超低温保存对胡杨种子活力及生理指标的影响研究
2020-07-22刘艳萍朱中原刘涛阳
刘艳萍 朱中原 刘涛阳
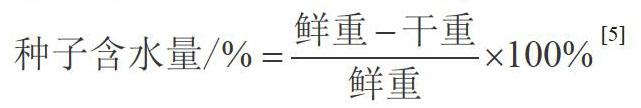
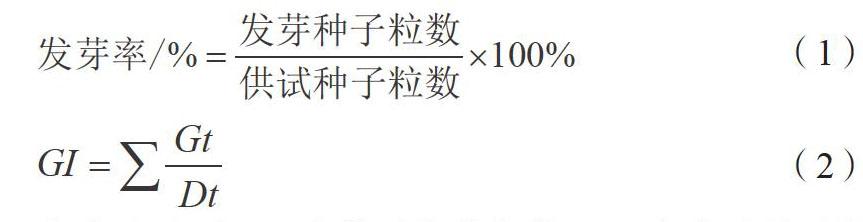
摘 要 胡杨种子在自然状态下极易失活,因此建立适合胡杨种子的保存方法十分必要。而超低温可以实现种子的长期保存。基于此,用硅胶对胡杨种子进行脱水处理,使种子的含水量形成不同梯度,再对不同含水量的种子进行超低温保存,然后对保存后的种子进行化冻处理,确定种子的存活率,测定种子的各种生理生化指标。实验结果表明,含水量是影响胡杨种子存活的重要因素,含水量为4.30%时胡杨种子的发芽率较高。影响胡杨种子的另外一个重要因素是化冻方法,采用快速化冻方法胡杨种子具有较高的存活率,且各种酶活性及种子的电导率、丙二醛含量都较低。对含水量为4.30%的胡杨种子进行超低温保存后,快速化冻对胡杨种子伤害较小,且对各种生理生化指标影响较小,适合胡杨种子超低温保存。
关键词 胡杨种子;超低温保存;种子活力;生理指标
中图分类号:S792.11 文献标志码:B DOI:10.19415/j.cnki.1673-890x.2020.14.071
胡杨属杨柳科落叶乔木,主要分布在中亚、西亚和地中海地区,在我国荒漠区有大面积天然分布,是生长于极端干旱荒漠区的唯一优势成林树种,但由于自然和人为因素的影响,胡杨面临濒危,并被列为我国三级重点保护植物。胡杨种子成熟后在自然状态下生命力在30 d
内就几乎完全丧失,且种子萌发成苗过程中会遭受气候、地下水位、土壤含水量和土壤盐渍化等外界环境因素的干扰。如何让胡杨种子在逆境胁迫下仍然保持活力,引起了学者的关注。张琴等[1]采用室温未密封、室温密封、4 ℃密封三种方式贮藏胡杨种子,测定种子劣变过程中的活力变化指标,结果表明,随着贮藏时间延长,种子含水量、发芽率、活力指数逐渐下降,活力下降速度为室温未密封﹥室温密封﹥4 ℃密封,三种贮藏方式中,以4 ℃密封贮藏方式效果最好,种子活力最高。张肖等[2]研究了胡杨种子萌发对温度、光照和盐旱胁迫的敏感性,结果表明,胡杨种子萌发对温度、光照不敏感,适宜溫度下萌发快速集中,且萌发时对盐旱胁迫具有一定的耐性。王海珍等[3]研究了胡杨和灰胡杨种子对不同盐类的耐受限度和适应萌发生境,结果表明两种胡杨均具有较强的耐盐性。伊丽米努尔等[4]研究表明,低浓度盐对胡杨种子萌发有一定的促进作用,而高盐浓度下,种子萌发受到明显抑制。刘艳萍等[5]采用硅胶脱水的方法制备不同含水量的胡杨种子,再经不同相对湿度溶液对种子进行回湿处理,确定了最适含水量及最适回湿方法。张玲等[6]采用包埋玻璃化液氮冷冻法对胡杨休眠茎尖进行冷冻保存,总结出包埋玻璃化法在液氮中保存胡杨的深冬休眠芽是可行的。目前,超低温保存是实现生物种质资源永久保存的理想方法,被广泛应用,已成功保存了近300种植物材料[7-8]。国内外超低温保存实际生产应用主要集中在农作物和蔬菜上,目前也已成功对部分种子、花粉、茎段、悬浮细胞等组织培养物进行了保存试验[9-11]。
大多数植物材料在超低温环境中细胞的新陈代谢活动会基本停止,从而使细胞活力能够得到稳定保存[12]。为了探寻胡杨种子长期低成本保存的新方法,本研究以胡杨种子为材料,对其进行超低温保存后种子的萌发特性、膜脂过氧化以及抗氧化保护系统研究。通过对不同含水量的胡杨种子进行保存,确定适合的胡杨种子超低温保存含水量,并对保存后的种子进行化冻处理,测定超低温保存后的各种生理生化指标,筛选出适合胡杨种子超低温保存的化冻方法,探究适合胡杨种子超低温保存的方法,为胡杨资源的永续利用提供理论依据。
1 材料与方法
1.1 实验材料的采集
于2018年8月在新疆阿拉尔市塔里木大学周边采集胡杨种子,自然晾干后,用纱网手工揉捏、过筛,剔除杂质,得到所需的种子,保存到4 ℃冰箱里备用。
经测定,所得胡杨种子初始含水量6.06%,发芽率为46.7%。
1.2 硅胶干燥法对胡杨种子的干燥处理
称取15组胡杨种子,每组7 g,采用硅胶干燥法干燥种子。将称取好的种子和变色硅胶置于同一干燥器中对胡杨种子进行干燥,变色硅胶和种子质量比为5∶1,干燥时间分别为1 d、2 d、3 d、4 d、5 d。干燥后按时取出种子,将不同含水量的种子密封放置于4℃冰箱保存备用。
1.3 不同化冻方法对胡杨种子的处理
将不同含水量的种子置于液氮中冷冻24 h后,采用两种不同化冻办法对种子进行化冻处理,分为两组:第一组取出后置于25 ℃温水中快速化冻15 min后置于4 ℃冰箱保存备用;第二组先置于-10 ℃冰箱化冻10 min后置于4 ℃冰箱继续化冻后保存备用。
1.4 种子含水量测定
采用分析天平随机称取1 g鲜种子于玻璃皿中,转入恒温烘箱并调节温度至121 ℃烘干17 h,冷却至室温后用分析天平称取干重,计算种子含水量(重复三次),测出种子初含水量为6.06%。计算公式如下。
1.5 发芽率和发芽指数测定
在培养皿内垫纸4层,用蒸馏水润湿芽床均匀放置30粒种子,移入人工气候培养箱,调节温度至24 ℃左右,设置光照15 h;温度15 ℃左右,黑暗9 h。分别记录第2天、第3天、第4天的种子发芽数。计算种子发芽率以及发芽指数,发芽率计算公式见公式(1),发芽指数计算公式见公式(2)。
公式(2)中GI为种子发芽指数,Gt为当天种子发芽数,Dt为种子发芽天数。
1.6 相关生理指标的测定
电导率的测定:随机称取1 g胡杨种子,放于烧杯中,倒入50 mL去离子水,用玻璃棒搅拌充分后,在25 ℃条件下静置1 d,次日再次用玻璃棒搅拌均匀后用电导仪测其浸出液,测出单位质量电导率。
超氧化物歧化酶(SOD)活性测定采用邻苯三酚自养化法,具体方法参照何乐韵等[13]的研究方法,本次稍有改动。
过氧化物酶(POD)活性测定采用愈创木酚法,过氧化氢酶(CAT)活性测定采用高锰酸钾滴定法,丙二醛含量(MDA)测定采用硫代巴比妥酸法。可参照刘艳萍等[5]的测定方法,本次稍有改动。
各项指标均设3个重复,每个重复测定3次,求平均值和标准差。
1.7 数据统计分析
采用统计软件DPS和Excel处理数据。
2 结果与分析
2.1 不同处理对胡杨种子活力的影响
2.1.1 超低温保存对种子活力的影响
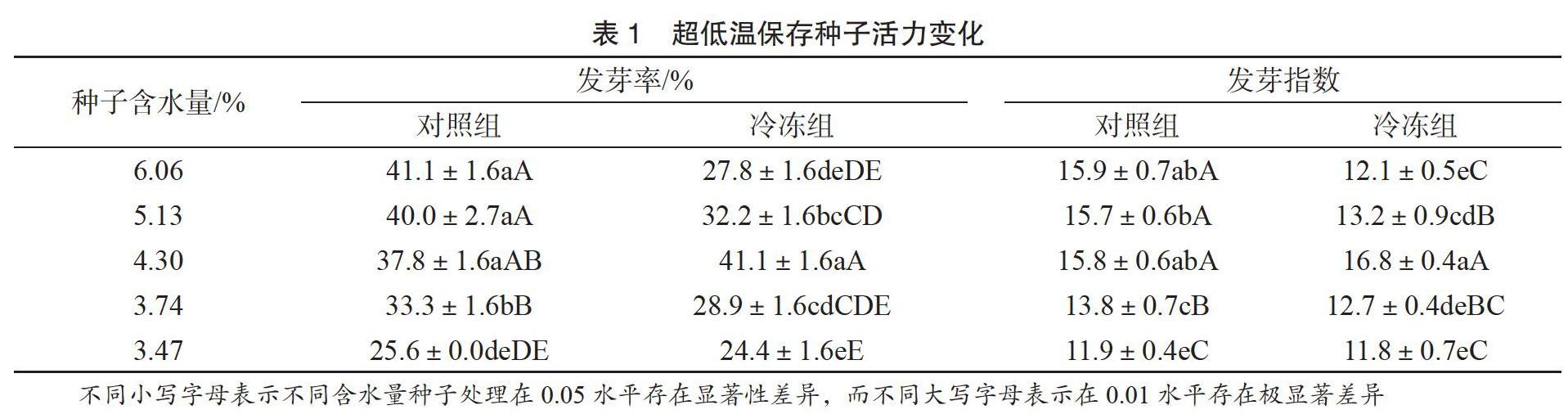
由表1可知,未冷冻处理的种子发芽率受种子含水量影响,随着含水量的降低,种子发芽率逐渐降低。经液氮超低温处理后并采用快速化冻方法化冻处理,随着种子含水量的降低,种子发芽率先升高后降低,当种子含水量在4.30%时,冷冻组种子的发芽率和发芽指数与对照组差异不显著,但均高于对照组。因此胡杨种子超低温保存的临界含水量为4.30%时对种子萌发有一定的促进作用。
2.1.2 不同化冻方法对种子活力的影响
由表2可知,对比采用两种不同化冻方法对胡杨种子进行化冻处理,其中采用快速化冻方法对胡杨种子进行化冻,种子整体活力水平均高于采用慢速化冻胡杨种子的活力,说明采用快速化冻比慢速化冻对种子造成的伤害小。两种不同化冻方法处理的胡杨种子,随着含水量的降低,胡杨种子发芽率和发芽指数均呈现出先升高再降低的趋势,并且在种子含水量为4.30%时种子活力最高。
2.2 不同处理对胡杨种子膜脂过氧化的影响
2.2.1丙二醛(MDA)含量
丙二醛(MDA)是膜脂过氧化作用最终分解产物,会抑制细胞保护酶的活性,降低抗氧化物含量,可以用来反映膜脂的过氧化程度[14-16]。图1显示种子经过液氮超低温处理后,不论采用何种化冻方式,含水量为4.30%时,种子中丙二醛含量均较低,其他含水量种子MDA含量均有明显升高,说明在超低温保存过程中对种子造成了一定伤害,选择合适含水量对胡杨种子进行超低温保存是有必要的[5]。
2.2.2 胡杨种子电导率的变化
由图2可知,經过超低温处理,且采用快速化冻方法化冻的胡杨种子整体水平电导率明显低于慢速化冻,并且当胡杨种子含水量在3.66%~5.02%,采用快速化冻种子电导率与对照组电导率相近,且当种子含水量在4.30%时,其电导率达到最低。因此,采用快速化冻方法对种子进行化冻,且含水量应控制在4.30%时,对种子的伤害程度较小。
2.3 不同处理对胡杨种子抗氧化保护系统的影响
2.3.1 过氧化物酶(POD)活性
如图3所示,随着种子含水量的降低,采用慢速化冻的种子POD活性上升幅度比采用慢速化冻明显且在含水量4.30%时达到峰值,接近未处理的种子活性,当含水量低于4.30%时采用慢速化冻的种子POD活性下降程度很大。相对而言,采用快速化冻方法对种子进行化冻,对种子POD活性影响不大。
2.3.2 超氧化物歧化酶(SOD)活性
超氧化物歧化酶(SOD)可以催化超氧化物歧化反应[17],是活性氧清除体系中第一个发挥作用的抗氧化酶。植物在低温寒冷等逆境胁迫下会产生对细胞有害的活性氧和自由基,SOD通过催化歧化反应使活性氧生成过氧化氢和氢气,减轻细胞活性氧伤害。由图4可知,经超低温处理后无论采用何种化冻方式,不同含水量的胡杨种子超氧化物歧化酶活性均有所下降,说明超低温冷冻处理对种子具有一定的伤害。相对而言,采用快速化冻方法对种子进行化冻比慢速化冻伤害小。不同含水量种子的超氧化物歧化酶活性也不同,在含水量4.30%时其活性最高。
2.3.3 过氧化氢酶(CAT)活性
过氧化氢酶(CAT)是植物体内与抗性相关的酶,不仅可以保护生物膜,还可以促进种子代谢,增加种子的抗逆性。采用快速化冻的种子和慢速化冻种子CAT活性均随含水量的降低呈先上升后下降的趋势,但采用快速化冻的种子CAT活性始终高于慢速化冻(图5)。因此,采用快速化冻的方法对种子进行化冻优于慢速化冻。
3 结论与讨论
超低温冷冻过程中,种子的含水量是影响其生命力的一个关键因素。理论上讲,含水量过高会使种子在冷冻和解冻过程中受到伤害甚至死亡,过低又会造成脱水伤害影响其活力[17]。种子发芽率和发芽指数是反映种子活力的综合指标,未冷冻处理的种子发芽率受种子含水量影响,随着含水量的降低种子发芽率逐渐降低。经超低温处理后并采用快速化冻方法化冻,随含水量降低,种子发芽率先升高后降低,在快速化冻种子含水量达到4.30%时,其发芽率和发芽指数均高于对照组,且采用快速化冻方法对胡杨种子进行化冻其活力整体水平均高于采用慢速化冻后胡杨种子的活力。说明超低温保存对胡杨种子萌发有促进作用,且胡杨种子超低温保存的临界含水量为4.30%。同时,快速化冻方法对胡杨种子进行化冻减少了对种子的伤害,可使种子保持较好的活力。
植物在逆境或衰老过程中,细胞内自由基代谢的平衡被破坏,有利于自由基产生,自由基过剩会引发或加剧膜脂过氧化,造成细胞膜系统损伤,导致细胞膜透性增大,同时使膜脂过氧化的产物MDA含量增加,而MDA是膜脂过氧化作用的最终产物,是膜系统受伤害的重要标志[18]。当细胞原生质膜受到破坏时,膜透性就会增加,细胞内部物质外渗,导致相对电导率增加,相对电导率反映了外渗程度和膜受损伤的程度[19]。本研究中,对胡杨种子进行超低温处理,采用快速化冻方法化冻胡杨种子,当种子含水量在4.30%时,对胡杨种子的伤害最小,说明在超低温保存过程中对种子造成了一定的伤害,当含水量降至4.30%时,MDA含量与对照组无差异,说明适宜的含水量可以缓解低温处理对细胞膜的伤害。
过氧化物酶(POD)是植物种子重要的保护酶,为植物细胞提供高效专一的活性氧清除体系,其活性与植物种子的超低温耐性相关[20]。植物在低温寒冷等逆境胁迫下会产生对细胞有害的活性氧和自由基,超氧化物歧化酶(SOD)通过催化歧化反应使活性氧生成过氧化氢和氢气,减轻细胞活性氧伤害。过氧化氢酶(CAT)是植物体内与抗性相关的酶类,不仅可以保护生物膜,还可以促进种子代谢,从而增加种子的抗逆性。
比较慢速化冻和快速化冻两种方法化冻后胡杨种子的抗氧化保护系统发现,快速化冻的种子POD、SOD、CAT活性指标均高于慢速化冻的种子的相应指标,说明快速化冻的种子这几种酶组成了完整的防氧链条,能够降低细胞膜脂过氧化,保持细胞膜的完整性,减缓低温对植物种子的伤害,促进种子在低温胁迫后萌发。环境胁迫破坏了植物体自由基的产生和清除之间的平衡,会导致自由基在植物体内积累,造成膜脂过氧化作用,对植物造成损伤[21- 22],因此采用慢速化冻的方法,对胡杨种子细胞膜造成损伤较重,这与宋红[23]超低温保存玉蝉花(Iris ensata)种子生理生化研究结果一致。
综上,胡杨种子不仅要在合适含水量范围内,还要采用合适的化冻方法对超低温冷冻处理后的种子进行化冻,才能有效减少超低温保存过程中对种子造成的伤害。超低温保存后,快速化冻对胡杨种子伤害较小,适度降低胡杨种子含水量可以提高超低温保存后种子的发芽率,最适含水量为4.30%。
参考文献:
[1] 张琴,马春晖,吴运来.三种不同贮藏方式对胡杨种子活力变化的影响[J].塔里木大学学报,2009,21(2):55-57.
[2] 张肖,王瑞清,李志军.胡杨种子萌发对温光条件和盐旱胁迫的响应特征[J].西北植物学报,2015,35(8):
1642-1649.
[3] 王海珍,韩路,贾文锁.胡杨(Populus euphraticu)与灰胡杨(Populus pruinosa)种子萌发对不同盐类胁迫的响应[J].中国沙漠,2013,33(3):743-750.
[4] 伊丽米努尔,木巴热克·阿尤普,雪合拉提·伊明诺夫,等.不同种源胡杨(Populus euphratica)种子萌发对盐胁迫的响应以及与生态因子的关系[J].西北林学院学报,2017,32(4):13-21,68.
[5] 刘艳萍,李利民,周晓明,等.超干处理对胡杨种子抗逆生理生化指标的影响[J].西北植物学报,2017,37(7):
1357-1363.
[6] 张玲,田洪涛,刘锦涛,等.胡杨休眠茎尖的包埋玻璃化法冷冻保存研究[J].新疆农业科学,2017,54(12):
2232-2238.
[7] 林富荣,顾万春.植物种质资源设施保存研究进展[J].2004,17(4):19-23.
[8] 王晗.人参种质资源超低温保存方法及其对遗传变异特性研究[D].北京:中国农业科学院,2016.
(下转第页)
(上接第页)
[9] 刘燕,周慧,方标.园林花卉种子超低温保存研究[J].北京林业大学学报,2001,23(4):39-44.
[10] 王越,刘燕.观赏植物种质资源的超低温保存[J].植物生理学通讯,2006,42(3):39-44.
[11] 张亚利,尚晓倩,刘燕.花粉超低温保存研究进展[J].北京林业大学学报,2006,28(4):139-147.
[12] 史锋厚,喻方圆,沈永宝,等.超低温贮藏对油松种子的影响[J].南京林业大学学报(自然科学版),2005,29(6):119-122.
[13] 何乐韵,周后珍,谭周亮,等.活性污泥中超氧化物歧化酶(SOD)活性测定影响因素[J].环境科学与技术,2012,35(12):146-149.
[14] 孔令琪.不同老化处理对燕麦种子生理、蛋白质及抗氧化基因的影响[D].北京:中国农业大学,2015.
[15] 高建强,农杆菌介导反义磷脂酶Dγ基因转化玉米的研究[D].泰安:山东农业大学,2007.
[16] 余欣欣.人工老化条件下不同基因型小麦种子活力的多态性及生理变化的研究[D].杨凌:西北农林科技大学,2015.
[17] 曾琳,何明军,陈葵,等.降香黄檀种子和离体胚超低温保存研究[J].中國中药杂志,2014,39(12):2263-2266.
[18] Alegre L, Munne-bosch S.Leaf senescence contributes to plant survival under drought stress[J].Functional Plant Biology,2004(31):203-216.
[19] 马丽,郝文芳,刘得方,等.水杨酸对低温胁迫种子萌发及细胞膜稳定性的影响[J].西北农林科技大学学报(自然科学版),2010,38(4):183-188.
[20] 曾琳,吴怡,何明军,等.超低温冷冻对益智种子生理生化特性的影响[J].广西植物,2018,38(4):529-535.
[21] Lorna S,Monk Kurt V,Fagerstedt Robert M,
et al.Oxygen toxicity and superoxide dismutase as an antioxidant in physiological stress[J].Physiologia Plantarum,1989,76(3):456-459.
[22] Pei ZM,Murata Y,Benning G,et al.Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells[J].Nature,2000,406(6797):731-734.
[23] 宋红,曹柏,王玲.超低温保存对玉蝉花种子生理生化特性的影响[J].东北林业大学学报,2016,44(4):44-47.
(责任编辑:赵中正)
