广西夏季不同生境福寿螺繁殖力研究
2020-07-14梁碧霞陈欣宇武正军程瑞黄锦龙
梁碧霞 陈欣宇 武正军 程瑞 黄锦龙


摘要:为了解广西不同地区、不同生境福寿螺(Pomacea can aliculata)繁殖力大小及其影响因素,于2018年7-8月采集广西桂林市、来宾市、北海市3个地区6个生境(稻田、荒田、水沟、池塘、河流、沼泽)的福寿螺卵块,测量卵块长、宽、重量、卵粒总数和卵粒直径,并进行相关性分析。结果表明,广西夏季福寿螺单块卵粒数为48-1 041粒,均值为(256.19+155.67)粒,其中池塘生境卵粒数最多,均值为(395.61±213.33)粒,荒田生境卵粒数最少,均值为(168.39+55.47)粒;卵粒直径为1.72-2.72 mm,均值为(2.17±0.17) mm,其中水沟生境卵粒直径最大,均值为(2.32+0.13) mm,稻田生境卵粒直径最小,均值为(2.07±0.16) mm;卵粒数和卵粒直径在不同地区均表现为来宾市>桂林市>北海市,卵粒数量在生境间表现为池塘>河流>沼泽>水沟>稻田>荒田,卵粒直径大小在生境间表现为池塘>沼泽>水沟>河流>荒田>稻田。夏季廣西福寿螺繁殖力与纬度无关,离水面高度适宜繁殖或食物丰富的生境下福寿螺卵粒数较多、卵粒直径较大。在不同地区、生境间福寿螺卵粒数和卵粒直径有显著差异。
关键词:福寿螺(Pomacea canaliculata);繁殖力;广西
中图分类号:Q958.2
文献标识码:A
文章编号:0439-8114( 2020) 08-0083-06
D01:10.1408 8/j .cnki.is sn043 9- 8114.2020.08.019
福寿螺(Pomacea canalicutata)又名大瓶螺、苹果螺,属软体动物门( Mollusca)腹足纲(Gastropoda)瓶螺科(A itlpullaridae)[1,2]]。原产于南美洲亚马逊河流域,属于热带和亚热带种[3,4]。福寿螺自20世纪80年代被当作养殖品种引入中国台湾和广东省后,由于其食味不佳,市场供大于求,大量福寿螺被养殖户遗弃至野外[5],而后随水流、灌溉流人农田、沟渠、江河、池塘等。因福寿螺食性广、耐受性强、生长繁殖快等因素,在野外迅速建立了种群,在广西、福建、海南、浙江、上海、江苏、江西、湖南、四川、贵州等省均报道有福寿螺的分布[6,7]。福寿螺在2000年被IUCN外来人侵物种专家委员会列为世界100种恶性外来人侵物种之一[8]。2003年也被国家环保总局列入了首批入侵中国的16种外来物种的“黑名单”[9]。福寿螺不仅为害农田经济作物,其作为广州管圆线虫( Angiostrongylus cantonensis)的中间宿主,还对人类造成严重威胁,北京等地已经有因人生食或半生食福寿螺而感染患病的相关报道[10,11]。近年来,政府不断加强防治福寿螺繁殖扩散的力度,一定程度上抑制了其增长进程,但效果不显著。江西、广东、广西等大部分地区已经成为福寿螺主要集中区域,其扩散速度达7.04 km/年,目前福寿螺在中国南部的扩散速度仍在持续增长[12]。
纬度是生态学研究中重要的空间尺度,是各种环境因素的综合体现,对生物体生存的各种生态因子均能产生一定的影响,甚至直接作用于生物体[13-16]。前人研究分析结果显示,年均温是影响福寿螺分布的主要环境变量[13]。而纬度是影响地区年均温的重要因素,随着纬度的增加年均温会随之下降,福寿螺的生长、繁殖均受温度条件的影响。尽管张灿宇等[17]研究了中国18。N-28。N3个生境的福寿螺变异情况,但国内许多学者还是就部分地区福寿螺繁殖量或福寿螺繁殖月份、温度、历时、卵粒等进行了研究,主要集中在气温与福寿螺繁殖关系[18-21]、月份与繁殖力关系[22-24]、螺规格大小与卵块、卵粒差异[25.26]等方面。针对广西不同纬度的不同生境的福寿螺繁殖力差异研究鲜见报道。广西属于福寿螺的重点危害区,研究不同生境的福寿螺繁殖力,有助于了解不同纬度、不同生境福寿螺的繁殖特点,以及近年来广西各地不同生境的福寿螺防控效果,为进一步做好不同生境的福寿螺防治对策提供理论支持,对广西福寿螺防控具有重要意义。
1 研究区域及方法
1.1 研究区域概况
广西壮族自治区位于104° 26' E-112° 04' E、20°54' N-26°20'N,北回归线横贯全区中部,全境南北宽约610 km,东西长约750 km,陆地边境线1 020 km,大陆海岸线1 595 km,东与广东、海南为邻,南临北部湾,西南与越南交界,西北连云南、贵州,东北同湖南相接。全自治区总面积23.6万多平方千米,占全国土地总面积的2.46%[27]。本试验研究地点主要为广西桂林市、来宾市、北海市3个地区。桂林市位于广西东北部,地处109°36' E-111。29tE、24°15' N-26°23'N,位于越城岭南麓,属于亚热带季风气候,冬短夏长,雨热同期[28]。来宾市位于广西中部,地处108°24' E-110°28' E、23°16' N-24。29' N,属于中亚热带向南亚热带过渡的季风气候区,北回归线从市内南缘通过,气候温和,雨量充沛[29]。北海市位于广西南部、北部湾东海岸,地处108°50' E-109°47'E、20°26' N-21°55'N,属于海洋性季风气候,具有典型的亚热带特色,秋春相连,长夏无冬[30]。
1.2 材料
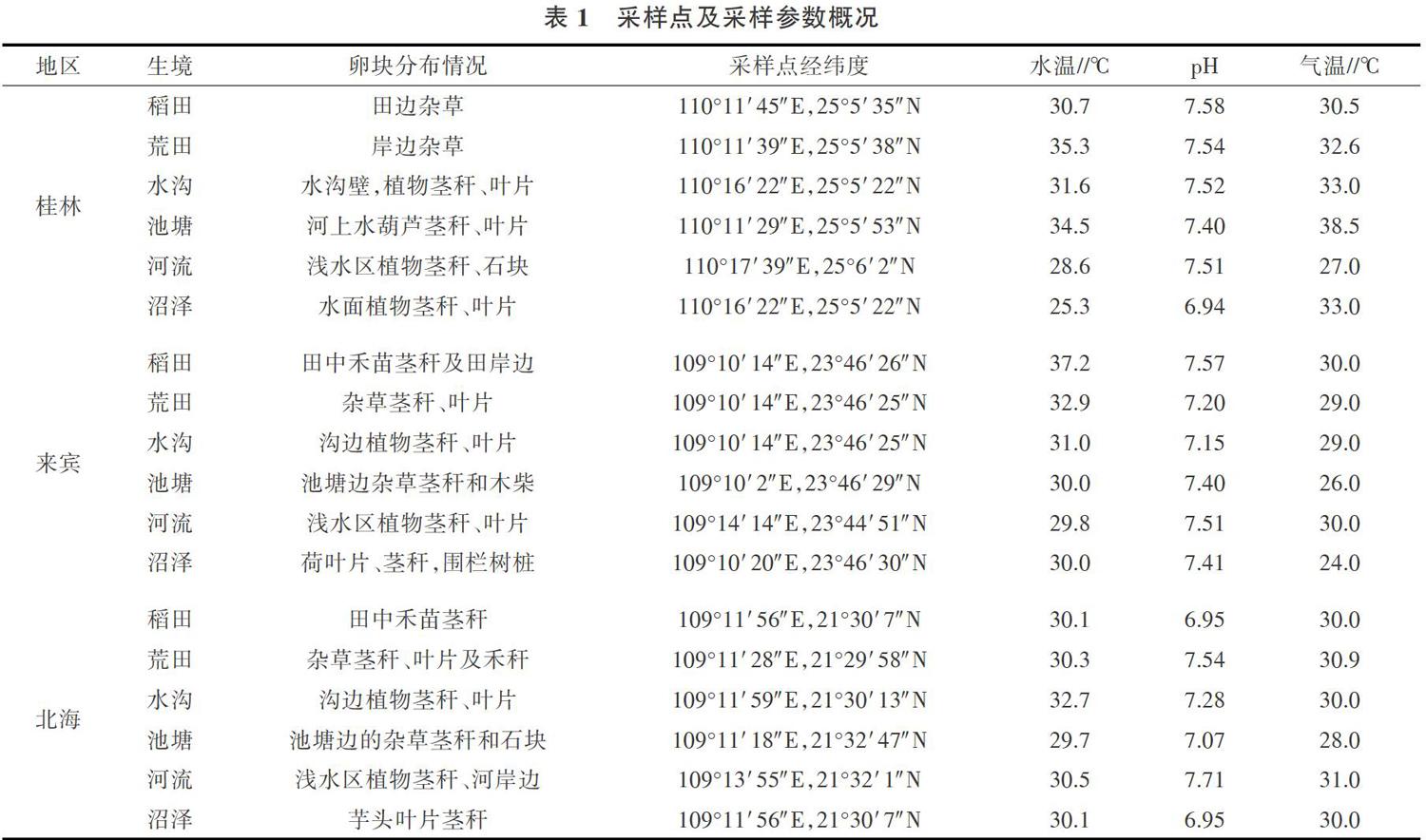
2018年7-8月于广西3个地区的6个生境(稻田、荒田、水沟、池塘、河流、沼泽)随机采集540枚完整的福寿螺卵块样品,每个地区、每个生境各采集30枚。并对采集的福寿螺卵块进行研究。取样点经纬度及采样参数情况如表1所示。
1.3 方法
将采集到的样品用电子天平(精确到0.01 g)测量福寿螺卵块重量,用量程为15 cm的数显游标卡尺(精确到0.01 mm)测量福寿螺卵块长(卵块最长距离)、宽(与卵块最长距离相垂直的最宽距离)。随机选择每枚福寿螺卵块中的20粒卵粒,用游标卡尺(精确到0.01 mm)进行直径测量(每粒卵粒测两次,取平均值),最后统计出每个福寿螺卵块的总卵粒数。
1.4 数据统计分析
采用Excel 2010、SPSS 13.0和R软件v 3.5.1对540组试验数据进行整理。利用描述统计法对福寿螺卵块样品各繁殖力参数情况进行说明,采用混合线性模型分析3个地区、6个生境福寿螺卵粒数和卵粒直径相关性及差异,整体绘图采用Excel 2010和Sigmaplot 12.5进行。
2 结果与分析
2.1 福寿螺卵块参数情况
3个地区6个生境采集的540枚试验样品的参数情况见表2。夏季广西3个地区的福寿螺卵块总卵数量平均为(256.19+155.67)粒,北海市所有生境的卵粒数均小于该值;卵粒直径平均为( 2.17+0.17)mm,除河流、池塘生境外,北海市所有生境的卵粒直径均小于该值。卵粒数量最多的是来宾市的池塘生境,为(574.47+249.14)粒,卵粒数量最少的是北海市的水沟生境,为(147.40+66.32)粒。所有样本中每枚卵块中卵粒数极大值来自来宾市池塘生境,为1041粒/枚,极小值来自桂林市河流生境和来宾市水沟生境,为48粒/枚,卵粒直径极大值来自桂林市沼泽生境,为2.73 mm,极小值来自北海市水沟生境,为1.73mm。
2.2 福寿螺卵粒数情况
广西3个地区福寿螺卵粒数的混合线性分析如表3所示,每个地区共计测量180枚卵块。北海市福寿螺繁殖力最小,卵粒数为(180.69+68.76)粒;来宾市繁殖力最大,卵粒数为( 320.25+194.88)粒;桂林市繁殖力居中,卵粒数为(267.62+142.38)粒。3个地区之间的福寿螺繁殖力存在显著性差异)X2=99.601,df=2,P<0.001),地区之间福寿螺繁殖力大小表现为来宾市>桂林市>北海市。
广西6个生境福寿螺卵粒数的混合线性分析如表4所示,每个生境测量90枚卵块。6个生境中荒田生境福寿螺繁殖力最小,卵粒数为(168.39+55.47)粒,池塘生境繁殖力最大,卵粒数为( 395.61+213.33)粒。6个生境之间的福寿螺繁殖力存在显著性差异(X2-137.83,df=5,P<0.001),生境之间福寿螺繁殖力大小表现为池塘>河流>沼泽>水沟>稻田>荒田。
2.3 福寿螺卵粒直径情况
广西3个地区福寿螺卵粒直径的混合线性分析如表5所示,每个地区测量180枚卵块。北海市福寿螺卵粒直径最小,为( 2.06+0.16) mm;来宾市卵粒直径最大,为( 2.25+0.12) mm;桂林市卵粒直径大小居中,为( 2.20+0. 16)mm。3个地区之间的福寿螺卵粒直径存在显著性差异(X2=166.61,df=2,P<0.001),卵粒直径大小表现为来宾市>桂林市>北海市。
广西6个生境福寿螺卵粒数的混合线性模型如表6所示,每个生境共计测量90枚卵块。6个生境中稻田生境福寿螺卵粒直径最小,为(2.07+0.16) mm,池塘生境卵粒直径最大,为( 2.24+0.13)mm。6个生境之间的福寿螺卵粒直径存在显著性差异(X2=92.925,df=5,P<0.001),生境之间福寿螺卵粒直径大小表现为池塘>沼泽>水沟>河流>荒田>稻田。
3 小结与讨论
3.1 福寿螺繁殖力情况
福寿螺繁殖能力极强,条件适宜时,单只雌螺1次能产卵50-1 000粒,在10-15 d孵化期内孵化率超过80%[31]。在本研究中,广西夏季3个地区(21°N-25°N)福寿螺卵块平均卵粒数为256粒/枚,卵粒数量较前人同季节研究中18°N-28°N福寿螺卵块平均卵粒数279粒/枚少;卵粒直径平均值為2.17 mm,较18°N-28°N福寿螺卵粒直径2.16 mm稍大[17]。广西不同地区之间福寿螺卵粒数与卵粒直径大小有一定关系,卵粒数多的地区其卵粒直径也相应较大,3个地区之间卵粒数及卵粒直径大小均表现为来宾市>桂林市>北海市。这说明来宾市、桂林市仍是福寿螺高危害区,特别是来宾市福寿螺繁殖量均值达到320粒,枚,应加强该地区的防控力度。
在人为的干预下,稻田、荒田等生境的福寿螺繁殖力控制得较好,但引水人农田的淡水生境中福寿螺繁殖力仍较高,6个生境之间福寿螺繁殖力大小表现为池塘>河流>沼泽>水沟>稻田>荒田。这说明人为防控范围缺失的淡水生境福寿螺严重泛滥,且前期经过翻田暴晒灭螺环节的农田,仍会因为引水灌溉带入螺或卵而再次暴发福寿螺危害。因此,在引水人田之前应对引入农田的淡水进行灭螺处理,并加强对农田周边淡水生境的福寿螺进行人为防控。
3.2 影响福寿螺繁殖力的因素
福寿螺的入侵和生长深受纬度因素的影响,纬度位置所影响的环境温度、生境条件、饵料来源等因素与福寿螺的繁殖能力有着密切的关系[32-34]。本研究中3个纬度的福寿螺繁殖力之间存在差异,但并不完全与纬度相关,还与不同地区、不同生境中能提供的营养物质有关。前人研究显示,福寿螺的生长发育和繁殖会受到外来因素的影响,环境的变化会干扰福寿螺的繁殖,会对福寿螺的产卵周期和产卵量有影响[35-37]。福寿螺在饥饿状态下产卵数量明显低于正常水平,福寿螺个体性状特征也会对卵粒数量有重要的影响[38.39J。不同生境营养物质的量不同,这可能是影响不同生境卵粒数量差异的原因,营养化越好的生境,福寿螺卵块平均卵粒数越多,卵粒直径也越大。
福寿螺卵粒数量还与其产卵特性相关[40]。福寿螺具有沿壁产卵的特性,主要聚集在田埂边、水沟壁及池塘、河流边露出水面的植物茎秆处,在这些区域发现鲜红色福寿螺卵块,其在生境的分布呈边缘向中心减少的趋势[41j。前人研究结果表明,福寿螺产卵高度与水面距岸壁的高度有关联[42],在自然条件下,由于福寿螺卵在水中不能正常发育,为了更好地繁衍后代,福寿螺总是将卵产在距水面一定高度的岸壁或者其他物体上[43],在露出水面10-80 cm的物体上均能发现福寿螺的卵块,其中露出水面30-50cm高的物体上的卵块数量最多[44]。池塘生境浅水区的植物茎秆露出水面10-30 cm,来宾市池塘生境富营养化严重,且池塘边的木柴长达100 cm,符合福寿螺最喜欢产卵的水面高度,所以露出水面的植物茎秆和木柴上附着较大的鲜红福寿螺卵块。而夏季的稻田、荒田处于返青期,水稻茎秆上的分蘖导致其表面凹凸不平,且成熟期的叶片表面有细小的毛刺,影响雌螺的爬行及卵块的附着,所以这两个生境的福寿螺卵粒数较少。这与前人研究的结论一致,即福寿螺产卵情况在不同水稻生长期有所不同[42]。
广西福寿螺夏季繁殖量与纬度关系甚微,主要是因为广西夏季日均温相差不大,都达到了福寿螺繁殖的适宜温度(25℃以上),故各地繁殖力的差异主要与该地区各生境的食物量、天敌量和人为的防控行为有关。不同的灭螺力度和灭螺方法会达到不同的灭螺效果。采用的主要灭螺方法有农业防治、生物防治、化学防治和物理防治[45.46]。经实际走访调查,发现北海市多处淡水生境受到人为喷洒灭螺剂影响,农田及周边水渠灭螺频率和灭螺力度都加大,市区周边的水域和生境都极少看到福寿螺卵块,在偏远的农田和其他生境的福寿螺卵块也因农药残留产卵量较少,大大缩小了福寿螺分布范围。该地区农民对福寿螺的防控意识也更为强烈,一旦发现大面积福寿螺卵块出现,还会人为进行挤压踩碎破坏,减少福寿螺发生率。说明对重点危害地区积极防治,能有效减轻福寿螺危害。
自福寿螺在中国成为危害严重的入侵生物以来,广大科研工作者也加强了控制福寿螺扩散和繁殖的研究,并取得一定成果[47]。本研究揭示,不仅在危害严重的地区,在数量较少的季节和地区也应加强对福寿螺的监测和防治工作,从薄弱处着手,严防危害:在农田以外的淡水生境也要进行全面防范,在引水人田前要对引入的淡水进行灭螺处理,以防引入福寿螺于农田暴发灾害;对池塘、水沟等与农田紧密相连且福寿螺繁殖力强的生境,应加强防控力度,防止福寿螺蔓延危害;对于福寿螺危害严重的重灾区,应当做好定期监测和预测工作,利用福寿螺季节性繁殖消减规律,及时控制灾情,把福寿螺的危害损失降到最低水平。
参考文献:
[1]丁建清,解焱.中国外来种入侵机制及对策[M].北京:中国环境科学出版社,1996.107-128.
[2]俞晓平,和田节,李中方,等.稻田福寿螺的发生和治理[J].浙江农业学报,2001,13(5):247-252.
[3]周卫川.外来人侵生物福寿螺的风险分析[J].检验检疫科学,2004.14(6):37-39.
[4]刘建文,王华生,谢茂昌.广西农田福寿螺发生现状及防治策略[J]广西植保,2006,19(4):21-23.
[5]杨叶欣,胡隐吕,李小慧,等.福寿螺在中国的入侵历史、扩散规律和危害的调查分析[J].中国农学通报,2010,26(5):245-250.
[6]江文彪,杨桂梅.浅谈防城港市农田福寿螺发生趋势及防治对策[J].广西农学报,2006,21(5):23-25.
[7]章家恩,方丽.关于我国农田福寿螺生物入侵需要加以研究的生态学问题[J].中国生态农业学报,2008,16(6):1585-1589.
[8] LOWE S M,BROWNE M. BOUDJELAS S, et al.100 0f the world'sworst invasive alien specics:A selection from the global invasivespecies datahase[ M]. Auckland, UK: The invasive species special-ist group.IUCN, 2007.206-208
[9]房苗,徐猛,罗 渡,等.不同生境下福寿螺的可量性状特征、种群结构和密度效应[J].生态学杂志,2016,35(3):747-754.
[10]赵本良,章家恩,戴晓燕,等.福寿螺对稻田水生植物群落实验的影响[J].生态学报,2014,34(4):907-915.
[11]黄达娜,黄亚兰,阳帆,等.输人性福寿螺传播广州管圆线虫的潜能研究[J].中国热带医学,2017,17(9):871-875.
[12]周宇,袁雪颖,杨子轩,等.福寿螺入侵中国的扩散动态及潜在分布[J].湖泊科学,2018,30(5):1379-1387.
[13] SHERMAN R E. FAHEY T J,MARTINEZ P. Spatial patterns ofbiomass and aboveground net primary productivity in amangroveecosvstem in the Dominican Republic [Jl. Ecosystems. 2003,6:384-398.
[14] COMLEY B W T,MCGUINNESS K A.Above and below-ground-biomass, and allometrv, of four common northern Australian man—groves[Jl. Australian journal of botany, 2005,53(5):431-436.
[15]曹庆先,徐大平,鞠洪波.基于TM影像纹理与光谱特征和KNN方法估算5种红树林群落生物量[J]林业科学研究,2011,24(2):144-150.
[16]王刚,张秋平,管东生.红树林植物生物量沿纬度分布特征[J]湿地科学,2016,14(2):259-270.
[17]张灿宇,徐猛,房苗,等.入侵物种福寿螺卵斑性状在纬度间的变异[J].生态学杂志,2017,36(6):1628-1636.
[18]周外,刀学琼,邓才明,等.福寿螺的繁殖与生长研究[J].安徽农业科学,2015,43(11):134-135,138.
[19]闫红艳,朱金凤,任业军,等.石门縣福寿螺分布和危害现状及防治对策[J].作物研究,2014,28(6):670-671.
[20]刘军,何跃进,谭济才,等.自然条件下福寿螺繁殖特性[J].应用生态学报,2012,23(2):559-565.
[21]黄鹏,林兆里,徐金汉.福寿螺卵期生物学特性[J].福建农林大学学报(自然科学版),2010,39(1):25-29.
[22]刘巧,杨淞,赵柳兰,等.四川地区外来生物福寿螺的繁殖力[J].生态学杂志,2014,33 (4):1042-1046.
[23]潘颖瑛,董胜张,俞晓平.温度胁迫对福寿螺生长、摄食及存活的影响[J].植物保护学报,2008,35(3):239-244.
[24]周兵,谢春扬迪,闫小红,等.江西省福寿螺的入侵危害现状及稻田发生规律[J].生态与农村环境学报,2015,31(6):902-909.
[25]徐建荣,韩晓磊,张磊.不同地区福寿螺繁殖力比较[J].湖北农业科学,2009,48(12):3015-3017.
[26]李军进,吴龙根,徐裕伦,等.鄞州区农田福寿螺的生物学特性测试[J].安徽农学通报,2008,14( 22):89,96.
[27]韦跃龙,李成展,陈伟海,等.广西岩溶景观特征及其形成演化分析[J].广西科学,2018,25(5):465-504.
[28]张丹.广西典型喀斯特河流沉积物重金属分布、来源及风险评价[D].南宁:广西大学,2014.
[29]林春华,唐赛春,韦春强,等.广西来宾市外来人侵植物的调查研究[J].杂草科学,2015,33(1):38-44.
[30]陈新军.广西北海红树林湿地生态系统定位研究站建设设计[J]陕西林业科技,2018,46(4):106-109.
[31] JOSHI R C.VREYSEN M J B,ROBINSON A S,et al.Problemswith the management of the golden apple snail Pomacea carzalicu-lata:An important exotic pest of ricein Asia[A].RAVINDRA C J.Area-wide control of in-sect pests: From research to field imple-mentation[C].Berlin: Springer Netherlands, 2007.257-264.
[32] WOOTTON R J.The mechanical design of insect wings[J]. Scien-tific American, 1990 .263:1 14-120.
[33] ALBRECHT E, CARRENO N B,CASTRO-VAZQUEZ A.A quan-titative studv of copulation and spawning in the South American ap-ple-snail, Pomacea canaliculata( Prosobranchia: Ampullariidae)[Jl. Veliger, 1996.39(2):142-147.
[34]方麗,秦钟,章家恩,等.福寿螺实验种群生殖力生命表研究[J].水生态学杂志,2011.32(2):105-109.
[35]傅先源,梅玉屏,王洪全光照时间、Ca0浓度和底质对大瓶螺(Ampullaria gigas)生长繁殖的影响[J].广州师院学报(自然科学版),1999.20(5):65-68.
[36] TANAKA K. WATANABE T,HIGUCHI H.Density-dependant-growth and reproduction of the apple snail. Pomacea carzalicula-ta:A density manipulation experiment in a paddy field[Jl. Re-searches on population ecology, 1999,41(3):253-262.
[37]朱丽霞,黄瑶瑶,张泽宏,等.pH、食物和光周期对福寿螺生长发育和繁殖的影响[J].生态学报,2015.35(8):2643-2651.
[38]郭靖,章家恩,赵本良,等.饥饿胁迫下福寿螺的存活和产卵情况及器官组织损伤观察[J].生物安全学报,2013,22(4):242-247.
[39] TAMBURI N E.MARTiN P R.Effects of food availability on repro—ductive output, offspring quality and reproductive efficiencv in theapple snail Pomacea canaliculata[J].Biological invasions, 2011,13( 10):2351-2360.
[40]储少媛,章家恩,郭靖,等.不同插竿方式诱集稻田福寿螺产卵的效应研究[J].华南农业大学学报,2018,39(5):39-46.
[41]毛琦,朱豪红,党绍东,等.稻田福寿螺空间分布型和抽样技术的研究[J].广西植保,2011,24(1):11-15.
[42]贤振华,覃荣乐,冷付春,等.稻田福寿螺为害程度上升原因分析及防控策略[J].广西农业科学,2009,40(8):1007-1009.
[43] ESTEBENET A L,MARTIN P R.Pomacea canaliculata( Gastrop—oda: Ampullaridae): Life-history traits and their plasticity[J]Bioce11.2002.26(1):83-89.
[44]杨爱娟,谭济才,袁哲明,等.福寿螺产卵高度与克氏原螯虾捕食卵块的研究[J].中国农学通报,2012,28(8):90-93.
[45]钟 锋.我国福寿螺防治技术研究进展[J].世界农药,2015,37(6):21-24.
[46]宋红梅,胡隐昌,牟希东,等.外来人侵生物福寿螺的生物学特性、危害与防治现状[J].广东农业科学,2009(5):106-108,110.
[47]杨海芳,杨姗萍,王沛,等.福寿螺在中国的潜在地理分布区预测[J].江西农业学报,2018,30(3):70-73.
基金项目:广西自然科学基金重大专项(201IGXNSFE018005);广西研究生教育创新计划项目(XYCSZ 2018057)
作者简介:梁碧霞(1993-),女(壮族),广西南宁人,硕士,主要从事动物生态学、外来人侵物种的研究,(电话)18677356413(电子信箱)lei.x@foxmail.com;通信作者,武正军,(电子信箱)wuzhengjun@aliYun.com。
