马铃薯黑痣病菌嘧菌酯低敏感菌株的Cyt b序列分析
2020-06-19房雅丽张治家贾晶晶周瑶瑶韩鹏杰
房雅丽,张治家,贾晶晶,周瑶瑶,邢 鲲,韩鹏杰
(山西省农业科学院 植物保护研究所,山西 太原 030031)
马铃薯黑痣病是危害马铃薯生产的土传病害之一,由RhizoctoniasolaniKühn[有性态Thanatephoruscucumeris(A.B.Frank) Donk]引起,主要危害幼芽、茎基部和块茎等,严重时造成整个植株死亡[1]。近几年,由于马铃薯规模化种植的发展以及重茬连作、远距离调种和种植方式的改变等造成马铃薯黑痣病上升为主要病害[2]。在严重发生年份,黑痣病造成的植株死亡率高达90%以上[3]。目前,马铃薯黑痣病的主要防治方法为化学药剂防治,其中甲氧基丙烯酸酯类杀菌剂 (QoI)嘧菌酯对马铃薯黑痣病的防效较好[4-5],然而,病原菌较易对嘧菌酯产生抗药性,其属于高抗药性风险杀菌剂[6]。因此,开展田间菌株对嘧菌酯的敏感性测定和抗性风险评估,筛选防治马铃薯黑痣病的生物源杀菌剂,对促进马铃薯生产发展具有重要意义。
嘧菌酯属于QoI类杀菌剂,QoI主要通过结合在细胞色素b(Cyt b)的Qo位置抑制线粒体的呼吸,从而抑制病原菌孢子萌发和菌丝生长[7]。有些病原菌在离体条件下可以通过旁路氧化途径(AOX)躲避嘧菌酯的抑制,因此,通常在测试对嘧菌酯的敏感性时加入水杨肟酸(SHAM)抑制AOX途径[8]。目前,已报道了4个马铃薯黑痣病菌菌株对嘧菌酯的敏感性[3-4,9],其EC50值差异较大,但目前尚不能确定其差异较大的原因。此外,ARABIAT等[10]研究结果显示,来自甜菜的立枯丝核菌对嘧菌酯的敏感性变异较大,EC50值为0.43~597.43 μg/mL,平均为49.7 μg/mL。
嘧菌酯作为第一个QoI类杀菌剂,在1996年进入市场,而后陆续报道了多种植物病原菌对其产生不同程度的抗性。产生抗性的机制与Cytb基因的点突变有关,目前,已报道的主要突变位点有3个,分别位于143位、129位和137位密码子,其中143位密码子由甘氨酸突变为丙氨酸的频率较高,并且导致高抗性的发生,如Alternariaspp.、Microdochiumnivale、M.majus、Podosphaerafusca和Pseudoperonosporacubensis等病原菌[11-13];而129位密码子由苯丙氨酸突变为亮氨酸、137位密码子由甘氨酸突变为精氨酸的频率较少,导致的抗性水平较低[14-16]。但是,也有部分植物病原菌表现对QoI敏感性下降却未在Cytb基因的上述3个位点检测到突变[6,17-18]。最近报道显示,在突尼斯马铃薯产区马铃薯黑痣病菌对嘧菌酯产生抗性,但未检测抗性菌株的Cytb基因序列是否发生改变[19]。
2007—2016年,我国化学农药商品使用量年均为175.8万t,全世界排名第一[2]。我国从2015年开始实施《到2020年农药使用量零增长行动方案》,生物源农药替代化学农药配套技术成为化学农药减施主要技术方案之一[20]。然而,目前我国登记的使用对象为马铃薯的生物源杀菌剂仅有3种,分别为苦参碱、枯草芽孢杆菌(Bacillussubtilis)和丁子香酚,主要用于晚疫病防治,没有用于防治马铃薯黑痣病的生物源农药[2]。
目前,尚不清楚对嘧菌酯表现低敏感的马铃薯黑痣病菌菌株Cytb基因序列是否发生抗性相关突变及其抗性风险,同时,关于生物源杀菌剂对马铃薯黑痣病菌的室内毒力测定研究较少。鉴于此,通过菌丝生长速率法测定分离得到的马铃薯黑痣病菌对嘧菌酯的敏感性以及SHAM在马铃薯黑痣病菌对嘧菌酯敏感性测定中的作用,分析高敏感和低敏感菌株的Cytb序列,初步探究其低敏感性的机制,此外,测定了4种生物源杀菌剂对低敏感马铃薯黑痣病菌菌株的室内毒力,为田间防治马铃薯黑痣病提供参考。
1 材料和方法
1.1 试验材料
1.1.1 供试菌株 供试15株立枯丝核菌(R.solani)分离自马铃薯黑痣病病薯,所有菌株采自内蒙古自治区呼和浩特市武川县。
1.1.2 供试药剂 本试验共使用7种药剂进行室内毒力测定,分别为0.3%苦参碱水剂(河北省农药化工有限公司)、30%乙蒜素乳油(河南中威高科技化工有限公司)、5%氨基寡糖素水剂(江苏克胜集团股份有限公司)、1%蛇床子素水剂(内蒙古清源保生物科技有限公司)、250 g/L嘧菌酯悬浮剂(先正达作物保护有限公司)、10%苯醚甲环唑水分散粒剂(先正达作物保护有限公司)、75%百菌清可湿性粉剂(利民化工股份有限公司)。SHAM购自上海麦克林生化科技有限公司。Bioeasy Taqman master mix购自博日生物科技有限公司。
1.2 试验方法
1.2.1 马铃薯黑痣病菌对供试药剂的敏感性测定 采用菌丝生长速率法测定供试药剂对马铃薯黑痣病菌的EC50值。将供试药剂用无菌水稀释成系列浓度,加入PDA培养基中配制成一定浓度梯度的含药平板,以加入等体积无菌水的平板为对照。供试药剂的终浓度梯度如下:苦参碱的终质量浓度分别为1、2、5、15、30 μg/mL;乙蒜素的终质量浓度分别为1、2、5、10、15 μg/mL;氨基寡糖素的终质量浓度分别为5、25、100、250、500 μg/mL;蛇床子素的终质量浓度分别为1.25、2.5、5、10、20 μg/mL;苯醚甲环唑的终质量浓度分别为2、5、10、20、30 μg/mL;百菌清的终质量浓度分别为0.5、1.5、3、6、10 μg/mL;因菌株的敏感性不同设定了2种嘧菌酯质量浓度梯度,分别为2、4、8、16、32 μg/mL和0.1、0.3、0.5、0.8、1.0 μg/mL。根据JIN等[21]的报道,SHAM在嘧菌酯对R.solani的抑制效果中的加和效应不明显,因此,在最初测试马铃薯黑痣病菌对嘧菌酯的敏感性时未在含嘧菌酯的平板中加入SHAM。供试菌株在PDA培养基上生长3 d 后,沿菌落边缘打取直径为5 mm的菌饼转接至含药平板中心,在25 ℃黑暗条件下培养3 d,用十字交叉法测量菌落直径。按公式(1)计算不同质量浓度药剂对菌丝生长的抑制率。在Microsoft Excel中运用NORMINV函数将抑制率转换为概率值,而后以概率值为纵坐标(y),以供试药剂系列质量浓度的对数为横坐标(x),得到毒力回归方程y=bx+a及相关系数(r),计算EC50值。
菌丝生长抑制率=(对照菌落直径-处理菌落直径)/(对照菌落直径-菌饼直径)×100%
(1)
1.2.2 SHAM对马铃薯黑痣病菌菌丝生长的影响 采用菌丝生长速率法测定SHAM对马铃薯黑痣病菌菌丝生长的影响。将SHAM用甲醇配制成100 mg/mL母液,而后用甲醇稀释成系列浓度,加入PDA培养基中配制成一定浓度梯度的含药平板,使SHAM最终质量浓度分别为0、10、20、40、60、80、100 μg/mL,以加入等体积甲醇(0.1%)的平板为对照。供试菌株在PDA培养基上生长3 d 后,沿菌落边缘打取直径5 mm的菌饼接种至含药平板中心,在25 ℃培养箱中黑暗培养3 d,用十字交叉法测量菌落直径。每个处理重复3次。按公式(1)计算不同质量浓度SHAM对菌丝生长的抑制率。
1.2.3 SHAM在马铃薯黑痣病菌菌株对嘧菌酯敏感性测定中的作用 根据1.2.2中试验结果确定的对马铃薯黑痣病病原菌菌丝生长影响较小(抑制率<20%)的SHAM质量浓度进行后续试验,以确定SHAM在马铃薯黑痣病菌株对嘧菌酯敏感性测定中的影响。将嘧菌酯用无菌水稀释成系列浓度,加入PDA培养基中配制成2、32 μg/mL含药平板,同时加入用甲醇配制的SHAM,以仅加入SHAM的PDA培养基为对照,每个处理重复3次。按照1.2.1中的方法测定菌落直径,并按公式(1)计算SHAM存在条件下嘧菌酯对菌丝生长的抑制率。
1.2.4 马铃薯黑痣病菌DNA的提取 马铃薯黑痣病菌DNA的提取方法参考CENIS[22]的方法:在1.5 mL离心管中加入0.5 mL PDB培养基,接种少量马铃薯黑痣病菌菌丝,在25 ℃下培养3 d后,10 000 r/min离心5 min,去上清,用0.5 mL 1×TE进行洗涤,之后加入0.3 mL快速提取缓冲液,在Qiagen TissueLyserⅡ上进行破碎后加入0.15 mL 3 mol/L NaAc,-20 ℃静置10 min后,12 000 r/min离心10 min,将上清转入另一离心管中,加入等体积异丙醇室温沉淀10 min后,12 000 r/min离心10 min,75%乙醇洗涤后干燥,加入0.04 mL 1×TE溶解备用。
1.2.5 马铃薯黑痣病菌Cytb基因部分序列分析 选取对嘧菌酯敏感性存在差异的8株马铃薯黑痣病菌,用于探寻病原菌对嘧菌酯抗性的分子机制。利用Vector NTI设计引物RsCytb143-F/RsCytb143-R和RsCytb129-F/RsCytb129-R(表1),扩增与嘧菌酯抗性相关的区域。将扩增得到的Cytb部分序列与立枯丝核菌线粒体基因组(Rhizoctoniasolanistrain AG3 Rhs1AP,NC_021436.1)进行比对。PCR反应总体系为30 μL,包含1 ×Bioeasy Taqman master mix,200 nmol/L正向引物和反向引物,以及基因组DNA模板。PCR 反应程序为:94 ℃ 2 min;94 ℃ 15 s,55 ℃ 30 s,72 ℃ 2 min,30个循环;72 ℃延伸2 min。用1%琼脂糖凝胶电泳检测,将纯化回收后的PCR产物送至英潍捷基(上海)贸易有限公司进行测序。通过DNAstar软件采用Cluster W方法进行序列比对分析。
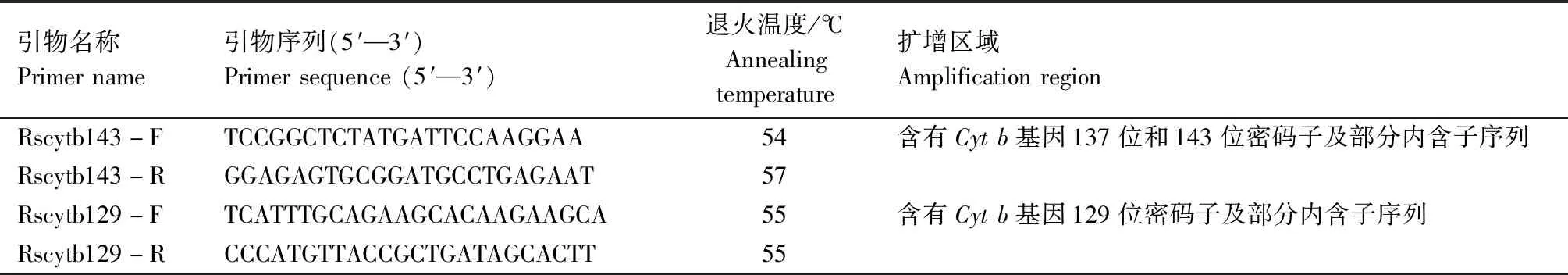
表1 立枯丝核菌Cyt b基因分子特征分析所用引物Tab.1 Primers used in molecular characterization of cytochrome b(Cyt b) gene from R.solani
2 结果与分析
2.1 马铃薯黑痣病菌对嘧菌酯的敏感性
由表2可见,在测试的15株马铃薯黑痣病菌中,H15对嘧菌酯较为敏感,嘧菌酯质量浓度为2、32 μg/mL时,对H15的抑制率分别为64.76%、75.62%,而对其他14株低敏感菌株的抑制率分别为3.62%~25.78%、9.50%~50.96%。当嘧菌酯质量浓度升高时,对H14菌株的抑制率增加不明显,而对其他14株菌株的抑制率显著增加。嘧菌酯对H15的EC50值为0.49 μg/mL,对H9的EC50值接近32 μg/mL,而对其余13株菌的EC50值均>32 μg/mL。结果表明,在武川县分离的15株马铃薯黑痣病菌中,14株菌对嘧菌酯的敏感性较低。
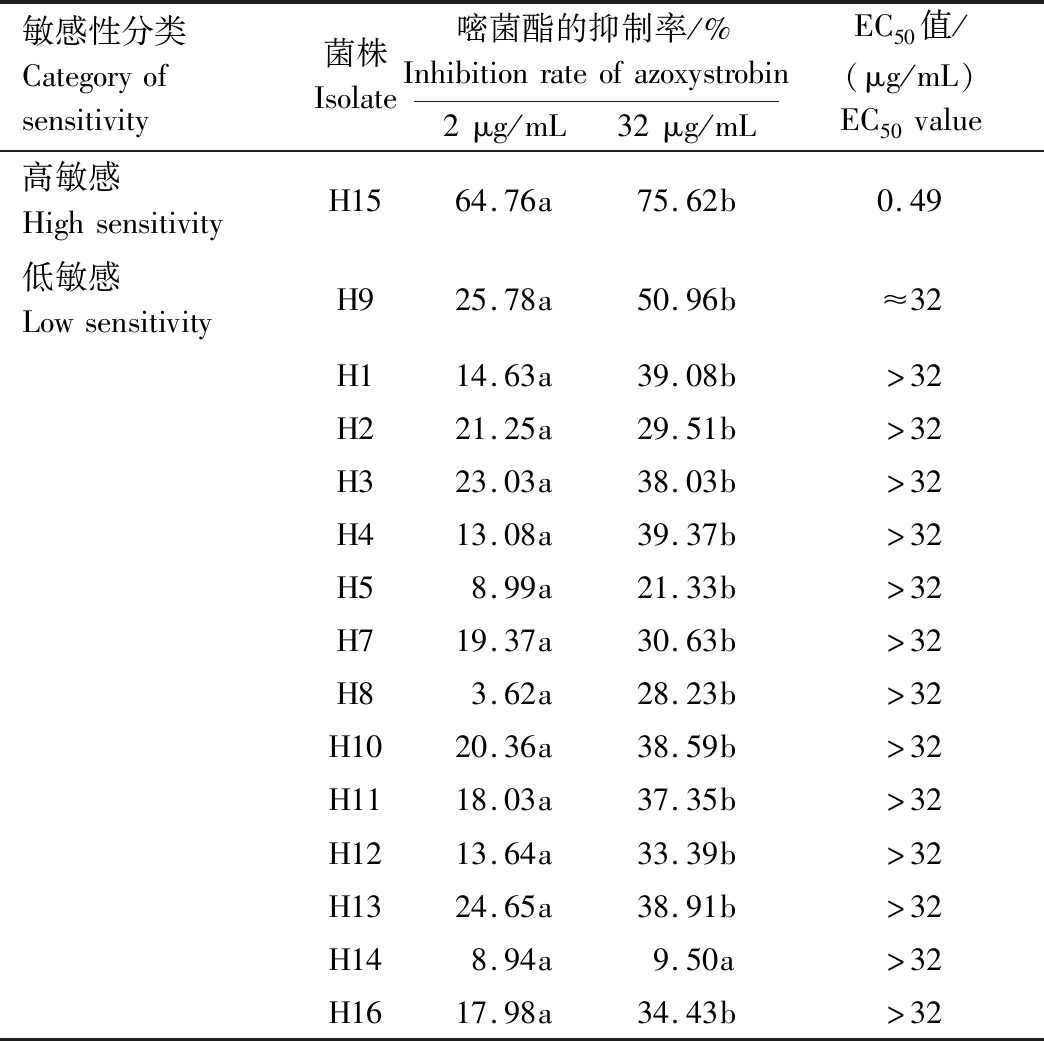
表2 马铃薯黑痣病菌对嘧菌酯的敏感性Tab.2 Sensitivity of R.solani to azoxystrobin
注:同行不同小写字母表示在5%水平差异显著。
Note:Different lowercase letters in the same row indicate significant differences at 5% level.
2.2 水杨肟酸对马铃薯黑痣病菌菌丝生长的影响
SHAM对马铃薯黑痣病菌具有一定的抑制作用,随着质量浓度的升高,抑制作用增加。当SHAM质量浓度为10、20、40 μg/mL时,抑制率分别为3.85%~4.92%、3.85%~17.29%和9.62%~18.18%。当SHAM质量浓度大于40 μg/mL时,对H9的抑制率≥32.33%(表3)。根据已有研究报道,SHAM的最佳质量浓度应对菌丝生长具有一定抑制作用,但抑制率小于20%[23],因此,后续选择SHAM质量浓度为40 μg/mL进一步测定马铃薯黑痣病菌对嘧菌酯的敏感性。
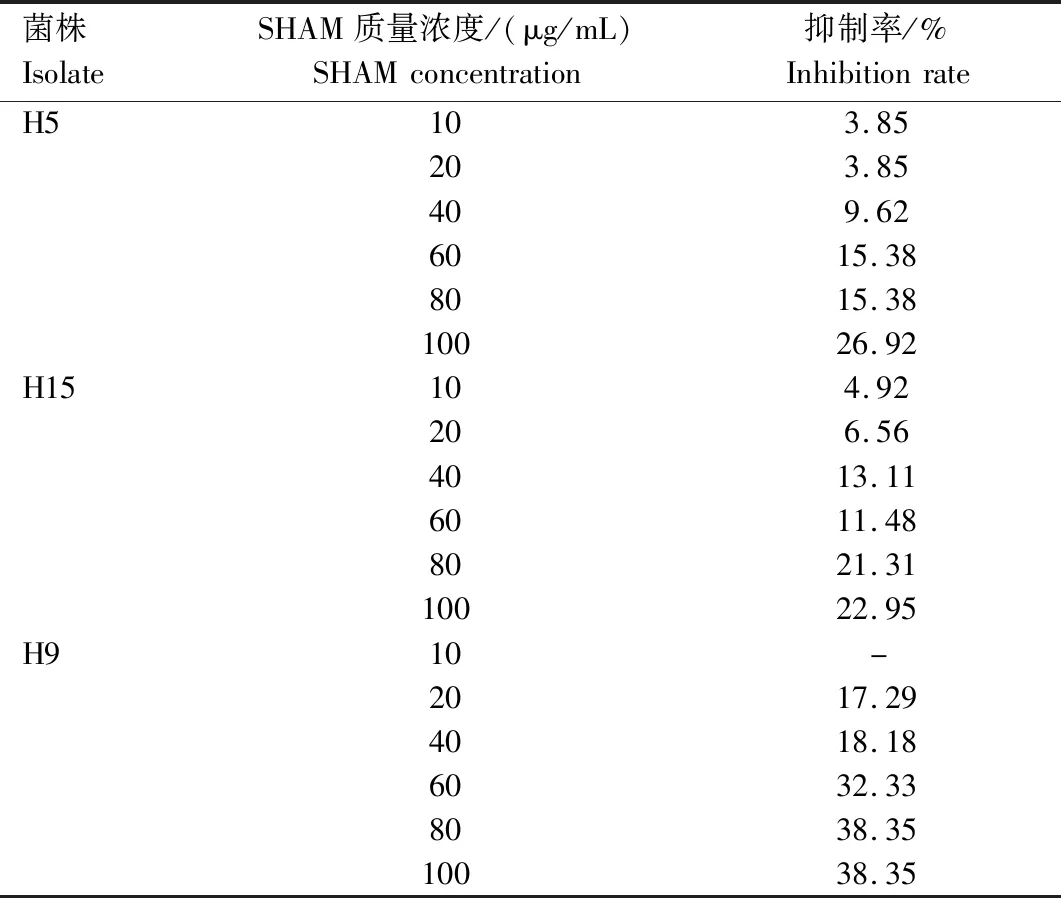
表3 不同质量浓度水杨肟酸对马铃薯黑痣病菌的抑制率Tab.3 The inhibitory rate of SHAM to R.solani at different concentration
注:-表示未进行测试。Note:- means untested.
2.3 水杨肟酸在马铃薯黑痣病菌对嘧菌酯敏感性测定中的作用
为了确定对嘧菌酯低敏感的马铃薯黑痣病菌菌株是否是因为存在旁路呼吸途径躲避QoI的抑制作用,加入SHAM阻断旁路氧化途径后,测试部分马铃薯黑痣病菌对嘧菌酯的敏感性。
由表4可知,添加SHAM后,嘧菌酯对马铃薯黑痣病菌的抑制率有一定程度的升高,高敏感菌株H15的EC50值由0.49 μg/mL降低为0.037 μg/mL,低敏感菌株中只有H9菌株的EC50值下降,其余菌株在嘧菌酯质量浓度为2、32 μg/mL时,抑制率显著增加,但EC50值仍然>32 μg/mL。可见,SHAM可以提高马铃薯黑痣病菌对嘧菌酯的敏感性,但对于不同菌株其效果有一定差别。

表4 添加及不添加水杨肟酸条件下马铃薯黑痣病菌对嘧菌酯的敏感性Tab.4 Sensitivity of R.solani to azoxystrobin with or without SHAM
注:同行不同小写字母表示分别在添加、不添加SHAM条件下相同质量浓度密菌酯的抑制率差异显著(P<0.05)。
Note:Different lowercase letters in the same row indicate significant differences of inhibition rate at the same concentration of axoxystrobin between with and without SHAM.
2.4 马铃薯黑痣病菌Cyt b序列特征分析
将测序结果与立枯丝核菌线粒体基因组(NC_021436.1)的Cytb序列进行比对分析,根据其注释结果,在与嘧菌酯抗性相关的G143位密码子后无内含子插入(图1A)。扩增高敏感的H15菌株和低敏感的7株马铃薯黑痣病菌基因组中的序列,其中包含3个抗性位点(F129、G137、G143)及部分内含子。比对结果显示,所有菌株Cytb基因的G143位密码子后无内含子插入,并且在上述3个位点上碱基序列一致,未出现与抗性相关的点突变(图1B)。

A.细胞色素b部分序列结构,方框表示外显子,线型表示内含子,虚线方框表示包含与QoI抗性相关的3个密码子;B.对嘧菌酯敏感性存在差异的8株马铃薯黑痣病菌株Cyt b基因部分序列比对结果,虚线方框表示129位、127位和143位密码子A.Sequence structure in the partial Cyt b fragment.Boxes indicate exons;Line indicates intron;Dotted line boxes illustrates three codons conferring QoIs resistance.B.Alignment of the partial nucleotide sequences of the Cyt b gene from 8 R.solani isolates.Dotted line boxes indicate codon 129,137 and 143图1 马铃薯黑痣病菌细胞色素b(Cyt b)基因的部分序列分析Fig.1 Characterization of parital cytochrome b gene(Cyt b) sequence obtained from R.solani
2.5 马铃薯黑痣病菌对生物源杀菌剂的敏感性
以百菌清和苯醚甲环唑为对照药剂,选择对嘧菌酯表现低敏感菌株H2测试其对4种生物源杀菌剂的敏感性,百菌清、苯醚甲环唑对H2菌株的EC50值分别为3.50、9.24 μg/mL,苦参碱、乙蒜素、氨基寡糖素、蛇床子素对H2菌株的EC50值分别为15.81、7.72、116.58、4.57 μg/mL(表5)。生物源药剂中,蛇床子素和乙蒜素的效果最好,其次为苦参碱,氨基寡糖素的抑菌率最低。蛇床子素和乙蒜素的抑菌效果与化学药剂百菌清和苯醚甲环唑相似,可以作为防治马铃薯黑痣病的备选药剂。
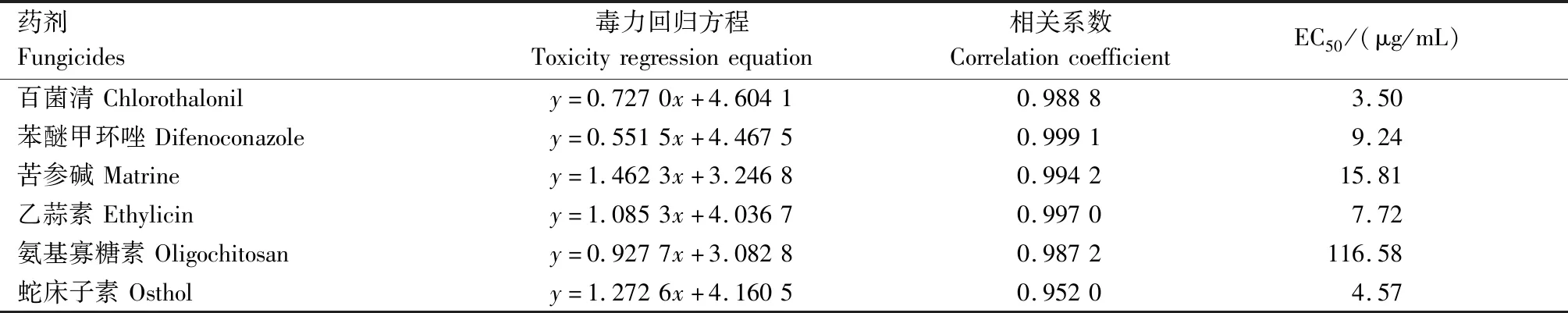
表5 几种生物源杀菌剂对马铃薯黑痣病菌的抑制水平Tab.5 Inhibition of several biological fungicides to R.solani
3 结论与讨论
嘧菌酯自1996年问世以来,用于多种植物病害的防治。目前,已经报道了4个马铃薯黑痣病菌株对嘧菌酯的敏感性,EC50值分别为4.45×10-9μg/mL[9]、8.32 μg/mL[3]、0.08 μg/mL和0.05 μg/mL[4]。其中,张智芳等[9]和刘宝玉等[3]采用菌丝生长速率法测定菌株对嘧菌酯的敏感性,与本研究采用的方法相同,与本试验中EC50值0.49 μg/mL和≥32 μg/mL相比,嘧菌酯对马铃薯黑痣病菌菌株的EC50值变异更大。由于在离体条件下,病原菌可以通过旁路氧化途径躲避嘧菌酯的抑制,故在菌株敏感性测试中加入SHAM,试验结果表明,加入SHAM后马铃薯黑痣病菌对嘧菌酯的敏感性增加,但对于低敏感菌株H5和H16敏感性的增加幅度不大,EC50值仍>32 μg/mL。来自甜菜的R.solani对嘧菌酯的敏感基线已经建立,EC50值为0.43~597.43 μg/mL,平均为49.70 μg/mL,活体试验显示,推荐剂量的嘧菌酯可以有效控制高EC50值菌株的侵染,推测较高的EC50值可能是由于存在旁路氧化途径的其他机制所造成的,该机制不同于目前已报道的可被SHAM抑制的旁路呼吸途径[10]。本试验中,嘧菌酯对马铃薯黑痣病菌的EC50值也存在变异较大的现象。嘧菌酯对在突尼斯分离到的4株马铃薯黑痣病菌的EC50值大于30 μg/mL,推测其可能是抗性菌株[19]。然而,本试验检测了7株低敏感和1株高敏感菌株的Cytb序列,在已报道的3个Cytb主要突变位点上未检测到点突变。虽然植物病原菌对嘧菌酯的抗性可能存在其他机制,例如,M.fructicola对嘧菌酯表现为敏感性下降,然而其Cytb基因上未检测到与嘧菌酯抗性相关的点突变[17,24],但是根据来自甜菜的R.solani对嘧菌酯的敏感性研究结果,对嘧菌酯表现低敏感的马铃薯黑痣病菌株可能需要进一步活体试验确定是否为抗性菌株,有可能是由于存在旁路氧化途径的其他机制使得离体条件下马铃薯黑痣病菌对嘧菌酯的敏感性较低。
对嘧菌酯表现高抗的菌株一般是发生了Cytb基因143位密码子的点突变,F129和G137位点的突变产生低或者中等抗性,目前报道高抗病原菌的Cytb基因G143位密码子后没有插入内含子,在Cytb基因G143密码子后有内含子插入的病原菌不容易产生高抗菌株[24-25]。G143位密码子后插入的内含子为Ⅰ型内含子,可以进行自我剪切,而G143位密码子位于该内含子的内在前导序列(Internal guide sequence,IGS)中,该位点的突变会影响Cytb基因mRNA的正确剪接,使得突变菌株死亡,因此这类病原菌不易产生对嘧菌酯的抗性[26],而本试验中,R.solani中G143位密码子后没有内含子,暗示马铃薯黑痣病菌较容易产生对嘧菌酯的抗性。因此,本研究测试了4种生物源杀菌剂对马铃薯黑痣病菌的抑制效果,以作为防治马铃薯黑痣病的备选药剂。其中,蛇床子素和乙蒜素的效果较好,与百菌清和苯醚甲环唑的效果相似,其次为苦参碱。氨基寡糖素的抑菌效果较差,但其具有激发植物免疫反应的作用,之后可通过活体试验确定其防治效果。这4种生物源杀菌剂中蛇床子素和乙蒜素可作为防治马铃薯黑痣病菌的候选药剂。
