热休克蛋白HSPA2 对精子发生及其功能成熟的作用研究进展*
2020-07-17曹全富综述张庆华审校
李 锐 曹全富 综述 张庆华 刘 威 审校
1. 陆军军医大学大坪医院生殖医学中心(重庆 400042)2. 陆军军医大学基础医学院免疫学教研室(重庆 400038)
热休克蛋白是一类非常保守的蛋白质,其主要功能是缓冲各种环境有害因素可能对细胞造成的不利影响。它们主要行使分子伴侣或伴侣辅助分子的功能,负责辅助完成细胞内蛋白质的合成、稳定、折叠、转运和降解,以及蛋白复合体的组装与维系。分子量70 kDa 的热休克蛋白(heat shock protein 70,HSP70)是数量最丰富、进化最保守的热休克蛋白家族之一,在人类基因组中至少含有13 个编码基因[1,2]。
精子发生是从精原细胞到精子的发育过程,此过程得以正常进行,部分归功于热休克蛋白。精子发生完成时,精子在结构上已发育成熟,但此时尚不具备完整的生物学功能,还需进行功能转化,其主要体现为精子结构重塑及其功能改变,这种功能转化不仅发生于男性生殖道中(主要是附睾),还发生于精子在女性生殖道内获能的过程中,最终产生功能成熟的精子,而此过程也与HSP 有关。本文重点讲述HSPA2,这是一种在睾丸中大量表达的HSP70 家族成员,以前的名称为HSP70.2。它的功能就是促进胞内蛋白质的折叠和运输,以及蛋白复合体的组装,其表达水平与体外受精能力呈正相关。本文从HSPA2 的分子特征和活性机制、HSPA2 在男性精子发生和功能成熟中的作用、 以及HSPA2 在氧化压力下发生4- 羟基壬烯醛(4-hydroxynonenal,4HNE) 修饰等几个方面进行综述,以期为HSPA2 的进一步研究提供理论指导。
一、HSPA2 的分子特征和活性机制
HSP70 家族成员具有一个共同的、 高度保守的模块化结构,包括一个N 端的核苷酸结合结构域(nucleotide binding domain,NBD),也称ATP 酶结构域,一个位于中心的连接区域,以及位于C 端内的底物结合结构域(substrate-binding domain,SBD)。与所有HSP70一样,HSPA2 内的NBD 和SBD 通过一段保守的疏水序列连接(图1A)。在结构上,NBD 包括两个亚结构域,核苷酸即结合于两者之间的缝隙中。该结构域与ATP有很强的亲和力,一旦结合,就可以通过水解ATP 所提供的能量来驱动其构象改变(图1B)。SBD 由一个β 夹心子域SBD-(β)和一个α 螺旋盖区SBD-(α)组成。(图1A)。SBD 是结合底物的关键区域,其能够与5-7 个疏水氨基酸所组成的肽段相结合。
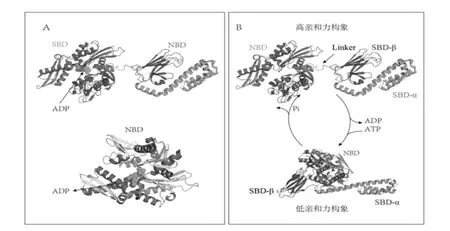
图1 HSPA2 的结构和反应周期
HSPA2 依赖NBD 的ATP 酶活性来调节底物的结合和释放。当ATP 与NBD 结合时,HSP70 的构象呈现“开放状态”,表现出低底物亲和力(图1B 下),此构象可稳定新生或变性蛋白,防止这些未折叠多肽的聚集。当ATP水解成ADP 时,会形成“封闭状态”的高亲和力构象(图1B 上),此构象可引发蛋白质折叠(与伴侣辅助分子一起),多聚体复合物的形成,以及目标蛋白跨膜转运。整个反应周期在HSP70 家族的大多数成员中以类似方式进行。
二、HSPA2 在男性精子发生中的作用
基于小鼠的研究发现,HSPA2 最早出现于生精细胞第一次减数分裂早期(细线期-偶线期)。被选择性敲除hspa2 基因的雄性小鼠其生精细胞在第一次减数分裂的G2-M 阶段受阻,在粗线期经凋亡被清除,最终引发不育。目前认为HSPA2 在小鼠生精细胞中有两个主要功能,第一HSPA2 似乎促进细胞周期蛋白依赖性激酶2 与细胞周期调控蛋白B1 之间的相互作用,第二其作为联会复合体的“调节器”在同源染色体之间介导染色体配对、联会和减数分裂重组事件。除了支持减数分裂外,HSPA2 还在减数分裂后辅助过渡蛋白完成特定的功能[3]。一项基于大鼠的研究表明,在精子顶体形成过程中胞质中的HSPA2 被募集到高尔基体中[4],但其功能尚不明确,有待进一步研究。
HSPA2 在人类精子发生过程中似乎以双相模式进行表达。该蛋白第一次表达始于精母细胞,推测其支持减数分裂。尽管没有足够证据支持人类HSPA2 如小鼠研究中所报道的即在调节联会复合体或辅助细胞周期蛋白- 周期蛋白依赖性激酶复合体的过程中发挥类似作用,但大量数据显示HSPA2 在睾丸中的表达水平与射精精子浓度之间存在着很强的相关性。据报道,在患有少精子症、 精母细胞发育停滞或唯支持细胞综合征所导致的完全非梗阻性无精子症 (即射精时没有可检测的精子)的不育患者中,HSPA2 表达水平显著降低[5]。此外,人类和小鼠HSPA2 同源物的氨基酸序列保守程度超过98%,有研究指出HSPA2 在精子发生方面很可能具有保守的功能。其第二次表达在长形精子细胞形成过程中开始[6],既往研究表明HSPA2 在此阶段参与细胞质挤压和质膜重构[7,8],HSPA2 表达水平下降会导致精子细胞质保留过多[7]。
总之,HSPA2 可以依赖其分子伴侣活性在精子发生的不同阶段发挥特定功能,是精子发生得以正常进行的必要因素。
三、HSPA2 在男性精子成熟中的作用
大量研究表明HSPA2 对精子的功能转化 (成熟)至关重要,一个直接证据是,HSPA2 被发现定位于人类成熟精子顶体表面,这一位置与精卵相互识别、相互作用的位置一致[9]。另有研究指出,弱精子症患者精子中HSPA2 表达水平升高[10],精浆中游离的HSPA2 也与正常人群存在显著差异,并且与精子活力呈负相关[11]。目前认为HSPA2 可促进并维系精子表面卵透明带(zona pellucida,ZP)识别复合物(也称ZP 受体)的组装、重定位和暴露[9,12,13]。
在正常精子中HSPA2 与一些蛋白多聚体存在稳定的相互作用[9,14-16],目前已发现人类精子蛋白质组中有几个潜在的HSPA2 结合底物[9,14,15],其中研究最深入的是精子粘附分子1 (sperm adhesion molecule 1,SPAM1)和芳香基硫酸酯酶A(arylsulfatase A,ARSA)。HSPA2 与SPAM1 和ARSA组成的复合物似乎经历了一个与精子获能相关的结构重塑过程。ARSA 发生重定位而易于被精子细胞膜表面抗体所标记,与之相反SPAM1 重定位后在精子细胞膜表面含量下降。ARSA与SPAM1 的这种结构重塑还与精子在获能晚期失去结合透明质酸的能力相一致[17]。值得注意的是,重定位后ARSA 所在位置正好与卵透明带相互作用相适应,而SPAM1 主要在精卵识别之前参与精子穿越卵丘基质的过程[9]。HSPA2 也能与血管紧张素转换酶(angiotensin converting enzyme,ACE) 和蛋白二硫键异构酶A6 (protein disulphide isomerase A6 ,PDIA6)形成稳定复合物[15]。只有当精子处于获能状态时,PDIA6 才能在精子表面被标记,而ACE 却无法被标记。另一方面,通过药物抑制ACE 后,精子诱发顶体反应的能力显著降低,说明HSPA2 所介导的PDIA6-ACE 复合物对于受精能力极其重要[15]。最近研究发现,HSPA2 与伴侣辅助蛋白(BCL2-associated athanogene 6,BAG6)及解整合素样金属蛋白酶 (A Disintegrin And Metalloproteinase Domain 30,ADAM30)也可形成稳定的复合物,该复合物位于精子头部顶端区域。推测BAG6 对于由HSPA2所介导的精子ZP 受体复合物的装配有所帮助,可防止发生蛋白质错误折叠或错误定位。在因精卵结合障碍而导致不育的患者中,不仅HSPA2 表达水平下降,BAG6 的蛋白表达也严重降低[14]。
尽管当前精子表面发生结构重塑的分子机制还知之甚少,但已发现HSPA2 复合体似乎与膜筏的结构标记物共定位,而膜筏是精子头部获能过程中重定位的微区(microdomain)[12]。研究显示,由精子获能驱动的HSPA2 翻译后修饰对于这种结构重塑至关重要,成熟精子经过广谱酪氨酸激酶抑制之后,与HSPA2 相关的重定位将不再发生[13]。另有研究报导,成熟精子暴露于氧化应激环境也会破坏HSPA2 所介导的精子表面结构重塑[18]。
此外,HSPA2 表达水平下降可导致不成熟精子产生[19],主要表现为精子在受精过程中结合ZP 的能力明显降低,最终导致受精失败[8,19]。最近一项比较蛋白质组学研究也证实,在精卵识别障碍的不育患者精子中,HSPA2 表达水平明显不足[9]。尽管此项研究未能将HSPA2 缺陷与精子活力或形态方面的缺陷联系起来,但这一证据显示HSPA2 对精子发生完成后的功能成熟极其重要[20]。
总之,这些实验结果表明HSPA2 很可能通过与多种ZP 受体关键蛋白分子形成复合物,稳定它们在精子表面的取向与定位,从而在精子获能、精卵识别、及精子穿越卵丘基质与透明带等过程中发挥关键作用。
四、 HSPA2 功能障碍与4- 羟基壬烯醛(4HNE)修饰
一个值得关注的问题是,部分男性不育患者精子中HSPA2 的功能障碍是如何发生的? 尽管与此相关的研究有很多,但目前主流的研究模型集中在氧化应激的影响上。氧化应激可通过诱导脂质过氧化级联反应,形成一系列小分子量的醛类化合物,4- 羟基壬烯醛(4-hydroxynonenal,4HNE)是其中常见的一种。研究表明,4HNE 所造成的HSPA2 化学修饰可能损害其分子伴侣活性,并进一步造成精子质膜重构异常[18]。此外,这种修饰不仅会影响HSPA2/SPAM1/ARSA 受体复合物的组装与维系,也会影响上述HSPA2/PDIA6/ACE 复合物[15,18]。另有研究表明,4HNE 加合会优先靶向半胱氨酸、赖氨酸和组氨酸残基,从而引发一系列结构和功能变化,包括底物识别能力的丧失或重折叠能力的丧失[21-25]。已有明确生化实验证实,4HNE 可以共价修饰HSPA2 分子NBD 内的Cys267 残基[26],从而减弱HSPA2的ATP 酶活性,并阻止其在蛋白质运输和重折叠事件中发挥作用[26,27]。共价修饰的HSPA2 很可能通过蛋白酶体途径降解[28],此外,4HNE 还可能通过增强细胞蛋白酶体活性而加速细胞内蛋白质的水解[29]。但有争议的是,较高浓度的4HNE 可以极大促使蛋白质交联,从而导致蛋白酶体失活[30,31]。另有报道称,4HNE 修饰可经溶酶体途径介导蛋白降解[32]。总之,基于这些实验数据可以推测患病群体精子发育过程中HSPA2 水平下降很可能与4HNE 修饰所致的蛋白质降解有关。此外,在BAG6 缺失的转基因小鼠睾丸中,HSPA2 的保护性伴侣分子BAG6 被敲除后,HSPA2 蛋白水解速度也随之加快[33],这可以作为上述推测的一个佐证。
五、总结
HSPA2 表达水平、 结构完整性与功能状态直接关系到精子发生、成熟、获能及精卵识别等过程,从而决定最终能否成功受精。从此意义来说,HSPA2 很有可能成为临床上评判男性生育力的生物标志物。然而,目前关于HSPA2 的表达调控、作用机制与环境压力下活性丧失的研究还很不充分。综合已有研究成果来看,HSPA2 可在两个阶段发挥重要作用,一是精子发生过程中维系减数分裂复合物,二是精子功能转化(成熟)过程中促进精子顶体表面ZP 受体复合物的组装、定位与稳定,从而保证精卵的正确识别。与此相应,HSPA2的表达也经历了两次高潮。另一方面,环境因素如氧化压力既可通过影响DNA 甲基化而造成HSPA2 的转录抑制,从而造成HSPA2 表达水平下降,也可通过共价修饰造成HSPA2 三维结构改变,进而导致其活性丧失。但目前为止还有很多问题尚不明确,解决这些问题将对不育症的诊断、治疗和预防具有重要意义。
