分数 0.62 g·kg−1、全钾质量分数25.14 g·kg−1、碱解氮质量分数 384.18 mg·kg−1、有效磷质量分数 37.73 mg·kg−1和有效钾质量分数152.34 mg·kg−1。种植 25 周后,对植物形态特征进行测量,并采集整株植物测定生物量。
1.2 植物生物量分配及形态性状的测定
用直尺测定每株植物的枝条长度,计算分枝数。收集植株的所有叶片,使用扫描仪器扫描后用Adobe Photoshop CS2 9.0.2 (Adobe Systems, 美国)统计叶面积。所有植株的根、茎、叶等器官分离后于70 ℃烘箱中烘72 h,达到恒质量后用电子天平称质量,得到每棵植株的根系及地上部生物量。对根系及地上部生物量求和得到单株生物量。比叶面积(SLA)= 总叶面积/叶片干质量。根冠比为根系与地上生物量的比值。其中,单株生物量、各器官生物量、总叶面积、总茎长、分枝数和比叶面积均表示单棵植株的对应指标,盆中有多棵植物时取相应指标的平均值代表1个重复。
1.3 植物生长指标的计算和数据分析
计算相对产量(RY)、总相对产量(RYt)和竞争平衡指数(CB)[13]:

式中,YH和YJ分别为皇竹草和假臭草在单种时每盆产量的平均值,YHJ和YJH分别为二者在不同处理中每盆的产量,RYH和RYJ分别为二者的相对产量;q和p分别为皇竹草和假臭草在某个处理中的种植占比(q+p=1),当皇竹草和假臭草比例为4∶0、3∶1、2∶2、1∶3 和 0∶4 时,q分别为 1.00、0.75、0.50、0.25和 0,p分别为 0、0.25、0.50、0.75 和 1.00。RYt<1表明2种植物种间竞争作用强于种内竞争作用;RYt>1 表明情况相反;RYt=1 表明种间与种内竞争的强度基本持平。CBJ和CBH分别为二者的竞争指数,由于CBJ= −CBH,本文只计算假臭草的竞争平衡指数CBJ来表示2个物种之间的竞争关系。CBJ> 0表明假臭草在竞争中处于优势;CBJ=0表明2个物种竞争能力相等;C BJ<0表明物种假臭草在竞争中处于劣势;CBJ的绝对值越大,说明2种植物的竞争能力差距越大。根据种植比例和单种产量可得到植物在每个处理中的理想相对产量(RYe),在混种比例为 4∶0、3∶1、2∶2、1∶3 和 0∶4 时,皇竹草的RYe分别为 1.00、0.75、0.50、0.25 和0,假臭草的RYe则为 0、0.25、0.5 0、0.75和1.00。
采用单一样本t检验分别比较RY与RYe、RYt与1及CB与0之间的差异;皇竹草和假臭草在单种与混种处理间各性状的种内差异采用单因素方差分析及LSD多重比较进行检验,皇竹草和假臭草观测值与预测值的差异采用t检验进行比较。所有分析在 R 软件 (Rver.3.5.3)[14]中完成。
2 结果与分析
2.1 皇竹草和假臭草的形态差异
单作条件下,皇竹草植株体型远大于假臭草。皇竹草的单株生物量、地下部生物量及地上部生物量均显著高于假臭草,分别约为后者的8、24和6倍(P< 0.01,表1)。这表明皇竹草比假臭草具有更高的资源利用能力。另一方面,皇竹草的根冠比也显著大于假臭草,是假臭草根冠比的3.7 7倍(P=0.009,表1),而单株总茎长、分枝数和比叶面积均显著小于假臭草(P< 0.001,表1),表明2种植物具有不同的资源获取策略。
表1 皇竹草和假臭草单作条件下的单株生物量及形态特征(±SE,n=5)Table 1 Biomass per plant and morphological traits forPennisetum hydridum andPraxelis clematidea under monoculture
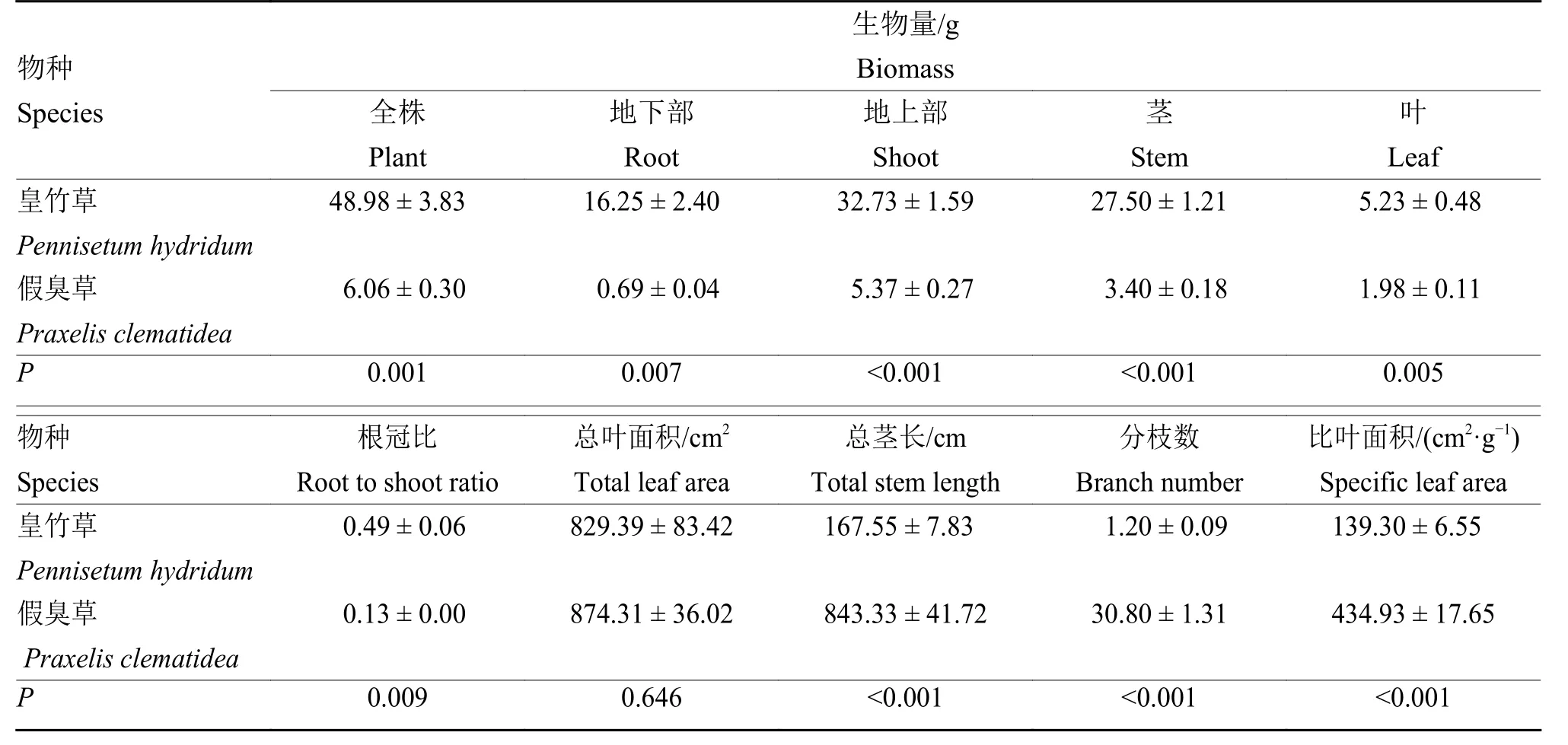
表1 皇竹草和假臭草单作条件下的单株生物量及形态特征(±SE,n=5)Table 1 Biomass per plant and morphological traits forPennisetum hydridum andPraxelis clematidea under monoculture
物种Species 茎叶Leaf皇竹草Pennisetum hydridum 48.98 ± 3.83 16.25 ± 2.40 32.73 ± 1.59 27.50 ± 1.21 5.23 ± 0.48 假臭草Praxelis clematidea 6.06 ± 0.30 0.69 ± 0.04 5.37 ± 0.27 3.40 ± 0.18 1.98 ± 0.11P 0.001 0.007 <0.001 <0.001 0.005物种Species比叶面积/(cm2·g−1)Specific leaf area皇竹草Pennisetum hydridum根冠比Root to shoot ratio总叶面积/cm2 Total leaf area总茎长/cm Total stem length分枝数Branch number 0.49 ± 0.06 829.39 ± 83.42 167.55 ± 7.83 1.20 ± 0.09 139.30 ± 6.55 假臭草Praxelis clematidea 0.13 ± 0.00 874.31 ± 36.02 843.33 ± 41.72 30.80 ± 1.31 434.93 ± 17.65 P 0.009 0.646 <0.001 <0.001 <0.001生物量/g Biomass全株Plant地下部Root地上部Shoot Stem
2.2 皇竹草和假臭草的竞争关系
由图1可知,皇竹草种内竞争激烈,2种植物的种内及种间竞争关系随着混种比例的变化而变化。皇竹草和假臭草混种比例为2∶2和1∶3处理的RYt显著大于 1.00,分别为 1.27(P= 0.009)和1.80(P= 0.008),表明混种处理中皇竹草的种内竞争作用强于种间竞争作用。在所有处理中,假臭草的相对产量都与预期值无显著差异,表明假臭草的种内竞争和种间竞争相对平衡。在混种比例为2∶2和1∶3时,皇竹草的相对产量显著高于预期值0.50 和 0.25,分别为 0.89(P< 0.001)和 0.83 (P=0.002),观测值与预期值的差距随着种植比例的降低而增高,表明皇竹草种内竞争较强,混种处理缓解了皇竹草的种内竞争。所有混种处理中的假臭草的竞争平衡指数均小于0,范围为−0.97~−0.40,其中混种比例为2∶2和1∶3时的竞争平衡指数与0差异显著,分别为−0.79(P= 0.020)和−0.97(P=0.030),表明在与皇竹草的竞争中,假臭草处于劣势。而且假臭草的竞争平衡指数随着皇竹草种植比例降低而降低,说明皇竹草的种间竞争能力随着自身密度的降低而增强。
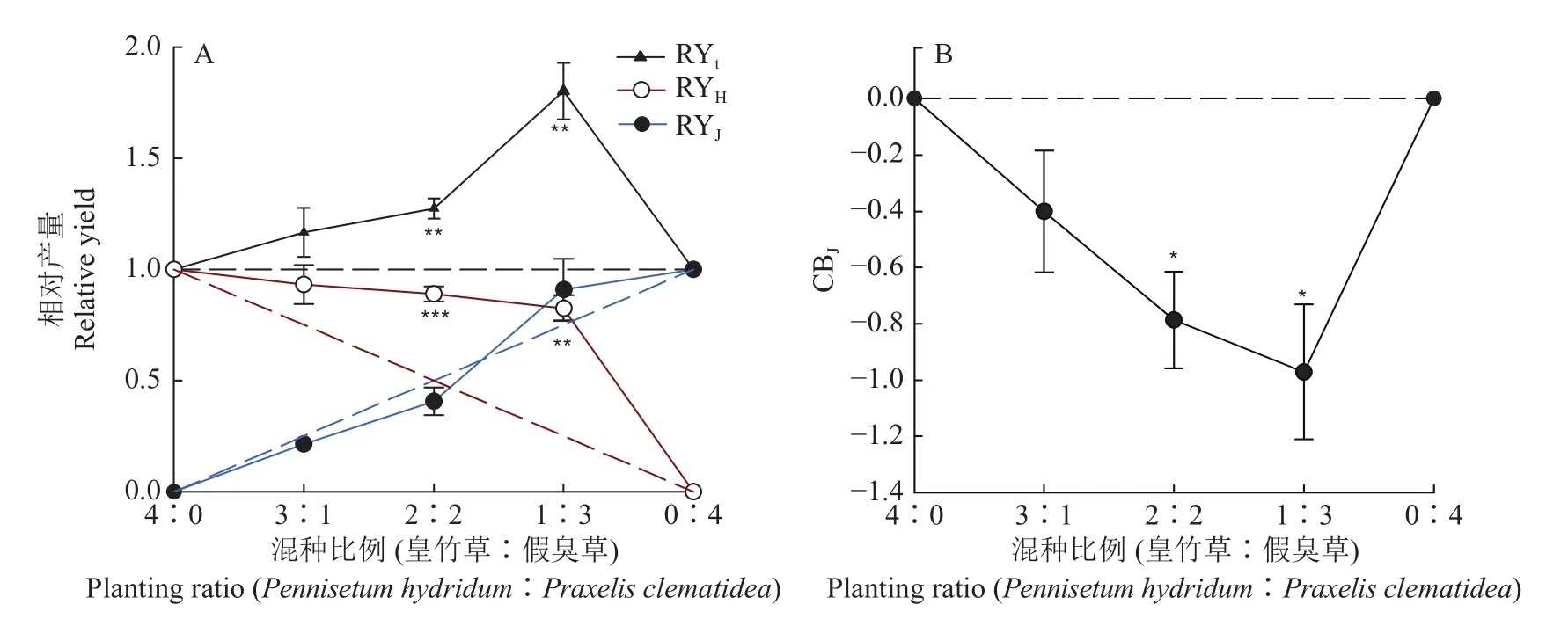
图1 不同混种比例下皇竹草和假臭草的相对产量(RYH和RYJ)、总相对产量(RYt)以及假臭草的竞争平衡指数(CBJ)Fig.1 The relative yield ofPennisetum hydridum andPraxelis clematidea (RYH and RYJ), the total relative yield (RYt)and the competitive balance index (CBJ)forPraxelisclematidea under different planting ratios
2.3 混种比例对2种植物生物量分配的影响
单因素方差分析显示2种植物的混种显著影响了皇竹草的单株、地下部、地上部、茎、叶生物量及根冠比(P< 0.05),但只对假臭草地下部生物量和茎、叶生物量有显著影响 (P< 0.05, 表 2)。皇竹草的单株生物量、地下部、地上部生物量及茎、叶生物量均随着自身种植比例的下降发生不同程度的增加,在自身种植比例为0.25 (皇竹草和假臭草混种比例为1∶3)时达到最大值,分别为单种时的3.3、3.8、3.1、3.0 和 3.1 倍 (P< 0.05,图 2),其单株生物量自单种时的48.98 g增加至161.66 g(混种比例为1∶3)。皇竹草的根冠比在混种比例为3∶1时是单种时的1.6倍(P< 0.05),但混种比例为2∶2和1∶3时只略为升高,与单种没有显著差异。假臭草的单株生物量和地下部生物量在自身种植比例为0.25(混种比例为3∶1)时最高,分别为单种时的1.5和1.6倍(P< 0.05)。其地上部生物量无显著变化,但混种处理中茎生物量比单种高,叶生物量比单种低,其中茎生物量最高时(混种比例为3∶1)是单种时的2.0倍,叶生物量最低时(混种比例为2∶2)是单种时的 0.4 倍 (P< 0.05)。混种处理中,假臭草的根冠比与单种中的均无显著差异,但在混种比例为2∶2和3∶1时,其根冠比显著高于混种比例为1∶3的处理(P< 0.05)。表明混种比例显著影响了2种植物的生物量积累及分配。随着自身种植比例下降,皇竹草生物量积累大幅增加,且通过增加地下部生物量的分配适应种间竞争压力的上升;假臭草生物量积累有小幅上升,并通过增加茎生物量的分配适应种间竞争压力的上升。
2.4 混种比例对2种植物形态的影响
混种比例不仅影响了植物的生物量分配,还影响了植物的其他形态指标。方差分析显示,混种比例显著影响了皇竹草的总叶面积、总茎长和比叶面积(P< 0.05),对分枝数无显著影响;混种比例同样显著影响了假臭草的总叶面积、总茎长和分枝数(P< 0.05),但对其比叶面积无显著影响(表2)。皇竹草在混种处理中的总叶面积显著低于单种,且在皇竹草和假臭草比例为2∶2时最低,其比叶面积在混
种比例为2∶2时显著低于单种,总茎长度在混种比例为2∶2和1∶3时显著大于3∶1混种,分枝数则随着混种比例的降低而降低(P< 0.05,图3)。表明随着种内竞争缓和、种间竞争加剧,皇竹草适应性地减少了自身分枝数并延长了茎的长度。假臭草在混种处理中总叶面积、总茎长和分枝数显著低于单种(P< 0.05),和单种相比,混种处理中的假臭草总叶面积减少43.4%~68.2%、总茎长减少49.4%~ 63.6%、分枝数减少46.1%~71.6%(图3)。表明在混种处理中,皇竹草和假臭草的叶片生长和分枝的形成均受到了抑制,但皇竹草通过延长茎的长度适应种间竞争。假臭草的总叶面积、总茎长和分枝数均显著减小,表明其地上生物量没有显著变化是枝条加粗、茎生物量增加的结果。
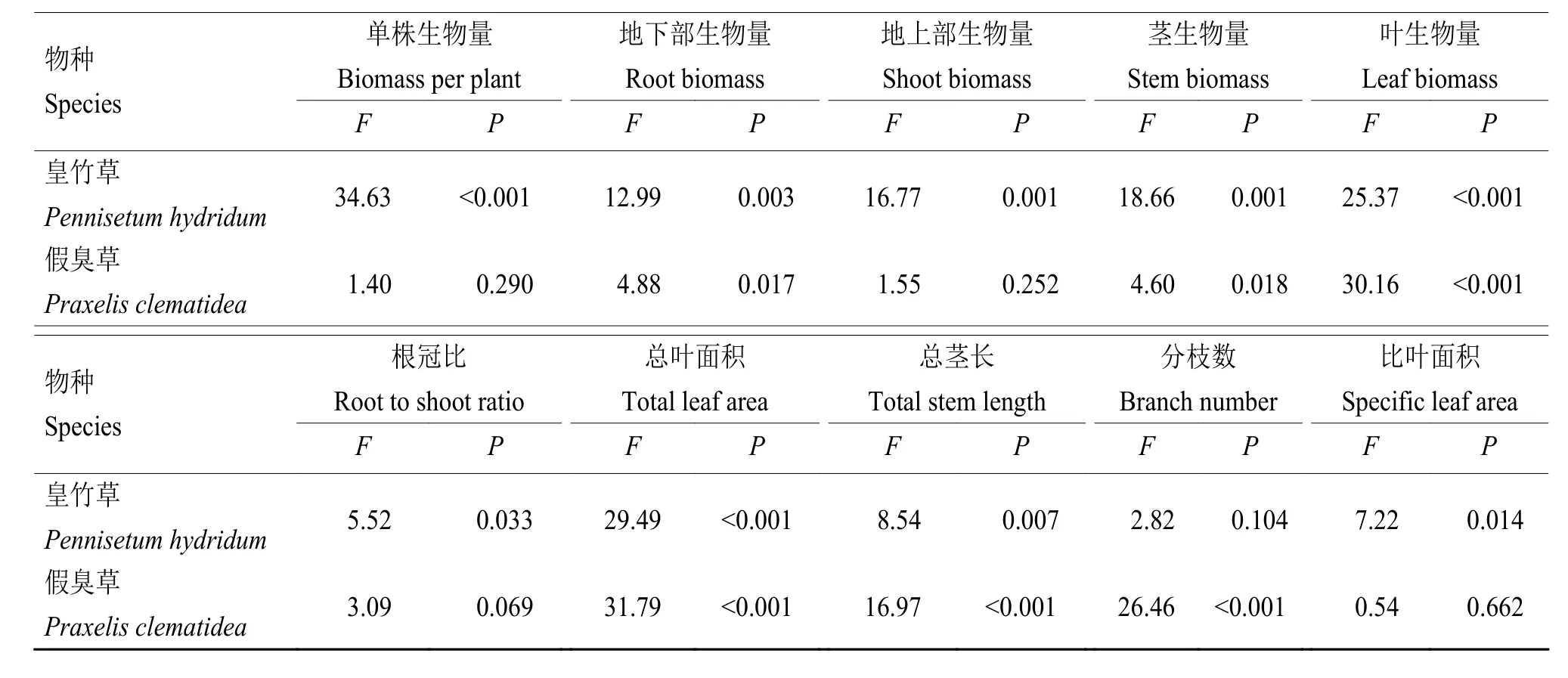
表2 皇竹草和假臭草混种对其单株生物量及形态特征影响的单因素方差分析(n = 5)Table 2 Effects of planting ratio on biomass per plant and morphological traits ofPennisetum hydridum andPraxelis clematidea analyzed by one-way ANOVA

图2 不同比例混种处理中皇竹草和假臭草的单株生物量分配(±SE,n = 5)Fig.2 Biomass distributions per plant ofPennisetum hydridum andPraxelis clematidea in treatments of different planting ratios

图3 不同比例混种处理中皇竹草和假臭草的形态特征(±SE,n = 5)Fig.3 Morphological characteristics ofPennisetum hydridum andPraxelis clematidea in treatments of different planting ratios
3 讨论与结论
植株高大的植物在光照竞争上往往较有优势,同时强壮的根系可使植物在土壤养分竞争中较易占据优势地位[6]。由于具有这些优势,皇竹草等高大且入侵风险低的植物常被作为控制入侵害草的优良候选植物[15–17]。本研究结果显示,随着竞争者假臭草的种植比例增大,皇竹草减少了总叶面积和分枝,同时延长了茎长,从而增加光照的截获,加强对假臭草的遮阴抑制。这是植物通过波长变化感受到相邻植物的存在后,调整生物量分配以增强光竞争能力的结果[7-8]。尽管假臭草与高大的皇竹草在光资源竞争中处于明显劣势,但随着皇竹草的种植比例增加,假臭草的生物量积累并没有受到明显的抑制。这可能是由于假臭草面对在光竞争方面毫无机会的对手时,调整了自身的竞争策略[8],其根冠比的变化及总叶面积和茎长的减小正说明了这一点。一般而言,植物生物量的分配与植物地上地下的竞争能力息息相关,例如根冠比随着光照降低而降低、随着养分和水分减少而增加[18]。面对过于高大的竞争者时,植物的叶片和茎的生长可能受到抑制,转而选择忍耐遮阴的策略,植物长期采取这种策略将导致有机物积累减少、竞争能力下降[8]。本研究发现,随着皇竹草的种植比例上升,假臭草选择了耐受遮阴并增强地下部分竞争能力的策略。皇竹草不仅在光竞争中表现出优势,而且表现出对假臭草化感作用的耐受性。有研究表明,假臭草能分泌化感物质,对其他杂草有明显的排斥作用,尤其对种子萌发的抑制作用明显[19]。一方面本研究发现皇竹草生物量积累不仅没有因假臭草种植比例增加而受到抑制甚至有所促进,另一方面皇竹草以种茎繁殖可避免其萌发受到假臭草的化感抑制[12]。
前人研究发现,光照较强的生境中植物的比叶面积较小[20-21],另一方面比叶面积与环境可获取水量具有正相关关系[22]。本研究中,皇竹草比叶面积的减少可能是光竞争缓和的结果,也可能是该混种比例(2∶2)下土壤水分竞争剧烈所导致的。有研究者在柱花草Stylosanthes guianensis替代控制假臭草的研究中发现,柱花草需要较高的植株比例才能有效抑制假臭草生长[23]。但在本研究中,由于皇竹草种内竞争较剧烈,虽然在所有处理中皆处于竞争优势地位,但在混种比例较低时(1∶3)竞争优势更为明显,因此在利用皇竹草的竞争优势控制假臭草时,应注意适当降低其自身的种植密度。
综上,皇竹草在与入侵杂草假臭草的竞争中处于优势地位,能影响假臭草的生物量分配,并且可耐受假臭草的化感作用[19],具有替代控制该入侵杂草的潜力。但皇竹草的种内竞争较强,故在野外应用时需控制好其自身的种植密度,以免影响其本身的产量及对假臭草的控制效果。
