3种城郊防护林土壤呼吸与温湿度的关系
2020-05-11张成芳李德生魏佳宇赵亮玉
张成芳 李德生 魏佳宇 赵亮玉


摘要: 以天津市3种代表性的城郊防护林(杨树、火炬树、刺槐)为研究对象,利用便携式土壤碳通量全自动分析仪ACE进行定期观测,研究结果表明:(1)在2018年4—10月生长季3种城郊防护林土壤呼吸速率的日变化表现为单峰曲线,最大值出现在10:00—15:00,最小值出现在20:00—5:00。(2)土壤呼吸速率的季节变化为明显的单峰曲线,杨树土壤呼吸速率在7月达到峰值,为3.80 μmol/(m2·s);而火炬树和刺槐土壤呼吸速率在8月达到峰值,分别为3.84、4.75 μmol/(m2·s);杨树、火炬树和刺槐的土壤呼吸速率平均值分别为2.01、2.25、2.62 μmol/(m2·s),差异达显著水平(P<0.05)。(3)土壤呼吸速率与土壤温度之间具有显著的二次函数关系(P<0.05),拟合度为78.8%~84.7%;与土壤湿度之间呈显著或极显著线性正相关,拟合度为66.8%~90.2%。(4)对土壤呼吸速率和10 cm深度的土壤温度(T10)及5 cm深度的土壤湿度(M5)之间进行多元线性拟合,相关系数为0.826~0.950,说明多元线性模型能够很好地解释土壤温度和湿度对土壤呼吸的协同作用,表明土壤温度和湿度是3种城郊防护林土壤呼吸速率的主要影响因子。
关键词: 城郊防护林;土壤呼吸速率;土壤温度;土壤湿度;相关性
中图分类号: S181 文献标志码: A
文章编号:1002-1302(2020)05-0243-07
在全碳循环中,土壤呼吸是碳从生态系统向大气层移动的主要途径[1],大约是人类化石燃料燃烧的10倍以上[2]。由于生物量碳库更容易受到自然因素和人为因素的干扰[3],所以土壤中的固碳可能比现存生物量中的固碳更具稳定性和持久性[4]。土壤中含有大部分的陆地碳,并在陆地碳循环中起着重要作用[5]。因此,研究土壤呼吸及其影响因子对全球气候变化的响应是碳循环研究的一个重要内容。
當今,众多土壤呼吸作用的相关研究在国内外已广泛展开。国外对陆地生态系统土壤呼吸重视较早,大规模的研究始于20世纪70年代,但主要针对耕作土壤即农田生态系统[6]。而我国于20世纪90年代开始土壤呼吸方面的研究,研究对象多集中在我国亚热带、温带的森林、草原等生态系统以及不同土地利用方式等,但涉及城市森林土壤呼吸方面的研究相对较少[7]。城市森林作为吸收CO2、释放O2的一个巨大碳汇,是生物群中对地球初级生产的最大贡献者,在全球碳循环、减缓温室效应中起着非常重要的作用[8]。土壤呼吸是陆地生态系统中仅次于光合作用的第二大碳循环途径[9],森林生态系统碳循环过程,尤其是土壤呼吸对于陆地生态系统的净碳收支具有重要作用[10]。
在新时代推动生态文明建设的过程中,城郊防护林成为城市森林生态系统的重要组成部分。天津市城郊防护林广泛使用的树种有杨树(Populus)、刺槐(Robinia pseudoacacia)和火炬树(Rhus typhina),其对城市森林生态系统土壤碳循环具有重要作用。因此,本研究选择天津地区3种代表性的城郊防护林为研究对象,利用便携式土壤碳通量全自动分析仪ACE测量城市森林生态系统土壤碳排放及其影响因素,分析3种城郊防护林环境因子与土壤呼吸的相关关系,旨在为有效维护城市生态环境的良好状况以及城市健康的持续发展和为更好地探索城市森林生态系统土壤碳排放及其影响因子之间的相互关系提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于天津市西青区(117.132~117.136°E,39.56~39.64°N),属暖温带半湿润季风性气候,四季分明,春季温暖少雨;夏季炎热多雨水;秋季冷暖适中;冬季寒冷干燥。年平均气温约为14 ℃,极端高温达41.6 ℃,极端低温为 -17.8 ℃,7月最热,月平均温度为28 ℃;1月最冷,月平均温度为 -2 ℃。年平均降水量为 584.6 mm,年内50%的雨日和75%以上的降水量集中在夏季,植物生长季为4—10月。天津市西青区总面积是545 km2,林地面积达到120.6 km2。3种城郊防护林基本概况见表1。
1.2 土壤呼吸及土壤温湿度的测定
本研究选择3种城郊防护林,每种防护林设1个固定的样地(100 m×100 m),每个样地中布设3个监测点做平行试验,于2018年4—10月(植物生长初期、中期和末期)。在测量前提前24 h将钢圈(直径30 cm、高8 cm)嵌入离树干约0.5 m的土壤内,并清除环中植被以及凋落物,以避免土壤扰动及地上植被呼吸对监测结果产生影响。采用便携式土壤碳通量全自动分析仪ACE(EN110NT,ADC BioScientific Ltd.,UK)同时测定土壤呼吸速率、土壤温度及土壤湿度。土壤呼吸速率日变化的测定为测量当日的08:00至次日的08:00,以保证24 h的连续监测,仪器每30 min测定一组数据,间隔 30 min 后自动监测下组数据。10 cm深度的土壤温度(T10)和5 cm深度的土壤湿度(M5)由便携式土壤碳通量自动分析仪ACE配套的土壤温度探针及湿度探针分别进行测定。
1.3 数据分析
原始数据经Excel 2007整理后,采用SPSS 21.0进行统计分析,单因素方差分析(One-way ANOVA)不同月份、不同防护林的土壤呼吸、土壤温度、土壤湿度的相关性,显著性差异水平设定为P<0.05,同时采用二次曲线方程和线性方程分别对土壤呼吸速率与土壤温度和湿度间进行回归拟合并得到拟合度参数R2。采用Origin 9.0进行绘图。
2 结果与分析
2.1 3种城郊防护林土壤温湿度的变化
由图1-A可知,3种城郊防护林的T10生长季变化趋势基本保持一致,呈现出明显的单峰变化趋势。从2018年4月开始,T10上升,到7月均达到最大值,此后开始呈下降趋势且下降范围较大。杨树、火炬树和刺槐的T10变化范围分别为12.1~27.5 ℃、13.2~27.8 ℃、12.6~28.1 ℃,生长季T10的平均值分别为21.8、22.4、21.7 ℃(图1-B)。显然,3种城郊防护林生长季T10排序为火炬树>杨树>刺槐,但三者差异并不显著(P>0.05),这说明土壤温度的季节变化规律与气温相同,呈现出春秋低、夏季高的格局。3种城郊防护林的M5生长季变化趋势呈现明显差异(图2)。杨树、火炬树和刺槐三者的生长季M5变化范围分别为:25.5%~58.6%、23.3%~45.9%、28.5%~61.3%。显然,3种城郊防护林生长季M5平均值排序为杨树>刺槐>火炬树,且火炬树的M5平均值显著低于刺槐和杨树的M5平均值(P<0.05)。
2.2 3种城郊防护林土壤呼吸速率特征
2.2.1 3种城郊防护林土壤呼吸速率的日变化
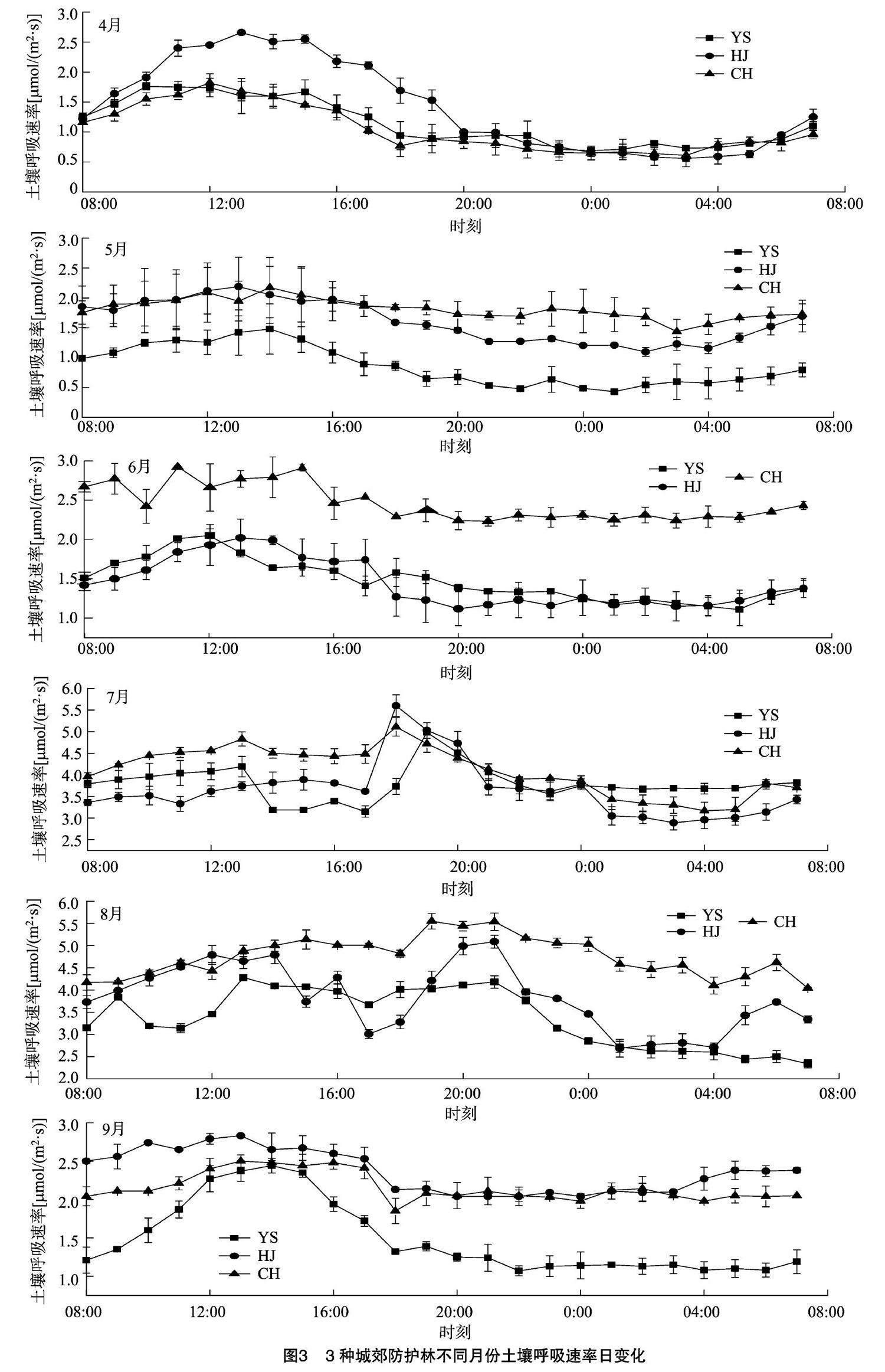
由图3可知,3种城郊防护林土壤呼吸速率的日动态变化具有明显的季节特征。主要表现为以下3种情况:(1)2018年4—10月,3种城郊防护林的土壤呼吸具有明显的日变化规律,均呈现单峰型;其中4—6月、9—10月,土壤呼吸速率最大值均出现在 10:00—15:00,最小值均出现在20:00—5:00;7月与其他月份不同,其土壤呼吸速率虽然也呈现单峰型,但土壤呼吸速率最大值出现在18:00—20:00,最小值出现在02:00—04:00。这可能是由于夏季高温持续时间长,大气温度的峰值出现在15:00—16:00,但是土壤呼吸的最大值有时候会出现滞后现象,这主要因为此时土壤温度并未出现与气温具有一致的高峰值,而是滞后于气温,而土壤呼吸对土壤温度的响应速度比较快[11],因此土壤呼吸的最大值也出现滞后现象。(2)8月,刺槐和火炬树的土壤呼吸速率日变化趋势相一致,最大值均出现在 19:00—21:00;杨树较前两者有所不同,变化趋势较紊乱且无明显规律,但白天土壤呼吸速率明显高于夜间。在整个生长季土壤呼吸速率日变化表现为4月土壤呼吸速率日变化表现为火炬树>杨 树> 刺槐;5—8月土壤呼吸速率日变化表现为刺槐>火炬树>杨树;9月土壤呼吸速率日变化表现为火炬树>刺槐>杨树;10月土壤呼吸速率日变化表现为刺槐>杨树>火炬树,且日变化幅度均表现为白天大于夜间,这主要是因为白天温度较高,土壤中微生物和根系呼吸比较活跃所导致[12]。由显著性分析结果可知,3种城郊防护林土壤呼吸速率的日动态变化在生长季(4—10月)差异明显。土壤呼吸的日变化主要受土壤温度控制,而其他重要的环境变量,如土壤湿度、生物量和土壤性状等在一天内的变化相对较小,对土壤呼吸作用的影响不明显[13]。
2.2.2 3种城郊防护林土壤呼吸速率的季节变化
3种城郊防护林土壤呼吸速率存在着明显的季节变化规律,如图4所示。由于测量地点位于我国中纬度的温带地区,四季分明,生长季时间较长,为每年的4—10月,加之该地区受季风和海洋气候的影响,土壤呼吸速率的季节变化特征受温度、水分、地上植被的生长发育等多方面的影响。因此,从图4可以看出,3种城郊防护林的土壤呼吸速率在生长季内的变化均呈现单峰曲线,并有明显的季节变化规律。
4月处在冬春交替季节,气温偏低的同时T10也较低,导致3种植被根系生长缓慢且土壤中微生物活动不够活跃,因此土壤呼吸速率偏低;到春季开始的5—6月,气温逐渐上升,T10也随之增加,植被开始生长,根系和土壤微生物的活性增加,土壤呼吸速率出现明显上升的趋势;杨树防护林土壤呼吸速率在7月达到最大值,为(3.80±0.09) μmol/(m2·s);而火炬树和刺槐土壤呼吸速率则在8月达到最大,分别为(3.84±0.12)、(4.75±0.04) μmol/(m2·s)。这由于7—8月正值夏季,气候炎热,气温最高,光照较强,从而土壤温度受到影响,温度升高促使土壤微生物活性增强;且此时植物光合作用加强,从而影响到根系呼吸,这与其他学者的相关研究结果[14]相一致。到了秋季的9—10月,气温逐渐下降,植被生长缓慢,根系和土壤微生物的活性减弱,因此土壤呼吸速率逐渐下降。由显著性分析结果可知,3种城郊防护林土壤呼吸速率的季节变化差异不明显。
2.3 3种城郊防护林土壤温湿度对土壤呼吸速率的影响
3种城郊防护林土壤温湿度与土壤呼吸速率的相关关系如表2所示。国内外的学者利用线性方程、二次方程和指数方程等多种模型对土壤温度与土壤呼吸速率的关系进行拟合。本次研究结果中将T10和土壤呼吸速率之间的关系进行模型拟合并筛选出拟合度最佳的方程,即r2最大,P值最小的曲线模型。结果显示,3种城郊防护林土壤呼吸速率与T10呈顯著的二次曲线关系(P<0.05),其中,火炬树和刺槐与T10之间的相关程度最好,r2分别为0.847、0.844。而土壤呼吸速率与M5呈显著的线性正相关关系,其中,杨树和火炬树与M5的线性关系达到了极显著水平(P<0.01)。这说明对于3种城郊防护林而言,土壤含水量控制在一定范围内,土壤湿度会促进土壤呼吸速率,这与Birge等的研究结果[15]相吻合。
3种城郊防护林的土壤呼吸速率与土壤的温度及湿度均表现出显著的相关关系,但通常情况下,土壤温度与湿度会相互影响、相互作用于土壤呼吸[16],为进一步探究土壤温度和湿度对土壤呼吸速率的协同影响, 本研究对土壤呼吸速率与土壤温度及湿度的关系进行多元线性拟合,结果(表3)显示多元线性曲线能够很好地解释三者之间的相关关系,3种城郊防护林的土壤呼吸速率与土壤温度及湿度的相关程度非常显著,R2值分别为0.950、0.861、0.881,其中,火炬树和刺槐与T10和M5协同影响的线性关系达到了极显著水平(P<0.01)。
2.4 3种城郊防护林植被对土壤呼吸速率的影响的土壤呼吸速率呈现出明显的季节动态变化(图4),均表现为单峰曲线。杨树、火炬树和刺槐3种防护林土壤呼吸速率的变化范围分别为(0.86±0.15)~(3.80±0.09)、(1.43±0.02)~(3.84±0.12)、(1.05±0.03)~(4.75±0.04) μmol/(m2·s),平均值分别为(2.01±0.05)、(2.25±0.015)、(2.62±0.09) μmol/(m2·s),通过相关显著性分析发现,3种防护林植被的土壤呼吸速率均值差异明显,具体表现为刺槐>火炬树>杨树(图5),这说明除受到环境因子的影响外,土壤呼吸速率及强度同时还受到生物因子的影响,如细根生物量(表4),3种城郊防护林的土壤呼吸速率与细根生物量均呈显著的线性正相关关系(P<0.05),说明细根生物量越大,根系越发达,在生长季的根际呼吸越活跃,从而促进了土壤呼吸速率。
3 讨论与结论
3种城郊防护林所在的土壤温度和湿度均存在明显的季节变化规律。三者的土壤温度T10均呈现出明显的单峰曲线,并且在7月达到峰值;而土壤湿度M5的季节变化趋势则明显不同,4月开始,M5开始下降,到7月或8月达到峰值,随后又处于下降趋势。这是由于4月处于冬春交替季节,地面冰雪融化,雪水渗透到土壤,导致土壤湿度较高;到春季5—6月,降水较少,M5呈下降趋势;夏季是雨水集中期,7—8月降雨增多,M5明显上升,达到峰值;而后进入秋季,气候干燥、降雨减少,M5迅速下降。在本研究中,除8月外,3种城郊防护林的土壤呼吸在日变化尺度上均表现为单峰型,峰值出现在 10:00—15:00,这一时间段内,大气温度也处于高峰值,有研究表明土壤呼吸的日变化和气温、地表的温度呈显著相关性[17],因此在该时段内出现最大值。从整个生长季来看,3种城郊防护林的土壤呼吸速率基本表现为刺槐>火炬树>杨树,而在4月、9月、10月,三者的土壤呼吸速率变化趋势较为紊乱。但总体来看,刺槐的土壤呼吸速率最大,火炬树次之,杨树的最小。
3种城郊防护林土壤呼吸速率在各月的变化趋势基本一致,表现出明显的季节变化规律,均呈现单峰曲线,这与陈亮的研究结果[18]一致。在7—8月,各防护林的土壤呼吸速率均明显高于其他月份,这可能是受到季节性气候变化的影响,从而导致环境因子以及各生物因子的活性发生变化[19]。除此之外,3种城郊防护林土壤呼吸速率的季节变化特征还主要受到土壤温度与湿度的影响[20]。在植被生长初期的春季,气温较低的同时,土壤温度T10也较低,根系处于生长状态,细根生物量的含量较少,同时土壤微生物活性低,使根际呼吸以及土壤微生物的呼吸都很弱,从而导致土壤呼吸速率较低;随着气温和T10的升高,植被生长旺盛且土壤微生物活动十分活跃,导致根系和土壤微生物呼吸加强,从而使土壤呼吸速率迅速加强,达到峰值。
本研究表明,3种城郊防护林生长季的土壤呼吸速率均呈现出单峰曲线格局,具有明显的春秋低、夏季高的季节变化规律,并与土壤温度有着相同的季节变化规律,由此可以说明土壤温度是3种城郊防护林土壤呼吸速率的主要影响因素之一,通过相关显著性分析,土壤呼吸速率与T10有着显著的二次曲线关系,这与Gaumont-Guay等的研究结果[21-22]一致。有研究表明,土壤温度影响土壤呼吸速率主要是通过改变土壤微生物与植物呼吸酶的活性来实现的。土壤温度升高,在一定范围内会增强植物呼吸酶及土壤微生物的活性。同时,土壤湿度也是土壤呼吸速率的另一个主要影响因素。本研究中3种城郊防护林的土壤呼吸速率与M5呈显著的线性正相关关系。通常情况下,土壤温度与湿度会相互影响、相互作用于土壤呼吸,特别是在植被的生长季,两者的协同效应更为明显,因此本研究将T10、M5以及土壤呼吸速率三者进行多元线性拟合,发现其相关系数可以达到0.80以上,最低也可达到0.826,这说明多元线性模型能够更好地解释土壤呼吸与T10和M5之间的关系。
参考文献:
[1]Ryan M G,Law B E. Interpreting,measuring and modeling soil respiration[J]. Biogeochemistry,2005,73(1):3-27.
[2]Wu X G,Yuan J J,Ma S L,et al. Seasonal spatial pattern of soil respiration in a temperate urban forest in Beijing[J]. Urban Forestry & Urban Greening,2015,14(4):1122-1130.
[3]Wellock M L,Laoerle C M,Kiely G. What is the impact of afforestation on the carbon stocks of Irish mineral soils?[J]. Forest Ecology and Management,2011,262(8):1589-1596.
[4]Batjes N H. Mitigation of atmospheric CO2 concentrations by increased carbon sequestration in the soil[J]. Biology and Fertility of Soils,1998,27(3):230-235.
[5]Vesterdal L,Elberling B,Christiansen J R,et al. Soil respiration and rates of soil carbon turnover differ among six common European tree species[J]. Forest Ecology and Management,2012,264(15):185-196.
[6]張鸽香. 城市不同植被类型土壤呼吸与微生物量碳研究[D]. 南京:南京林业大学,2011.
[7]Raich J W,Tufekciogul A. Vegetation and soil respiration:correlations and controls[J]. Biogeochemistry,2000,48(1):71-90.
[8]张鸽香,徐 娇,王国兵,等. 城市3种类型人工林土壤的呼吸动态特征[J]. 南京林业大学学报(自然科学版),2011,35(3):43-48.
[9]Yue K,Peng Y,Peng C,et al. Stimulation of terrestrial ecosystem carbon storage by nitrogen addition:a meta-analysis[J]. Scientific Reports,2016,6:19895.
[10]Liu Y C,Liu S R,Wan S Q,et al. Differential responses of soil respiration to soil warming and experimental throughfall reduction in a transitional oak forest in central China[J]. Agricultural and Forest Meteorology,2016,226/227:186-198.
[11]秦 娟,上官周平. 白榆/刺槐不同林型生长季土壤呼吸速率的变化特征[J]. 西北农林科技大学学报(自然科学版),2012,40(6):91-98.
[12]Peng S S,Piao S L,Wang T,et al. Temperature sensitivity of soil respiration in different ecosystems in China[J]. Soil Biology & Biochemistry,2009,41(5):1008-1014.
[13]陶 晓,崔 珺,徐 俊,等. 城市绿地土壤呼吸特点及影响因素[J]. 土壤通报,2016,47(1):54-59.
[14]王春燕,陈秋波,袁 坤,等. 橡胶林土壤呼吸速率及其与土壤温湿度的关系[J]. 土壤学报,2013,50(5):974-982.
[15]Birge H E,Conant R T,Follett R F,et al. Soil respiration is not limited by reductions in microbial biomass during long-term soil incubations[J]. Soil Biology & Biochemistry,2015,81:304-310.
[16]Frank A B,Liebig M A,Hanson J D. Soil carbon dioxide fluxes in northern semiarid grasslands[J]. Soil Biology & Biochemistry,2002,34(9):1235-1241.
[17]刘绍辉,方精云,清田信. 北京山地温带森林的土壤呼吸[J]. 植物生态学报,1998,22(2):24-31.
[18]陈 亮. 亚热带地区3种典型森林生态系统碳通量特征研究[D]. 杭州:浙江农林大学,2017.[HJ2mm]
[19]Bae J,Ryu Y. Spatial and temporal variations in soil respiration among different land cover types under wet and dry years in an urban park[J]. Landscape and Urban Planning,2017,167:378-385.
[20]范洪旺,滕 臻,许克福. 城市绿地土壤呼吸研究综述[J]. 生态科学,2018,37(5):210-216.
[21]Gaumon-Guay D,Black T A,Griffis T J,et al. Influence of temperature and drought on seasonal and interannual variations of soil,bole and ecosystem respiration in a boreal aspen stand[J]. Agricultural and Forest Meteorology,2006,140(1/2/3/4):203-219.
[22]Luan J,Liu S,Wang J,et al. Tree species diversity promotes soil carbon stability by depressing the temperature sensitivity of soil respiration in temperate forests[J]. The Science of the Total Environment,2018,645(15):623-629.
收 稿日期:2019-02-22
基金項目:国家自然科学基金(编号:41303057);天津市应用基础与前沿技术研究计划项目(编号:14JCYBJC2300);天津市大学生创新创业训练计划项目(编号:201710060049)。
作者简介:张成芳(1995─),女,山东青岛人,硕士研究生,主要从事环境与城市生态研究。E-mail:15222713218@163.com。
通信作者:李德生,博士,教授,主要从事城市环境与城市生态教学及科研工作。E-mail:deshli@tjut.edu.cn。
