小菜蛾 PGRP-S2基因的克隆表达及功能分析
2020-05-11杜少萱苏月华
杜少萱,吴 程,苏月华,杨 梅
(福建师范大学生命科学学院,福州350100)
昆虫是世界上种类最多的一个群体,数量众多且分布广泛(Mayhewetal.,2007)。由于昆虫自身不存在获得性免疫系统,所以先天性免疫作为昆虫抵御病原微生物感染的重要防御系统显得尤为重要(Kimbrelletal.,2001;Choeetal.,2005)。肽聚糖识别蛋白PGRPs是昆虫模式识别受体中至关重要的一种病原识别蛋白,在昆虫的先天免疫反应过程中发挥着重要的作用。
PGRPs在结构上存在一个能与肽聚糖(peptidoglycan,PGN)结合的PGRP结构域,约由165个氨基酸构成,该结构域在无脊椎动物到脊椎动物中都具有高度的保守性。PGRP结构域与噬菌体T7的溶菌酶高度同源(Liepinshetal.,2003),噬菌体T7溶菌酶具有裂解PGN的酰胺酶活性,因此有些病原微生物能够被PGRPs直接杀死并清除(Werneretal.,2000;Liuetal.,2001;Dziarskietal.,2006)。根据肽聚糖识别蛋白分子量的大小,可以将已知昆虫PGRPs蛋白分为2种形式,即长型(L型)跟短型(S型),一般短型的PGRPs分子量大约在20~25 kDa,而长型的PGRPs分子量一般大于90 kDa(Guanetal.,2005;Dziarskietal.,2006)。研究表明大多数的S型 PGRPs主要在脂肪体内合成,而大多数L型PGRPs在血细胞中有着较高的表达量(Dimopoulosetal.,2002)。根据PGRPs有无酰胺酶活性又可将PGRPs分为有酰胺酶活性的PGRPs和无酰胺酶活性的PGRPs两种类型(Christophidesetal.,2002;Dziarskietal.,2003)。具有酰胺酶活性的PGRPs能断开细菌肽聚糖中肽桥-酰胺键,使肽聚糖裂解,而失去免疫活性,导致IMD通路信号下调(Zaidmanrémyetal.,2011)。有些PGRPs因缺少酰胺酶活性中所必须的位点,而无酰胺酶活性,使其不能裂解肽聚糖,但是这种类型的PGRPs仍然可以在免疫反应中作为受体发挥作用,譬如在果蝇Drosophilamelanogaster中发现的PGRP-SA蛋白和PGRP-SD蛋白虽然没有酰胺酶活性,却可以使作为受体激活Toll信号通路(Zaidmanrémyetal.,2006;Leoneetal.,2008)。随着基因组和转录组测序技术的发展,目前已在鳞翅目、鞘翅目、双翅目、膜翅目、直翅目、半翅目等多类昆虫中鉴定到大量的PGRP基因(Mellrothetal.,2003;Guanetal.,2005;Dziarskietal.,2006;Hashimotoetal.,2007;Tanakaetal.,2008;Werneretal.,2008;Sumathipalaetal.,2010;Khajuriaetal.,2011;Kangetal.,2014;Jieetal.,2015)。
小菜蛾Plutellaxylostella属于鳞翅目Lepidoptera菜蛾科Plutellidae,是一种主要危害白菜、萝卜等十字花科植物的世界性迁飞害虫,每年用于防控小菜蛾的费用高达40~50亿美元(Furlongetal.,2013)。近年来基于免疫系统进行生物防治的概念逐渐被提出,因此研究小菜蛾的免疫防御反应对未来进行生物防控具有重要的意义。肽聚糖识别蛋白是一种高度保守的病原识别蛋白,在小菜蛾抵抗病原微生物的过程中发挥着重要作用。本研究利用RT-PCR和RACE技术克隆了小菜蛾PGRP-S2基因,以原核表达的方式获得了重组蛋白,并探究了重组蛋白与大肠杆菌E.coli和金黄色葡萄球菌S.aureus之间的相互作用,用来了解小菜蛾PGRP-S2在细菌侵染过程中的潜在功能,为后续进一步深入研究小菜蛾的天然免疫系统提供基础。
1 材料与方法
1.1 小菜蛾
小菜蛾品系采集自福建省福州市闽侯县南通镇。
1.2 菌株和质粒
大肠杆菌E.coliDH5α和BL21(DE3)以及金黄色葡萄球菌S.aureus由本实验室保存;pMD19-T载体购自TaKaRa公司;pET28a载体由本实验室保存。
1.3 供试试剂
Eastep® Super总RNA提取试剂盒、Wizard® SV Gel and PCR Clean-Up System DNA 胶回收试剂盒、DNA限制性内切酶BamHI和EcoRI购自Promega公司;ExTaq酶、250 bp DNA Marker、RACE反转试剂盒SMARTer®RACE 5′/3′ Kit Components、T4 DNA 连接酶购自TaKaRa公司。
1.4 总RNA提取及cDNA合成
取成虫形态的小菜蛾样品5头,按Eastep® Super总RNA提取试剂盒提取总RNA。紫外微量分光光度计及1.0 %凝胶电泳检测RNA浓度及完整性。以提取的总RNA为模板,利用SMARTer® RACE 5′/3′ Kit Components试剂盒合成5′-cDNA与3′-cDNA,于-20℃保存。
1.5 5′-RACE与3′-RACE扩增
根据NCBI数据库中小菜蛾肽聚糖识别蛋白转录组序列(NCBI Reference Sequence:XM_011562450.1)的保守区域设计5′-RACE与3′-RACE特异性引物(表1)。以1.4节中反转录的cDNA为模板,采用ExTaq酶进行第1轮扩增,反应体系为:10 ×ExTaqbuffer 1.5 μL,dNTP Mixture 0.5 μL,UPM 0.5 μL,引物10.5 μL,5′-RACE cDNA/3′-RACE cDNA 1.0 μL,ExTaq酶0.15 μL,ddH2O 10.9 μL。PCR扩增条件为:94℃预变性5 min;94℃变性30 s,56℃退火30 s,72℃延伸2 min,共30个循环;72℃延伸10 min;4℃保存。以第1轮PCR产物稀释10倍后作为第2 轮PCR反应的模板,UPS和引物2为扩增引物,进行第2轮扩增,扩增条件同上。PCR产物经1%凝胶电泳检测正确后,回收纯化,并送至铂尚生物技术(上海)有限公司测序,测序结果用DNAStar进行拼接。

表1 PCR引物Table 1 Primers for PCR
1.6 小菜蛾 PGRP-S2基因生物信息学分析
利用ORFfinder(https://www.ncbi.nlm.nih.gov/orffinder/)分析PGRP-S2的开放阅读框。利用CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/docs/cdd_search.html)预测PGRP-S2的保守结构域;利用Protparam(https://web.expasy.org/protparam/)预测PGRP-S2蛋白的理化性质;利用Protscale(https://web.expasy.org/protscale/)预测 PGRP-S2蛋白的亲疏水性;利用SignalP 4.1(http://www.cbs.dtu.dk/services/SignalP/)预测 PGRP-S2蛋白是否存在信号肽;利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)预测 PGRP-S2蛋白是否存在跨膜结构;利用ClustalX 2.0和MEGA 5.0 (Xie HYetal.,2017)进行序列比对分析和进化树构建(NJ法,1000次重复)。
1.7 小菜蛾 PGRP-S2基因的原核表达与纯化
根据对序列的生物信息学分析结果,结合表达载体pET28a酶切位点信息,设计含有酶切位点的引物。在上游引物引入BamHI酶切位点,下游引物引入EcoRI酶切位点,引物见表1。PCR扩增获得目的基因并连接到pMD19-T载体上,用BamHI酶和EcoRI酶对重组质粒pMD19-T-PGRP-S2和pET28a质粒进行双酶切后用T4 DNA连接酶进行连接,以构建重组表达载体pET28a-PGRP-S2。将重组表达载体转化到感受态细胞BL21(DE3)中,以空载的pET28a作为对照,接种单克隆菌株到含100 mg/mL kana的5 mL LB液体培养基中,37℃、230 rpm过夜摇菌;次日按照1%的接种量接种于含100 mg/mL kana的50 mL LB液体培养基中,37℃、230 rpm发酵至OD为0.6~0.9;向发酵液中加入1 mol/L的IPTG 30 μL,使IPTG的终浓度为0.6 mmol/L,18℃、150 rpm诱导24 h使目的蛋白表达,以未加IPTG诱导作为对照组。发酵结束后离心收集菌体进行超声破碎,再次离心分别收集细胞破碎沉淀和上清液。12% SDS-PAGE分析蛋白表达结果。使用AKTA蛋白纯化仪对目的蛋白进行Ni柱亲和层析纯化,利用HiTrapTMDesating脱盐柱将蛋白纯化缓冲液更换为Tris缓冲液以除去咪唑。
1.8小菜蛾 PGRP-S2蛋白功能验证
取金黄色葡萄球菌(G+)与大肠杆菌BL21(DE3)(G-)划线活化,挑取活化后的单菌落过夜培养,次日按1%的接种量加到30 mL LB液体培养基中发酵至 OD600为0.5,取1 mL发酵液离心收集沉淀,用Tris缓冲液洗涤3次,最后用1 mL Tris缓冲液悬浮菌体,备用。
1.8.1重组蛋白的凝集试验
将供试菌悬液稀释至原浓度的10-5倍,在1.5 mL离心管中建立如下反应体系:

表2 反应体系Table 2 The reaction system
加样混匀,放置于摇床中37℃、150 rpm孵育3 h后,在倒置显微镜下观察菌体的分布情况。
1.8.2重组蛋白的抑菌试验
配制如表2所示的反应体系,对实验组中的PGRP蛋白液设置浓度梯度(0.1 mg/mL、0.5 mg/mL、1.0 mg/mL),放置于摇床中37℃、150 rpm孵育3 h后,取70 μL反应液均匀涂布于LB平板上,37℃过夜培养,次日观察菌落的生长情况,每组3次重复。
2 结果与分析
2.1 小菜蛾5′-RACE与3′-RACE扩增
经过两轮的PCR扩增,1.0%琼脂糖凝胶电泳,结果发现5′端和3′端各出现一条特异性条带,条带大小与预期结果相同(图1)。
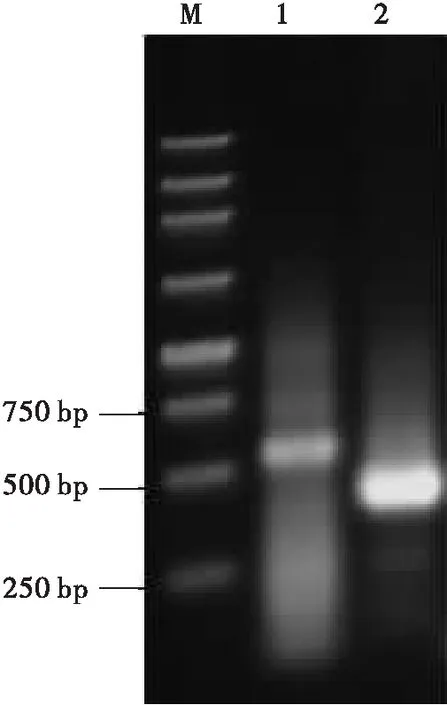
图1 RACE PCR 产物Fig.1 PCR product of RACE注:M,250 bp DNA 标准分子量;1,5′-RACE PCR产物;2,3′-RACE PCR产物。Note:M,250 bp DNA marker;1,PCR product of 5′-RACE;2,PCR product of 3′-RACE.
2.2 小菜蛾PGRP基因克隆及序列分析
对5′端和3′端的特异性条带进行回收测序,并用SeqMan进行序列拼接。结果成功克隆了小菜蛾PGRP基因(图2),命名为PGRP-S2(GenBank:MG570190),该序列的ORF全长为588 bp,编码195个氨基酸。CDD结构域预测显示该序列含有典型的PGRP超家族保守结构域跟酰胺酶结构域(图3)。Protparam分析结果表明其编码的蛋白质理论相对分子质量为21.46 kDa,理论等电点pI为8.46。该蛋白不稳定性参数为27.90,表明该蛋白属于稳定蛋白,平均亲水性(GRAVY)为-0.075,预测该蛋白质为水溶性蛋白质。Protscale分析结果表明该蛋白属于亲水性蛋白质,该结果与Protparam分析的结果一致。SignalP 4.1分析结果表明该蛋白N端存在一个由第1~21位氨基酸组成的信号肽。TMHMM预测该蛋白质不存在跨膜结构。
2.3 小菜蛾 PGRP-S2基因系统进化分析
将PGRP-S2编码的氨基酸序列与与已知功能的鳞翅目昆虫如棉铃虫和柞蚕等的PGRP蛋白的氨基酸序列进行多重比对(图4),结果显示,小菜蛾PxPGRP-S2中酰胺酶活性所必须的位点(52Q、161H、167T、169S,用*号表示)和大多数昆虫具有高度同源性。系统进化分析显示(图5),小菜蛾的PxPGRP-S2与家蚕的短(S)型肽聚糖识别蛋白BmPGRP-S1聚在同一分支上,两者的进化距离最近。

图2 PxPGRP-S2 基因序列及其编码产物Fig.2 The gene sequence of the PxPGRP-S2 and its deduced amino acid 注:单下划线表示信号肽序列;双下划线表示PGRP结构域。Note:The underlined signal indicates the signal peptide sequence;the double underline indicates the the PGRP domain.
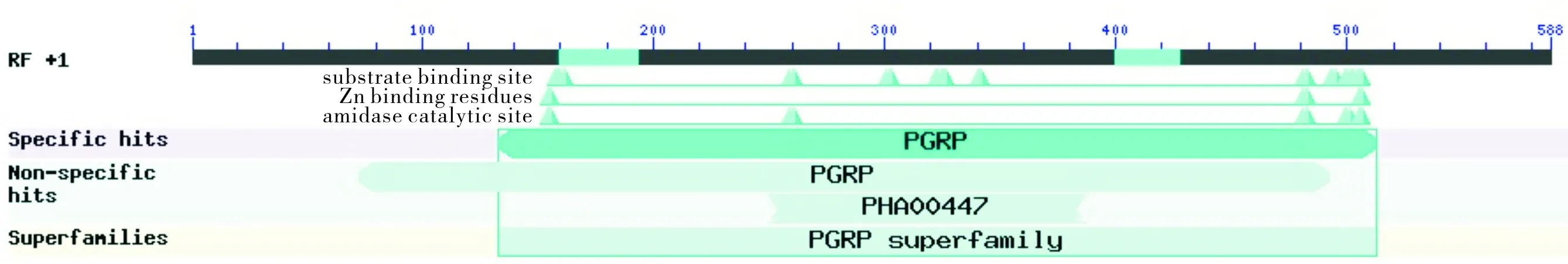
图3 克隆序列编码产物 PGRP-S2 的保守区域分析Fig.3 Analysis of conserved domains of the product PGRP-S2 encoded by the cloned sequence

图4 小菜蛾 PxPGRP-S2 氨基酸序列和其他昆虫PGRPs氨基酸序列比对Fig.4 Amino acid sequence alignment of PGRP-S2 from Plutella xylostlla with PGRPs from other insects注:*表示小菜蛾 PGRP-S2酰胺酶活性位点,PGRPs来源及GenBanak登录号如下:PxPGRPs,小菜蛾;ApPGRPs,柞蚕;BmPGRPs,家蚕;DmPGRPs,果蝇;HaPGRPs,棉铃虫;MsPGRPs,烟草天蛾;SlPGRPs,斜纹夜蛾。Note:* indicates the PxPGRP-S2 amidase active site of P.xylostella.The origin of PGRPs and their GenBank accession numbers were as follows:PxPGRPs,Plutella xylostella (PxPGRP-S2:MG570190);ApPGRPs,Antherea pernyi (ApPGRP-A:AME17978.1;ApPGRP-B:AME17979.1;ApPGRP-C:AME17980.2);BmPGRPs,Bombyx mori (BmPGRP-S1:NM_001043371.1;BmPGRP-S2:KF906541.1;BmPGRP-S5:NM_001043393.1);DmPGRPs,Drosophila melanogaster (DmPGRP-SC2:NP_610410.1);HaPGRPs,Helicoverpa armigera (HaPGRP-A:KF954940.1;HaPGRP-C:JX082167.1;HaPGRP-D:KF985962.1);MsPGRPs,Manduca sexta (MsPGRP-1A:AF413068.1;MsPGRP-1B:AF413061.1;MsPGRP-2:GQ293365.1);SlPGRPs,Spodopter litura (SlPGRP:XP_022825445.1)
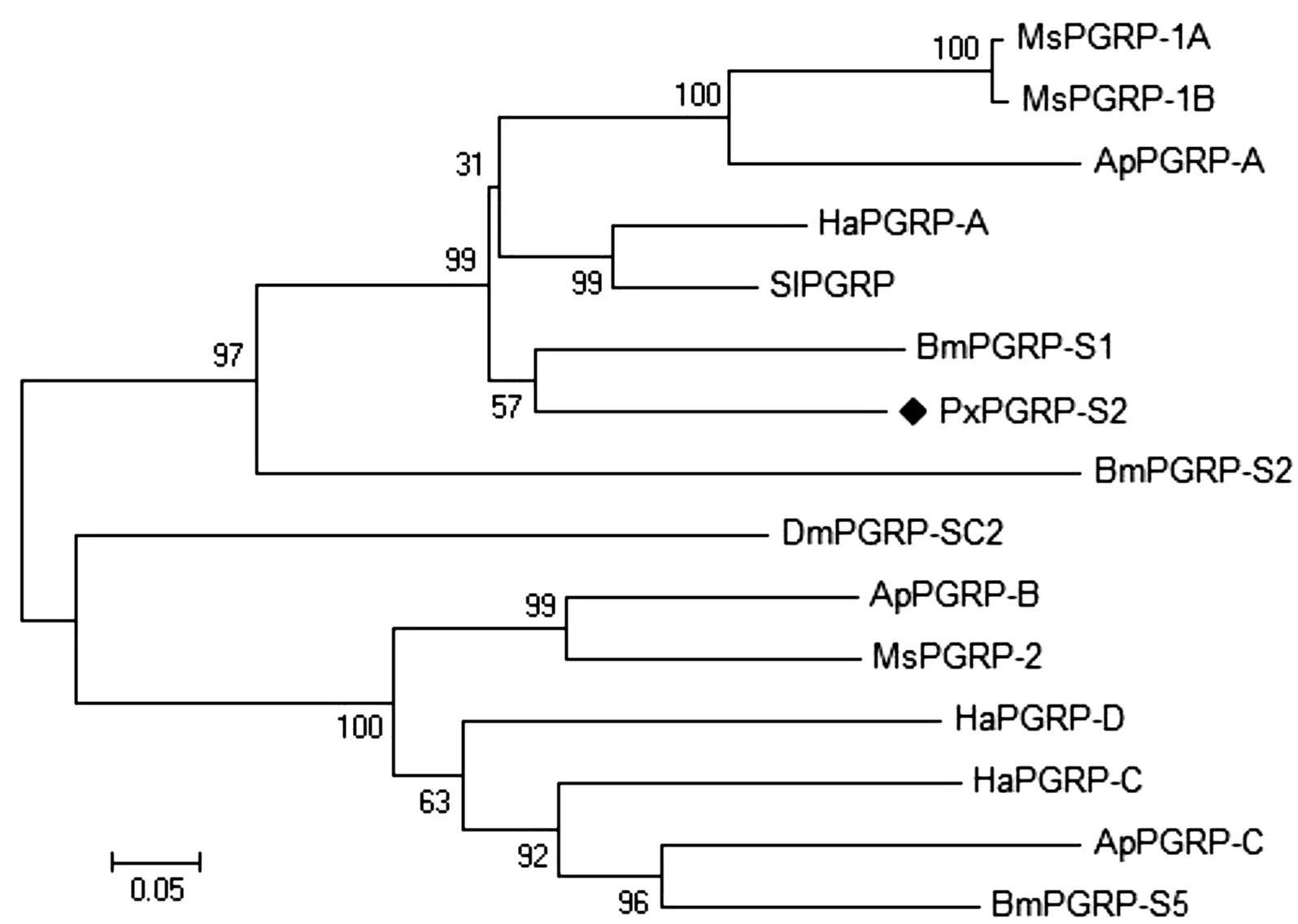
图5 小菜蛾 PxPGRP-S2与其它昆虫肽聚糖识别蛋白的系统发育树Fig.5 Phylogenetic tree of PxPGRP-S2 of Plutella xylostella and PGRPs from other different insect species注:PGRPs来源:同图3。Note:Origin species of PGRPs:The same to Fig.3.
2.4 小菜蛾PxPGRP-S2原核表达及纯化
重组工程菌BL21(DE3)-pET28a-PGRP-S2,经IPTG诱导后,高效表达可溶性蛋白(图6),诱导的上清液(图6第2泳道)相对于对照组(图6第1泳道)在约25 kDa处附近出现一条特异性条带,与预期的大小一致。使用AKTA蛋白纯化系统对目的蛋白进行Ni柱亲和层析纯化,洗脱得到目的蛋白(图6第6-9泳道)。

图6 SDS-PAGE 检测纯化的 PxPGRP-S2 蛋白Fig.6 SDS-PAGE analysis of purified PGRP-S2 protein purification注:M,蛋白质分子量标准品;1,未诱导对照组;2,破壁上清液;3,穿透液;4-9,20~500 mM咪唑浓度线性梯度洗脱蛋白。Note:M,Protein Marker;1,No induction control;2,Broken supernatant;3,Penetration;4-9,The protein was eluted with a gradient of 20~500 mM imidazole.
2.5 小菜蛾PxPGRP-S2蛋白功能验证
2.5.1重组蛋白的凝集试验
重组蛋白PxPGRP-S2和两种细菌的凝集试验结果如图7所示,在孵育3 h后,大肠杆菌对照组(图7 A)和金黄色葡萄球菌对照组(图7 C)中的菌体均以游离状态分布,未发生凝集现象;经过重组蛋白PxPGRP-S2处理后的大肠杆菌(图7 B)和金黄色葡萄球菌(图7 D)与对照组相比发生了凝集(图7 B,图7 D中箭头标注的位置),提示PxPGRP-S2蛋白对细菌具有凝集作用。
2.5.2重组蛋白的抑菌试验
利用重组蛋白PxPGRP-S2对大肠杆菌和金黄色葡萄球菌进行抑菌效应分析,结果发现对照组单菌落均匀分布在培养基上面,没有连成块状,而实验组因加入重组蛋白PxPGRP-S2后,菌体之间相互凝集在一起,在培养基上形成团状,且随着重组蛋白浓度的增加,形成的团状越大,表明随着重组蛋白浓度的增加,更多的菌体凝集在一起。但从对照组和实验组的菌落数量上观察,重组蛋白PxPGRP-S2并没有起到抑菌效果,不具有统计学意义。表明PxPGRP-S2蛋白对于这2种菌只有凝集作用而无杀菌功能。
3 结论与讨论
在果蝇PGRPs的研究中,目前从果蝇基因组中鉴定出来的13个肽聚糖识别蛋白,其中有7个L型的肽聚糖识别蛋白含有跨膜结构域和信号肽,它们被确认为跨膜蛋白(如PGRP-LC);而多数S型的肽聚糖识别蛋白,在它们的氨基酸序列中只有信号肽而不存在跨膜域,它们可能作为分泌蛋白(Lemaitreetal.,2007)。对小菜蛾PGRP-S2序列进行信号肽以及跨膜分析表明该序列编码的蛋白存在信号肽结构但不存在跨膜结构,推测该序列编码的蛋白属于分泌型蛋白的可能性较大。
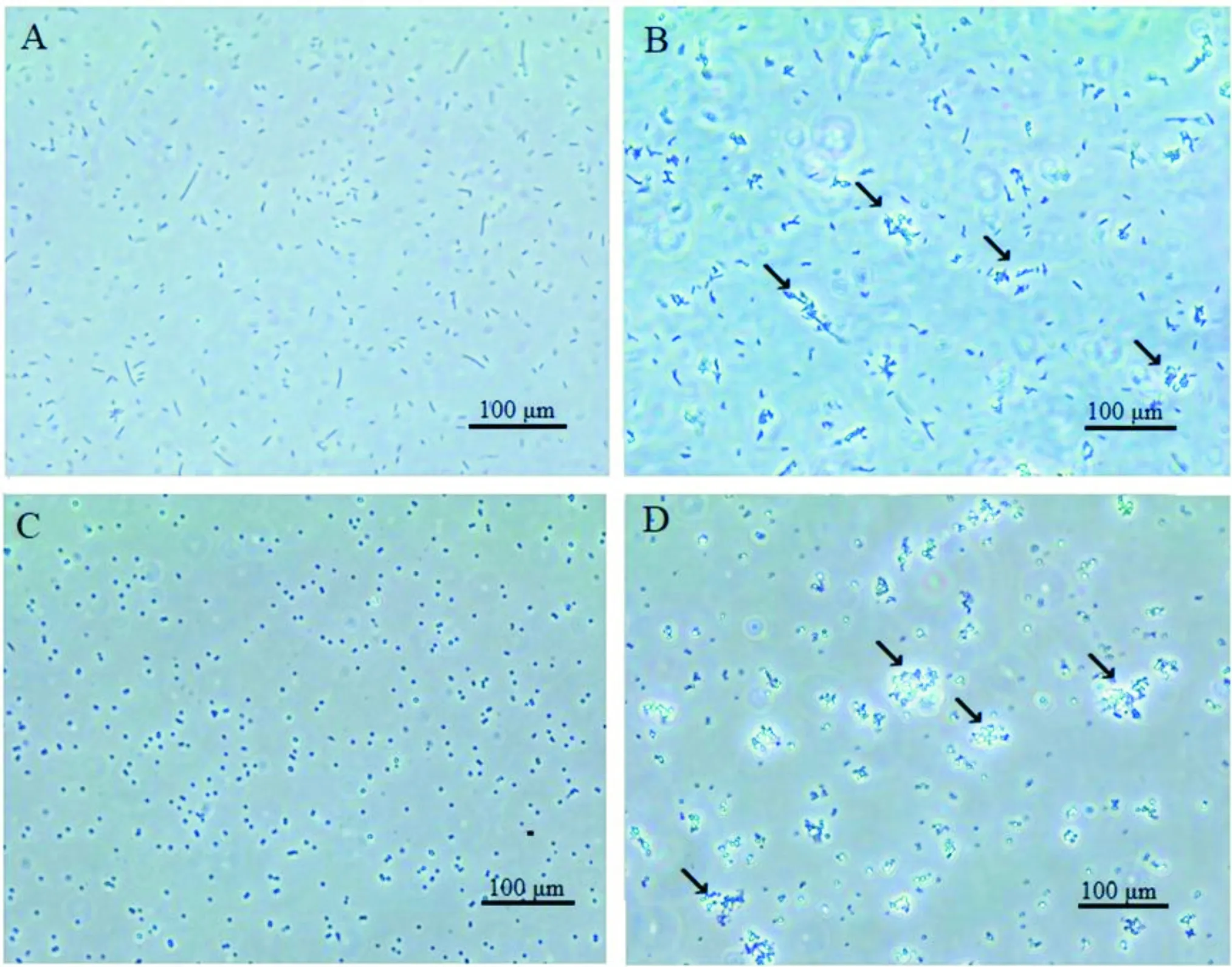
图7 重组蛋白PxPGRP-S2对两种细菌的凝集反应Fig.7 The agglutination of PxPGRP-S2 reconbinant protein with two types of bacteriaA,Escherichia coli/Tris(ZnCl2);B,E.coli/PxPGRP-S2(ZnCl2);C,S.aureus/Tris(ZnCl2);D,S.aureus/PxPGRP-S2(ZnCl2)
基于前期对PGRPs功能的研究发现不管是无脊椎动物还是脊椎动物,能够使细菌肽聚糖裂解的PGRP蛋白很可能具有抗菌活性,比如在果蝇中的PGRP-SB1蛋白(Geliusetal.,2003),以及人类、小鼠还有文昌鱼中的PGRP-L蛋白(Wangetal.,2003;Yangetal.,2010;Fengetal.,2012;Lietal.,2012),这些蛋白在免疫过程中直接作为杀菌因子。果蝇PGRP-SB1蛋白因其具有保守的酰胺酶结构而具有抗菌活性,但并不是所有具有酰胺酶活性结构域的PGRP蛋白都具有杀菌活性,比如果蝇PGRP-SC蛋白也具有保守的酰胺酶结构,但该蛋白就没有直接的抗菌活性(Mellrothetal.,2006),同时罗嫚等人(2016)对家蝇PGRP-SA蛋白的研究发现,该蛋白能够与大肠杆菌和金黄色葡萄球菌结合,但是并不能直接杀死它们,而是通过与它们结合并识别,从而激活Toll信号通路,引起下游抗菌肽的表达。
本研究发现小菜蛾PxPGRP-S2蛋白能够介导大肠杆菌和金黄色葡萄球菌发生凝集反应,但并不具备直接的抗菌活性,表明该蛋白虽然具有酰胺酶结构域,但不具备酰胺酶活性,其作用机理可能是通过识别并结合细菌表面的肽聚糖,激活下游信号通路产生抗菌肽等杀菌物质,实现对病原微生物的免疫防御。当然,对于该蛋白具体功能和作用机制的研究还有很长的路要走,比如该蛋白在小菜蛾体内的功能是什么?主要响应的是哪一类病原微生物?其介导的下游免疫信号通路是哪些?具体的机制如何?这些问题均需要在后续的研究中进一步深入探讨。本研究为深入研究该蛋白的体内功能提供了必不可少的序列信息、重组蛋白以及初步的功能预测。
