拟南芥T-DNA 插入株系SALK_037453 叶发育突变位点的鉴定和分析
2020-07-29司旭阳李文静张洪艳潘延云
司旭阳,李文静,张洪艳,张 科,潘延云
(河北农业大学 河北省植物生理与分子病理学重点实验室,河北 保定 071000)
基因突变是研究基因功能的最重要策略之一[1-2]。 突变体可以通过网站T-DNA Express(http://signal.salk.edu/cgi-bin/T DNA Express)订购。但是,通过转基因技术获得的T-DNA 插入基因组的拟南芥,插入的T-DNA 拷贝数未必是1 个,而网站提供的信息只是其中1 个的T-DNA 插入位点。目标基因的突变体的表型是否与该基因连锁,则需要进一步证实[3-5]。
肌醇磷脂依赖的磷脂酶C(PI-PLC)是肌醇磷脂信号系统中的关键酶之一,拟南芥中有9 个不同的亚型,不同的亚型可能具有不同的生物学功能[6-7]。本实验室为研究拟南芥AtPLC3亚型的功能,购得2 株T-DNA 插入AtPLC3不同位点的突变体SALK_037453(本室命名为atplc3-1)和SALK_054406(本室命名为atplc3-2),网站给的基因注释显示,这2 个突变体的T-DNA 均分别插入到AtPLC3基因的不同的外显子区域。表型分析发现,这2 个突变体具有不同的表型,其中atplc3-1具有叶发育缺陷的表型,而atplc3-2却无此表型,意味着atplc3-1的表型可能与AtPLC3基因不连锁,即SALK_037453 突变体中可以存在其他的T-DNA 插入位点。拟南芥莲座叶的“聚集”是发育表型之一,为了确认造成该发育异常的突变基因,本研究利用TAIL-PCR 的方法,分析鉴定SALK_037453 突变株中其他T-DNA 插入位点的基因序列,并确认该突变位点与叶发育表型的相关性,将为基因转录表达调控提供新的调控序列,并为拟南芥叶片发育机制的研究提供新的候选基因和突变体材料。
1 材料与方法
1.1 试验材料及试剂
野生型拟南芥(Arabidopsis thaliana)生态型为Columbia(Col),本实验室保存,SALK_037453和SALK_054406 购 自Salk Institute Genomic Analysis Laboratory。dNTP 和Taq DNA 聚合酶为 TaKaRa 公司产品;植物RNA 快速提取试剂盒来自Aidlab,反转录试剂盒以及荧光定量试剂和其他用品来自Vazyme;测序由华大生物工程(北京)有限公司完成。
1.2 拟南芥的种植及基因组DNA 的提取
将拟南芥的种子表面灭菌后播种在1/2 MS 培养基,冰箱4 ℃放置3 d 后,在22 ℃ 16 h 光照和8 h 黑暗条件下培养。待幼苗长出真叶后移栽到土培 培养基(营养土∶蛭石=1∶3)中培养。
剪取拟南芥幼苗叶片,以CTAB 法提取基因组DNA,用于突变体纯合植株的鉴定和TAIL-PCR 分析。
1.3 拟南芥突变体纯合体的PCR 鉴定
以1 μL 拟南芥突变体基因组DNA 为模板,选取T-DNA 插入位点两侧的基因序列设计引物(LB和RB),利用T-DNA 左臂引物LBb1.3 与LB 引物进行PCR,鉴定T-DNA 插入突变纯合体。如果只有LB 和LBb1.3 能扩增出目的条带,则为T-DNA插入纯合体。PCR 鉴定引物见表1。

表1 突变体PCR 鉴定引物Table 1 Primers for mutant PCR identification
1.4 RT-PCR 反应
取生长10 d 的拟南芥幼苗,根据试剂盒说明提取RNA,反转录成cDNA 作为扩增的模板,RTPCR 的引物见表2。
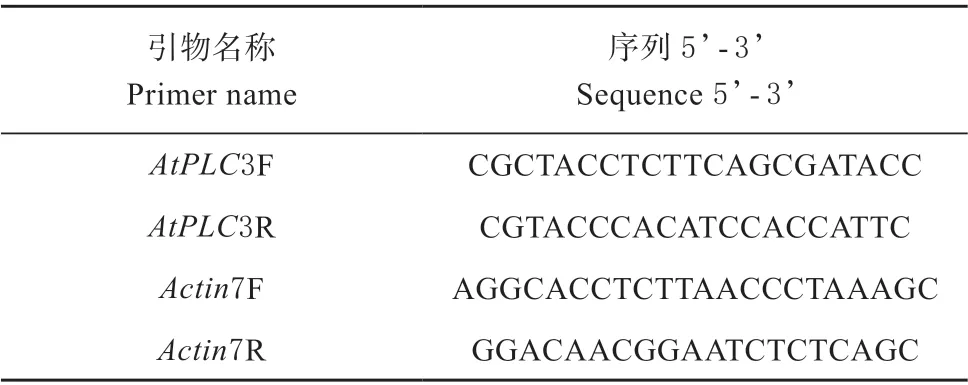
表2 RT-PCR 引物序列Table 2 Primers for RT-PCR
1.5 TAIL-PCR 扩增T-DNA 旁侧序列
T-DNA 的序列见http://signal.salk.edu/cgi-bin/T DNA Express,根据T-DNA 序列设计TAIL-PCR 反应的特异性引物见表3。另外3 条随机引物见表4。TAIL-PCR 反应体系和反应条件参考文献[8]。TAIL-PCR 获得的DNA 片段进行1%的琼脂糖凝胶电泳,将大小合适的条带送测序公司测序。
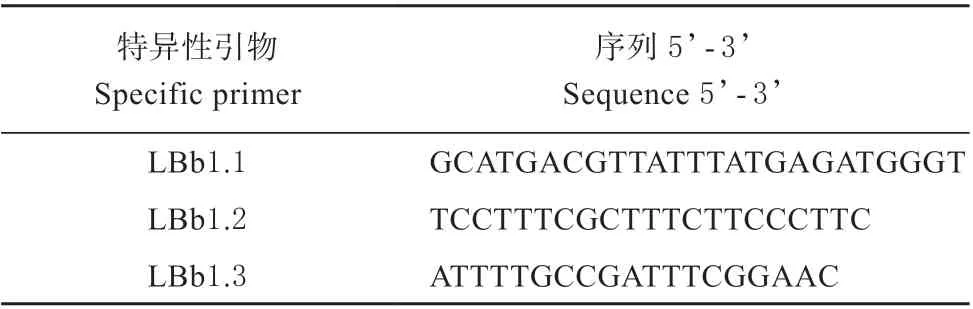
表3 TAIL-PCR 反应的特异性引物序列Table 3 Specific primer for TAIL-PCR reaction
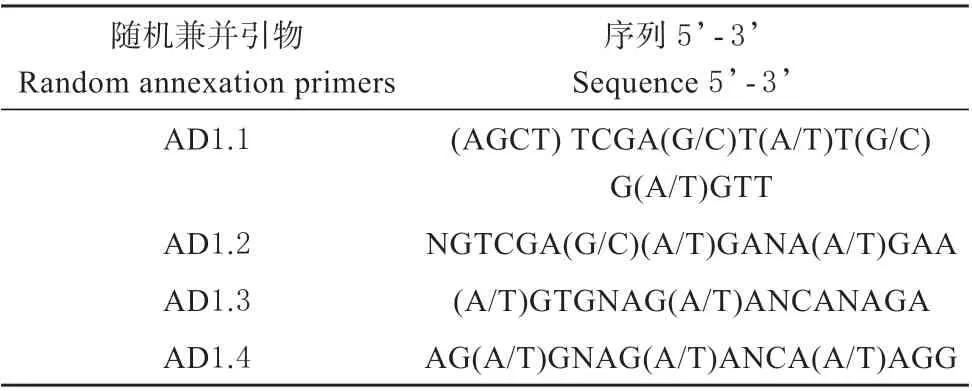
表4 TAIL-PCR 反应的随机兼并引物序列Table 4 Random annexation primer sequence for TAILPCR reaction
1.6 顺式作用元件分析
用 PLACE(http://www.dna.affrc.go.jp/PLACE/)和Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)2 个数据库分析序列的顺式作用元件。
1.7 实时定量 PCR 检测AtBRI1 和AtPLS 基因 的表达
选取生长28 d 的幼苗,利用EASYspin Plus 植物RNA 快速提取试剂盒提取总RNA,进一步利用HiScript ® III Reverse Transcriptase 反转录为cDNA,用于实时定量PCR 的检测。实时定量PCR 的引物为:AtBRI1-F:5’TGTTCTGAGCTGGTTTCGCT 3’AtBRI1-R:5’AGAGCCTAAGCTCGAAGGGA 3’AtPLS-F:5’GGAACACGAAATCCGAAGAGC 3’AtPLS-R:5’CGCCAACATCACCACAACAAG 3’; 内参基因引物:Actin2-F:5’CGTACAACCGGTATTGTGCT 3’;Actin2-R:5’GATGTCTCTTACAATTTCCCGCT 3’,qRT-PCR 条件为:95 ℃ 5 min;95 ℃ 10 s;60 ℃ 30 s (40 个循环)。
2 结果与分析
2.1 SALK_037453 突变体中未知序列的T-DNA 插入造成叶片发育缺陷
获得SALK_037453 和SALK_054406 突变体后,利用方法1.3 鉴定和筛选2 种突变体的纯合体。结果显示,与网站提供的注释相同,SALK_037453的T-DNA 插入到第4 个外显子区,破坏了AtPLC3基因的结构,命名为atplc3-1,SALK_054406 的T-DNA 插入到第3 个外显子区,命名为atplc3-2(图1A);RT-PCR 分析显示,2 个不同位点的插入均破坏了AtPLC3基因的转录表达(图1B)。
进一步对2 种突变体进行培养和表型观察,结果发现,atplc3-1表现出莲座叶聚集的发育表型,而atplc3-2与野生型拟南芥相同(图2A)。该表型可能不是由于AtPLC3基因的表达缺失引起的,atplc3-1中可能还存在其他位点的T-DNA 插入,进而导致某未知基因的突变,引起了该叶发育缺陷的表型。为了进一步确认该结论,首先将plc3-1与Col 杂交,发现在F2 代中无T-DNA 插入的AtPLC3植株(命名为SALK_037453X)中有相同的叶发育缺陷表型(图2A),并且利用T-DNA 引物序列对SALK_037453X 的基因组DNA 进行PCR 扩增,获得阳性结果(图2B)。证实SALK_037453X 中存在其他的T-DNA 插入位点,并且该插入可能影响了某未知基因的功能,从而对叶片发育造成影响。因此利用TAIL-PCR 检测新的插入位点,将新插入位点命名为X。
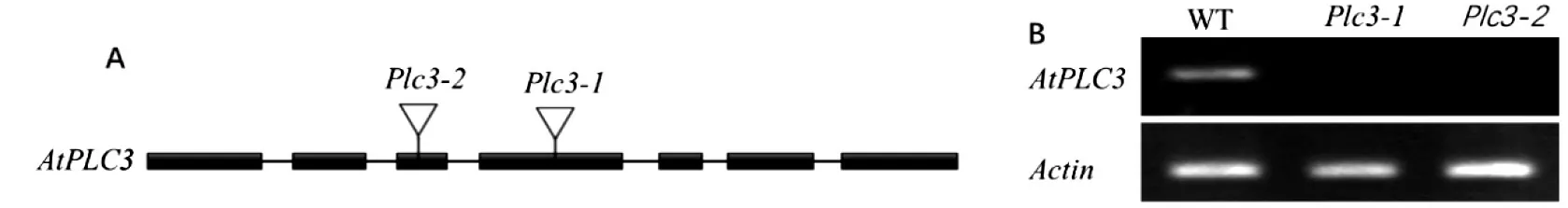
图1 SALK_037453(atplc3-1)和SALK_054406(atplc3-2)的T-DNA 插入位点(A)及AtPLC3 基因的表达(B)Fig.1 T-DNA insertion sites of SALK_037453 (atplc3-1), SALK_054406 (atplc3-2) (A) and the expression of AtPLC3 gene (B)

图2 SALK_037453(atplc3-1),SALK_054406(atplc3-2)和SALK_037453X 的叶发育表型(A)及T-DNA 序列检测(B)Fig.2 Leaf development phenotypes of SALK_037453 (atplc3-1), SALK_054406 (atplc3-2) and SALK_037453X (A) and the detection of T-DNA sequence (B)
2.2 利用 TAIL-PCR 捕获SALK_037453X 中未知T-DNA 插入位点X
本试验中以SALK_037453X 拟南芥基因组DNA 为模板,利用TAIL-PCR 技术以3 个T-DNA序列中的特异引物和4 个随机兼并引物扩增T-DNA侧翼序列,发现特异引物分别与AD1,AD2,AD3组合条件下可以获得清晰的条带,且比第2 轮小300 bp(图3A),符合TAIL-PCR 的要求。切下符合要求的3 个(图3A 中箭头所指)条带并送测序,比对后发现3 条带的序列是相同的,进一步在拟南芥基因组中BLAST,获得了T-DNA 插入的X 位点两侧区域的碱基序列(图3B),该序列位于基因At4G39400(AtBRI1)和At4G39403(AtPLS)之间,属于基因的调控区(图3C)。
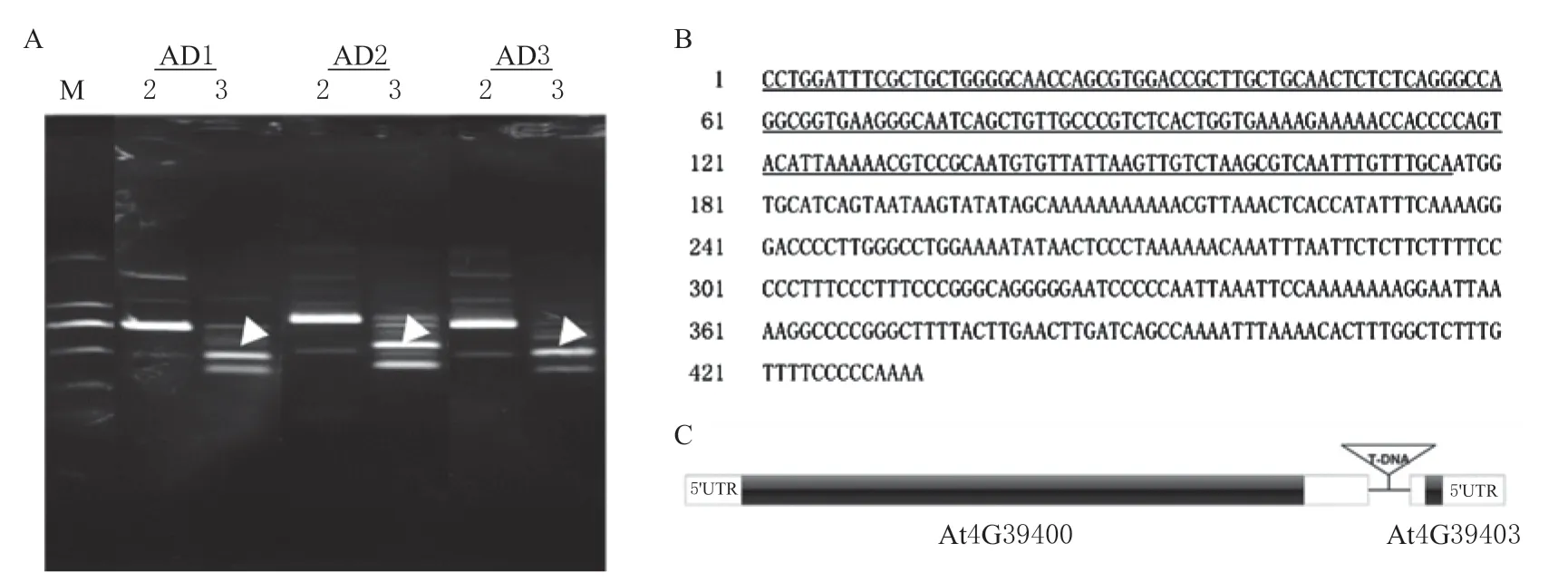
图3 SALK_037453X 中T-DNA 插入的序列Fig.3 Sequence of T-DNA insertion in SALK_037453X
获得X 位点的序列后,进一步通过遗传杂交结合PCR 检测的方法获取仅有X 位点T-DNA 插入的SALK_037453X 的纯合体,即SALK_037453X 中的T-DNA 只有1 个拷贝,且插入在X 位点,并观察表型。结果显示SALK_037453X 纯合体也呈现莲座叶聚集表型(图2A),证实该突变表型仅由T-DNA 在X位点的插入造成的。
2.3 T-DNA 在X 位点的插入突变影响其上下游基因的表达水平
进一步分析X 位点所在的序列。X 位点的T-DNA 插入在2 个基因的间区,对上下游基因的结构均未造成破坏(图3C)。而2 个基因之间的正常序列仅262 bp,既是At4G39400 的3’调控区,也是At4G39403 基因的3’调控区。利用PLACE和Plant CARE 对该序列进行了生物信息学分析,结果显示,该262 bp 的序列中,具有多种调控元件,包括多种胁迫响应元件以及转录增强子等调控序列(表5)。T-DNA 序列插入的位点在转录增强子CAAT box 的上游(图4A),由此,4.5 kb 长的T-DNA 不仅使得2 个基因之间的调控区域大大增长,且造成了上游基因At39400 远离了该CAAT box。进一步利用荧光定量PCR 检测了SALK_037453X中At4G39400(AtBRI1)和At4G39403(AtPLS)基因的转录水平的表达,发现与野生型拟南芥相比,SALK_037453X 中AtBRI1基因表达下调了22.9%,而AtPLS基因表达上调了8 倍(图4B)。
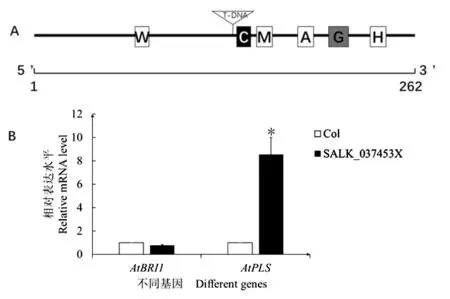
图4 X 位点的序列分析(A)及T-DNA 插入后对上下游基因表达的影响(B)Fig.4 Sequence analysis of X locus (A) and the effect of upstream and downstream gene expression after T-DNA insertion (B)

表5 SALK_037453X 顺式元件分析Table 5 Cis-element analysis of SALK_037453X
3 结论与讨论
叶片是植物的重要器官,植物叶片发育机制的阐明是以对所有参与叶片发育的基因功能的了解为基础的。尽管利用正向遗传学已经产生了大量的拟南芥叶片发育受损突变[9],但反向遗传学成为继拟南芥基因组测序之后的首选方法[10]。研究人员进行了大规模的表型研究,通过正向和反向的遗传筛选,鉴定出数百个参与叶片发育的拟南芥突变体,提高了对叶发育过程的认识[11-13]。但是,目前鉴定的叶发育相关基因在基因组中尚未饱和,仍需要发掘更多的参与叶片发育的基因,以及对已知基因的功能做更深入的分析。为此Wilson-Sánchez 等将已知数据存档在一个名为PhenoLeaf 的在线数据库(http://genetics.umh.es/PhenoLeaf)中[14]。
拟南芥叶的发育表型分为莲座叶表型、叶片表型、叶缘和叶柄表型等几个方面;其中莲座叶有大小、聚集和疏松等不同表型[14]。本研究从Salk 网站获得的突变体SALK_037453 的莲座叶即为聚集型表型。网站信息及RT-PCR 结果均显示,SALK_037453 突变体是AtPLC3基因表达缺陷突变体,尽管AtPLCs家族的基因参与了拟南芥生长发育和胁迫应答等多方面的生理过程[15],但是本研究的遗传分析结果显示,SALK_037453 的莲座叶聚集的表型不是由于AtPLC3基因表达缺失造成,而是由该突变体中还有一段T-DNA 插入到了At4G39400(AtBRI1)和At4G39403(AtPLS)2 个基因之间引起的。
At4G39400编码的基因AtBRI1(其他还有文献命名为BIN1,CABBAGE2,CBB2,DWARF2,DWF2等)编码质膜定位的富含亮氨酸的受体激酶,其配体是油菜素内酯。AtBRI1与配体结合后激酶结构域被磷酸化从而激活。油菜素内酯是重要的植物激素之一,而油菜素内酯信号途径在调控植物多种发育和应答反应中发挥着重要的作用[16]。作为该信号途径的受体,AtBRI1被广泛关注,迄今有该基因不同部位被破坏的多种突变体被用于对该受体作用机制的研究[17-19],而其表达缺失的突变表型之一就是形成聚集型的莲座叶[20],本试验获得的SALK_037453X 莲座叶的突变表型,即与该突变体中AtBRI1基因的表达下调相关。而且其表达下调是由于该基因的3’调控区引起的,这为目前对该基因功能的阐释提供了新的材料。
At4G39403编码1 个36 个氨基酸的短肽,被命名为Arabidopsis thaliana POLARIS,简写为AtPLS。AtPLS是根系和叶脉的正常发育所必需的,并且参与了乙烯、生长素等激素对植物的调节反应,但其作用的机制尚不清楚[20-21]。无论是乙烯还是生长素,对植物生长发育的调控都是非常重要的,而本研究获得的SALK_037453X 突变体,是AtPLS转录水平上升8 倍之多的超表达体,据目前对AtPLS功能的了解,该变化引起叶发育的变化也是有理论依据的。因此,SALK_037453X 突变体材料对于深入研究AtPLS的功能具有重要的价值。
基因组计划完成后,人们发现基因组序列中除了编码蛋白的基因外,还有大量的非编码蛋白的基因及更多的未知功能的序列。因此之后启动了“DNA 元件百科全书”计划(Encyclopedia of DNA Elements,ENCODE)。ENCODE 计划的研究对象除了编码蛋白基因外,还有非编码蛋白基因和调控区域等DNA 元件。多年来,该计划确定了许多之前不为人知的DNA 转录启动位点,并提出调控区域也有可能位于DNA 转录启动位点的下游等新观点。本研究获得的SALK_037453X 突变体,T-DNA的插入改变了2 个基因之间的调控区域的结构,从而导致2 个基因的表达均发生变化,暗示了2 个基因之间的262 个碱基中,存在着重要的调控基序。因此该突变体也为调控序列的研究提供了新的材料。
