鸡白细胞介素2和白细胞介素18基因的原核及真核表达
2020-05-25张艳雯张鹤曦周晴云唐子琼
张艳雯,张鹤曦,覃 静,周晴云,唐子琼
(1.南宁学院,广西 南宁 530200;2.钦州市畜牧站,广西 钦州 535099)
【研究意义】白细胞介素2(IL-2)是一种由激活T淋巴细胞产生的生长因子,也是CD4+和CD8+效应T细胞增殖和分化的关键因子[1-3],可通过诱导细胞毒性T细胞、淋巴因子激活的杀伤细胞及自然杀伤细胞促进T细胞产生的相关细胞因子表达,特异性增强细胞免疫反应[4]。白细胞介素18(IL-18)是由激活的单核细胞、树突状细胞和巨噬细胞产生的一种多功能细胞因子[5],具有诱导NK细胞或丝裂原激活T细胞产生IFN-γ的能力,同时提高NK细胞和T细胞杀伤活性[6-7],在提高机体抗感染能力和识别、清除异化自体组织等方面发挥重要作用[8]。IL-2或IL-18在人体及其他动物上具有广泛的生物学活性和应用价值[9-11],通过原核和真核表达载体制备获得的DNA疫苗可明显增强疫苗的免疫原性[12-14]。鸡白细胞介素2(ChIL-2)和鸡白细胞介素18(ChIL-18)蛋白具有重要的生物学功能,但二者的表达量因试验方法及条件的不同存在明显差异。因此,加强ChIL-2和ChIL-18研究对其蛋白功能、生物学活性及联合应用免疫佐剂的研发具有重要意义。【前人研究进展】Sundick等[15]用ConA刺激活化的鸡脾脏细胞,通过转染COS-7细胞,首次克隆获得ChIL-2基因,并筛选出T细胞增殖活性。KaiserI等[16]采用RACE克隆获得ChIL-2全序列,并对其进行基因组结构分析和染色体定位,结果发现其外显子和内含子结构与哺乳动物极相似,且存在其他潜在的转录因子结合位点。张春杰等[17]从鸡新城疫Ⅰ系疫苗诱导的鸡胚脾淋巴细胞中扩增获得ChIL-2全长cDNA序列,将ChIL-2基因克隆到原核表达载体上,获得重组质粒,重组质粒在大肠杆菌中的诱导表达产物经SDS-PAGE电泳分析,结果表明所克隆的ChIL-2基因在大肠杆菌中得到良好表达。郭川等[18]从鸡脾脏细胞中扩增出ChIL-2编码基因,将该编码基因克隆至原核表达载体并构建了ChIL-2基因的重组原核表达质粒,重组质粒转化大肠杆菌JM109后经诱导表达,分别得到两种表达产物,经过纯化复性后,表达产物具有明显促进鸡脾淋巴细胞增殖的作用。Schneider等[19]首次从脂多糖(LPS)刺激的鸡巨噬细胞系HD-11中扩增并克隆获得ChIL-18基因。Kaiser等[20]报道显示,火鸡与鸡的IL-18基因的编码区高度保守,具有很高的同源性,系统进化树分析表明二者的IL-18在生物学活性上有交叉反应。胡敬东等[21]从LPS刺激10 h活化的MDCC-MSB1细胞中克隆获得ChIL-18成熟活性蛋白的完整基因片段,完成了原核和真核表达。李宏梅等[22]研究发现,利用原核表达的重组鸡rChIL-18蛋白具有提高NK细胞和CTL杀伤活性作用。【本研究切入点】随着分子生物学、免疫学理论和技术的发展,ChIL-2和ChIL-18基因与功能已成为研究的热点,但针对ChIL-2和ChIL-18蛋白表达的研究鲜见报道。【拟解决的关键问题】根据GenBank中ChIL-2和ChIL-18基因序列设计并合成特异性引物,采用RT-PCR扩增ChIL-2和ChIL-18基因,并克隆到pET32a原核载体和pVAX1真核载体上,分别构建原核表达载体pET32a-ChIL-2和pET28a-ChIL-18及真核表达载体pVAX1-ChIL-2和pVAX1-ChIL-18,分别以Western blotting检测确定该蛋白表达量及间接荧光检测ChIL-2和ChIL-18蛋白在细胞中的表达量,为后续ChIL-2和ChIL-18蛋白功能、生物学活性及联合应用免疫佐剂的研发提供参考依据。
1 材料与方法
1.1 试验材料
1.1.1 菌种和克隆载体 大肠杆菌BL21感受态细胞、大肠杆菌DH10B感受态细胞、pET32a原核表达载体、pET28a原核表达载体、pVAX1真核表达载体和pMD19-T 载体购自大连宝生物工程TaKaRa公司。
1.1.2 主要仪器设备 小型台式离心机、低温高速离心机、移液枪、天平、pH计、电热恒温培养箱、磁力搅拌器、高压灭菌锅、脱色摇床、多用途旋转摇床、制冰机、核酸电泳仪、电泳槽、蛋白电泳仪、PCR仪、水浴锅、超声波细胞破碎机和凝胶成像系统。
1.1.3 工具酶和试剂 OMEGA质粒小提试剂盒、OMEGA质粒大提试剂盒、DNA胶回收试剂盒、常规限制性内切酶(KpnⅠ、SalⅠ、BamHⅠ、EcoRⅠ、PstⅠ、MluⅠ和XbaⅠ)、PCR用DNA聚合酶(Taq酶)、蛋白Marker、dATP(R0141)、T4DNA连接酶、DNA Marker、dNTPs、酵母提取物、NaCl、胰蛋白胨、磷酸盐缓冲液、甘氨酸、Tris碱、甲醇、刀豆蛋白A和四甲基偶氮唑盐。
1.2 试验方法
1.2.1 引物合成 根据GenBank已公布的ChIL-2(GU119890.1)和ChIL-18(NM_204608.1)基因序列,应用Primer 7.0设计扩增ChIL-2和ChIL-18基因的特异性引物(表1),引物委托生工生物工程(上海)有限公司合成。
1.2.2 无特定病原(SPF)鸡脾淋巴细胞分离培养及总RNA提取 30日龄SPF雏鸡购自九江博美莱生物制品有限公司。在无菌条件下,取鸡脾脏后制成单细胞悬液,400 r/min离心15 min,将离心沉淀的细胞悬浮于含10 %胎牛血清的RPMI1640培养液中,取一支新的离心管,先加入4 mL淋巴细胞分离液,再加入等体积的淋巴细胞悬液,用密度梯度离心法,400 r/min离心15 min,分离淋巴细胞;再用RPMI1640培养液在350 r/min离心条件下离心洗涤淋巴细胞2次后,用台盼蓝染色法进行活细胞计数,将细胞浓度调整至1×107个/mL,并加至6孔细胞培养板中,每孔加2 mL,置于40 ℃、5 % CO2细胞培养箱中培养。分别于培养的第6和20 h收集培养细胞,于4 ℃下400 r/min条件下离心15 min,用1 mL RPMI1640培养液重新悬浮细胞,用TRIzol试剂提取淋巴细胞总RNA,用1 %琼脂糖凝胶电泳检测总RNA完整性。提取的淋巴细胞总RNA置于-80 ℃冰箱保存备用。

表1 扩增ChIL-2和ChIL-18基因的特异性引物
1.2.3 ChIL-2和ChIL-18基因克隆与鉴定 用AMV反转录酶和Oligo(dT)15引物将总RNA反转录合成cDNA,再以cDNA为模板,采用表1中引物分别进行PCR扩增,扩增程序:98 ℃预变性30 s;98 ℃ 10 s,55 ℃ 30 s,72 ℃ 30 s ,进行33个循环;72 ℃延伸10 min。反应结束后用1 %琼脂糖凝胶电泳对PCR扩增产物进行检测,参考DNA胶回收试剂盒说明回收PCR产物,分别获得ChIL-2 和ChIL-18基因片段。将回收的ChIL-2和ChIL-18基因片段分别克隆到pMD19-T载体,并转化DH10B感受态细胞。参考OMEGA质粒提取试剂盒说明分别小量提取重组质粒pMD19-T-ChIL-2和pMD19-T-ChIL-18后,选择合适的酶切位点进行酶切鉴定和测序。
1.2.4 ChIL-2和ChIL-18基因原核表达 以重组质粒pMD19-T-ChIL-2和pMD19-T-ChIL-18为模板,用表2中引物分别进行PCR扩增,PCR产物进行胶回收,获得纯化的ChIL-2和ChIL-18基因片段。用BamH I和SalⅠ分别双酶切pET32a和pET28a载体。在22 ℃水浴条件下,用T4 DNA连接酶将ChIL-2基因片段与载体pET32a、ChIL-18基因片段与载体pET28a进行连接反应4 h,并转化DH10B感受态细胞。将感受态细胞涂布于含100 μg/mL氨苄青霉素的LB培养基上,置于37 ℃培养箱中过夜培养。挑选单个菌落,接种至2 mL LB液体培养液中,置于37 ℃气浴摇床中振荡培养。参考OMEGA质粒提取试剂盒说明对菌液进行质粒小量提取,得到阳性重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18,分别用PCR和双酶切鉴定并测序。将重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18转化至BL21中感受态细胞,涂板后37 ℃过夜。次日挑取单个菌落接种至LB液体培养基中,待菌液浓度A600=1.5时,加入IPTG至终浓度0.1 mmol/L,20 ℃诱导培养过夜。取部分菌液进行SDS-PAGE分析和Western blotting鉴定。
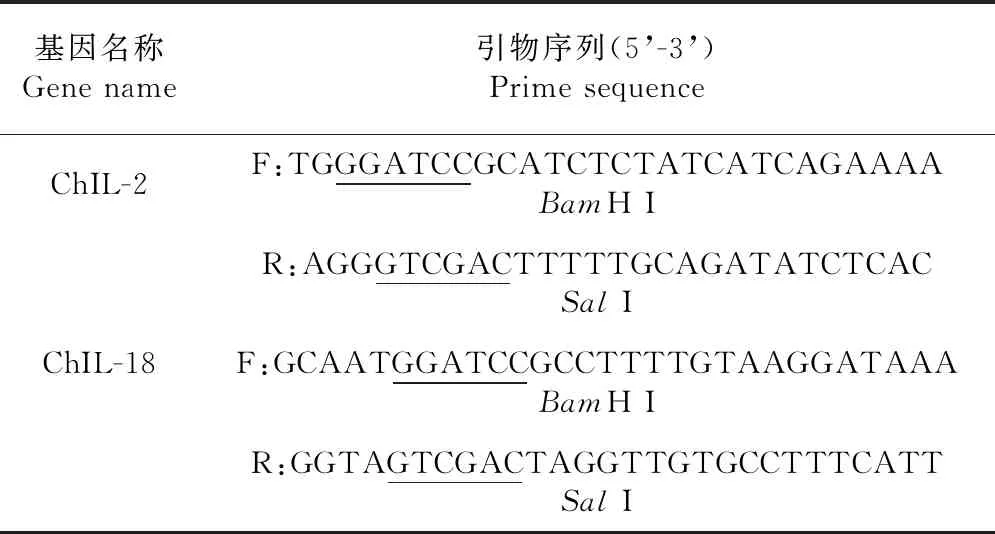
表2 构建重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18的引物
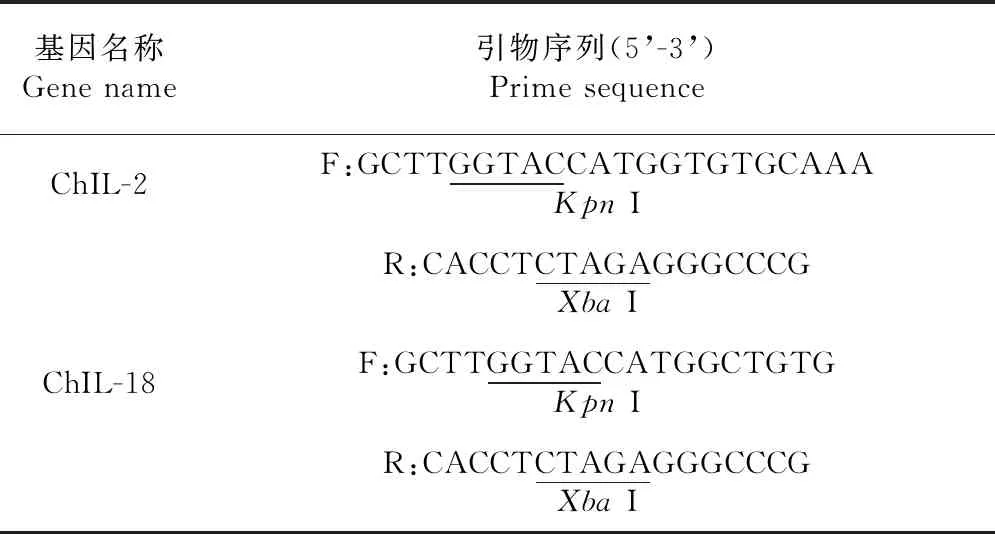
表3 构建重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18的引物
1.2.5 ChIL-2和ChIL-18基因真核表达 以重组质粒pMD19-T-ChIL-2和pMD19-T-ChIL-18为模板,用表3中的引物进行PCR扩增,将PCR扩增产物进行胶回收,获得纯化的ChIL-2和ChIL-18基因片段。用KpnI和XbaI双酶切pVAX1载体,在22 ℃水浴条件下,用T4DNA连接酶将ChIL-2和ChIL-18基因片段与pVAX1载体进行连接反应,并转化DH10B感受态细胞,将感受态细胞涂板后37 ℃过夜培养。参照OMEGA质粒小提试剂盒说明少量提取质粒,分别获得重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18。将重组真核表达质粒分别进行PCR、酶切鉴定和测序。将重组真核表达质粒分别转染非洲绿猴肾(Vero)细胞,24 h后用倒置荧光显微镜观察其蛋白表达情况。
2 结果与分析
2.1 ChIL-2和ChIL-18基因克隆结果
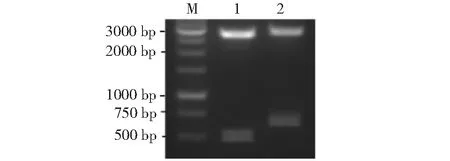
2.1.1 ChIL-2和ChIL-18基因PCR扩增结果 从图1的1和2泳道可见1条清晰特异性条带,其中,1泳道的条带约在400 bp处,2泳道的条带约在600bp处,与预测的目的基因片段大小相符。

M:DNA Marker;1:ChIL-2;2:ChIL-18
2.1.2 重组质粒pMD19-T-ChIL-2和pMD19-T-ChIL-18酶切鉴定结果 对构建的重组质粒pMD19-T-ChIL-2和pMD19-T-ChIL-18经BamH I和SalI双酶切,结果显示,1和2泳道的目的条带大小与预测结果相符(图2)。
2.1.3 ChIL-2和ChIL-18基因的测序 测序结果显示,ChIL-2和ChIL-18基因片段全长分别为429和593 bp,采用DNASTAR对其核苷酸序列及推导氨基酸序列进行比对分析,结果发现ChIL-2基因片段编码143个氨基酸,ChIL-18基因片段编码218个氨基酸残基。与NCBI数据库中公布的鸡IL-2(Sequence ID:AAC96064.1)和IL-18(Sequence ID:NP_989939.1)序列进行比对,其相似性分别为99.7 %和100.0 %(图3~4)。
M:DNA Marker;1:重组质粒pMD19-T-ChIL-2;2:重组质粒pMD19-T-ChIL-18
2.2 ChIL-2和ChIL-18蛋白的原核表达情况
2.2.1 重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18鉴定结果 选择PstI对构建的重组原核表达质粒pET32a-ChIL-2进行酶切,选择MluI和SalI对构建的重组原核表达质粒pET28a-ChIL-18进行酶切。从图5可看出,酶切获得的目的条带大小与预期结果相符;测序结果也表明,构建的重组原核表达质粒正确,无基因突变,即已成功构建获得重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18。

1:ChIL-2基因编码区核苷酸序列;2:ChIL-2基因编码区核苷酸推导氨基酸序列;3:NCBI数据库中鸡IL-2基因推导氨基酸序列

1:ChIL-18基因编码区核苷酸序列;2:ChIL-18基因编码区核苷酸推导氨基酸序列;3:NCBI数据库中鸡IL-18基因推导氨基酸序列
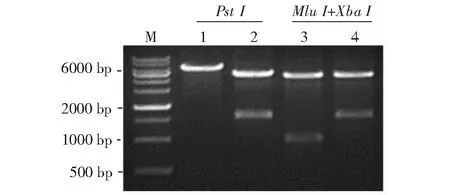
M:DNA Marker;1:pET32a载体;2:重组原核表达质粒pET32a-ChIL-2;3:重组原核表达质粒pET28a;4:重组原核表达质粒pET28a-ChIL-18
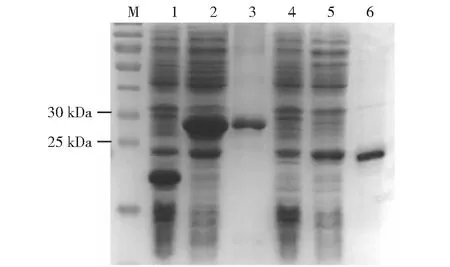
M:蛋白质分子量标准;1:pET32a载体;2:未纯化的重组原核表达质粒pET32a-ChIL-2;3:纯化的重组原核表达质粒pET32a-ChIL-2;4:pET28a载体;5:未纯化的重组原核表达质粒pET28a-ChIL-18;6:纯化的重组原核表达质粒pET28a-ChIL-18
M:蛋白质分子量标准;1:pET32a载体;2:未纯化的重组原核表达质粒pET32a-ChIL-2;3:纯化的重组原核表达质粒pET32a-ChIL-2;4:pET28a载体;5:未纯化的重组原核表达质粒pET28a-ChIL-18;6:纯化的重组原核表达质粒pET28a-ChIL-18
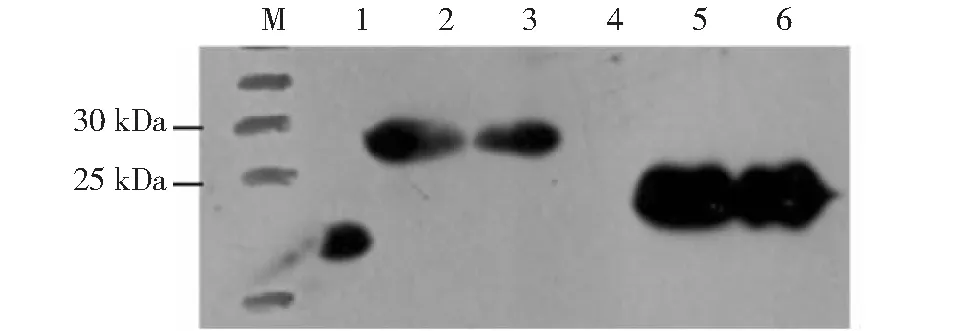
2.2.2 融合蛋白ChIL-2和ChIL-18的诱导表达情况 SDS-PAGE分析结果显示,经IPTG诱导、菌体培养和纯化后的重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18菌体蛋白在27 kD(3泳道)和23 kD(6泳道)处可见1条特异性条带,而空载质粒(1和4泳道)经诱导后未表达任何蛋白,说明诱导的原核菌体成功表达了融合蛋白ChIL-2和ChIL-18(图6)。Western blotting检测结果也显示,在27 kD(3泳道)和23 kD(6泳道)处可见特异性条带,说明原核表达产物即为预期的ChIL-2和ChIL-18(图7)。
2.3 ChIL-2和ChIL-18基因的真核表达情况
2.3.1 重组真核表达质粒 pVAX1-ChIL-2和pVAX1-ChIL-18鉴定结果 对构建的重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18采用KpnI和XbaI进行双酶切,结果目的条带大小与预期结果相符(图8),对应的测序结果序列也正确。说明已成功构建获得重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18。
2.3.2 融合蛋白ChIL-2和pVAX1-ChIL-18在Vero细胞内的表达情况 将所构建的重组真核表达质粒pVAX1-cCHIL-2与pVAX1-cCHIL-18分别转染至Vero细胞中,24 h后取其中3皿(加空白对照)用倒置荧光显微镜检测,结果(图9)显示,与空白对照(图7-C)比较,图7-A和图7-B中均可见荧光,说明融合蛋白ChIL-2和ChIL-18在Vero细胞中均有表达。

M:DNA Marker;1:pVAX1载体;2:重组真核表达质粒pVAX1- ChIL-2;3:重组真核表达质粒pVAX1-ChIL-18
3 讨 论
本研究从鸡脾淋巴细胞中扩增的ChIL-2和ChIL-18基因片段全长分别为429和593 bp,其中,ChIL-2基因片段编码143个氨基酸残基,ChIL-18基因片段编码218个氨基酸残基;氨基酸序列比对分析结果表明,扩增的ChIL-2基因序列与Sundick公布的鸡IL-2基因序列相似性达98.83 %[15],ChIL-18基因序列与GenBank中公布的鸡IL-18基因序列(GenBank:GEU334505.1)相似性为99.83 %,与五彩金刚鹦鹉(Aramacao)(GenBank:GU140075.1)的相似性为99.66 %。可见,本研究已成功扩增到ChIL-2和ChIL-18基因序列。
本研究将构建的重组原核表达质粒pET32a-ChIL-2和pET28a-ChIL-18分别转化大肠杆菌BL21感受态细胞,通过IPTG低温过夜诱导,融合蛋白ChIL-2和ChIL-18的表达量明显增高,表明采用pET32a和pET28a表达系统可高效表达ChIL-2和ChIL-18蛋白。Stepaniak等[23]采用pET32a表达系统高效表达了鸡IL-2蛋白量,高于采用大肠杆菌Pgex-2系统的表达量,且以可溶性蛋白的形式为主,易回收和纯化。Zhou等[24]研究发现,鸭白细胞介素2(DuIL-2)和ChIL-2会与其单克隆抗体起免疫交叉反应,且DuIL-2能促进鸡和鸭两类脾脏淋巴细胞增殖。原核表达蛋白需要提纯,其生物活性也受影响,但已有研究表明,在大肠杆菌中表达的鸡IL-2具有刺激T淋巴细胞增殖的生物学活性;重组的鸡IL-2蛋白可在24 h内引起外周血CD4+和CD8+细胞比例增加[25]。
本研究还成功构建了重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18,将二者转染Vero细胞,24 h后荧光检测发现融合蛋白ChIL-2和ChIL-18均获得表达,不仅可为后续将重组真核表达质粒直接用于体内获得表达,还可与疫苗共注射以提高疫苗的免疫原性。由振强等[26]构建了鸡IL-2真核表达质粒pCI-ChIL-2-EGFP,并与H5亚型禽流感油乳剂灭活苗(AIV)共同注射鸡体内,结果发现,pCI-ChIL-2-EGFP可明显提高AIV的免疫原性。邵攀峰等[27]将ChIL-18基因克隆至真核表达载体pcDNA3.1(+)中,构建重组真核表达质粒pcDNA3.1/ChIL-18(pIL-18),并用pIL-18和ILTV弱毒疫苗及其二者联合分别免疫21日龄SPF雏鸡,结果发现pIL-18与ILTV弱毒疫苗联合免疫组的淋巴细胞亚群CD3+、CD3+CD4+和CD3+CD8+明显高于ILTV弱毒疫苗免疫组,ILTV强毒攻毒后发病鸡数量少,且发病症状较轻。

A:融合蛋白ChIL-2;B:融合蛋白ChIL-18;3:空白对照
4 结 论
成功构建的ChIL-2和ChIL-18基因重组原核表达质粒pET32a-ChIL-2和pET32a-ChIL-18及重组真核表达质粒pVAX1-ChIL-2和pVAX1-ChIL-18,分别在大肠杆菌和Vero细胞中获得高表达,即克隆获得的IL-2和IL-18基因可在原核和真核细胞中高效表达,为后续研究IL-2和IL-18蛋白的融合表达、生物学活性及疫苗的免疫作用打下了基础。
