濒危植物驼峰藤的扦插繁殖及野外回归*
2020-04-16熊武建孙红梅刘盼盼冯璐关开朗廖文波由文辉
熊武建,孙红梅,刘盼盼,冯璐,关开朗,廖文波,由文辉
(1. 有害生物控制与资源利用国家重点实验室/广东省热带亚热带植物资源重点实验室/中山大学生命科学学院, 广东 广州 510275;2. 中山市国有森林资源保护中心,广东 中山528400;3. 上海市城市化生态过程与生态恢复重点实验室/华东师范大学生态与环境科学学院,上海 200241)
在传统的植物分类系统中,驼峰藤Vincetoxicumhainanense(synonymMerrillanthushainanensis) 隶属于萝藦科Asclepiadaceae、驼峰藤属Merrillanthus[1-2],但按基于分子系统发育建立的APG Ⅳ被子植物分类系统及最新的分类学研究,驼峰藤则被归并到夹竹桃科Apocynaceae[3]、白前属Vincetoxicum[4]。驼峰藤是国家Ⅱ级保护植物和第1批国家重点保护野生植物[5],具有重要的生态学价值。随着世界人口的迅速膨胀和经济活动的不断加剧,大面积森林被砍伐,生态系统正在受到严重的破坏,适宜驼峰藤生存的生境不断萎缩和消失。植物资源调查表明驼峰藤分布区域狭窄,仅在海南保亭、万宁、白沙及广东高要等地有过标本采集记录[6-8]。2007年中山市野生植物调查首次发现在中山田心伯公坳有驼峰藤野生分布[9]。2015年6月,在中山市田心森林公园再次发现驼峰藤的踪迹,林业部门立即对驼峰藤进行科学保护并开展科研试验。
目前关于驼峰藤的研究甚少,仅有侯俊等[10]对驼峰藤的组织培养及植株再生进行了相关研究,廖浩斌等[11]对驼峰藤的物候与繁育特征进行了研究,熊武建等[12]对驼峰藤进行了叶绿体基因组测序研究,而关于其扦插繁殖和野外回归等方面尚未有研究报告,因此探索驼峰藤扦插繁殖和野外回归的有效途径,为驼峰藤种植资源的有效保育和种群恢复提供理论依据,具有重要的科学价值和实践价值。回归(reintroduction)是基于迁地保护的基础,通过人工繁殖把植物引种到原来分布的自然或半自然的生境中,以建立具有足够的遗传资源来适应进化改变、可自然维持和更新新种群[13]。作为物种保护及种群恢复的重要策略之一,回归在越来越多的珍稀濒危植物保护实践中得到了应用[14-15]。
1 材料与方法
1.1 研究地点和材料
研究区位于中山市田心森林公园(22°24′39″N,113°28′21″E)。该地区属南亚热带海洋季风气候,年日照为1 694.4 h,光照充足,四季分布比较均匀,年平均气温20.3 ℃。年平均降雨量1 858.2 mm,雨量充足,水热丰富,地势大致呈东北高西南低,平地较少、山地较多,最高峰海拔为383.7 m。土壤以沙壤土和红壤土为主。植被群落组成以驼峰藤为特征种,主要伴生种有艳山姜Alpiniazerumbet、地桃花Urenalobata、土沉香Aquilariasinensis、粉单竹Bambusachungii、粗叶榕Ficushirta、海芋Alocasiamacrorrhiza、猴耳环Pithecellobiumclypearia、假苹婆Sterculialanceolata等。
选取生长茁壮的驼峰藤植株,剪取无病虫害的枝条带回中山大学引种栽培温室内进行扦插繁殖。如图1所示,将枝条分为四个等级,分别剪下一、二、三级枝条(其中一级枝条枝茎约0.3 cm、二级枝条枝茎约0.5 cm、三级枝条枝茎约0.7 cm),将每级枝条剪成长约12 cm的小段,每段保留2个节。截取插穗时,上端距节1 cm处平切并保留一片叶子,下端距节约1 cm处斜切,于2018年4月10日进行插穗扦插实验。
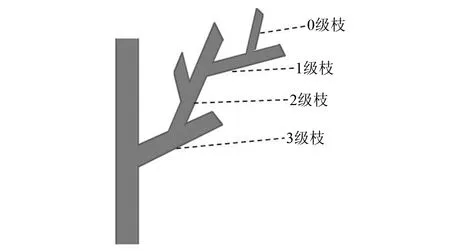
图1 枝条分级定义模式图
1.2 扦插繁殖正交试验
根据驼峰藤的适生环境及栽培技术要点,从影响驼峰藤扦插生根成活的插穗水平、扦插基质和生长激素等因素中,以生物统计的正交试验设计为理论依据,列出影响扦插生根成苗率的主要因子,进行3因素3水平正交试验(表1) ,共9个处理,每个处理30个插穗,3次重复。本试验以萘乙酸(NAA)作为生长素,共设2个不同浓度,以不加任何激素作对照,将驼峰藤插穗剪好后浸入0.25 g/L 的多菌灵溶液中进行消毒,浸泡时间15 min,后取出晾干,再将插穗基部浸入NAA激素液2~3 cm,浸泡10 min后取出晾干,然后进行扦插,从扦插开始7~10 d后开始观测最早生根和不定芽萌发时间,一个月后统计成苗率。待插穗生根成苗后进行复壮栽培,直至植株生长健壮并具有8~10片成熟叶片。
1.3 野外回归对比实验
2018年6月3日起苗保湿处理包装,于2018年6月4日定植于自然分布地进行野外回归对比实验。结合野外调查结果和驼峰藤的生长习性,野外回归选择中山市田心森林公园内3个具有代表性的生态群落,即原生森林生态群落:植被群落以艳山姜Alpiniazerumbet、鸭跖草Commelinacommunis、地桃花Urenalobata、土沉香Aquilariasinensis、粉单竹Bambusachungii、粗叶榕Ficushirta、海芋Alocasiamacrorrhiza、猴耳环Pithecellobiumclypearia、假苹婆Sterculialanceolata、鲫鱼胆Maesaperlarius和娃儿藤Tylophoraovata等植物群落组成;次生开发阴香林群落:植被群落由阴香Cinnamomumburmanni、樟树Cinnamomumbodinieri、木油桐Verniciamontana、小叶榕Ficusconcinna、锡叶藤Tetraceraasiatica、华山姜Alpiniachinensis、鬼针草Bidenspilosa、九节Psychotriarubra、五爪金龙Ipomoeacairica等植物群落组成;次生开发蒲桃林群落:植被群落由蒲桃Syzygiumjambos、牛耳枫Daphniphyllumcalycinum、华山姜Alpiniachinensis、鹅掌柴Scheffleraoctophylla、水东哥Saurauiatristyla、蟛蜞菊Wedeliachinensis、葛Puerarialobata、华南毛蕨Cyclosorusparasiticus、麦冬Ophiopogonjaponicus等植物群落组成。在进行回归引种对比试验时,每个群落定植100株回归试验苗,共300株。分别于2018年7月4日(定植1个月),2018年12月4日(定植6个月),2019年3月4日(定植9个月)对其成活及生长状况进行统计调查。回归引种结果调查计算:



生长状况记录株高,基径,主茎分枝数等。
1.4 数据处理
试验数据均采用SPSS 20.0进行单因素方差(ANOVA) 分析,并用Duncan(α=0.05)分析法对数据进行多重比较。统计分析和制图通过 EXCEL 实现。
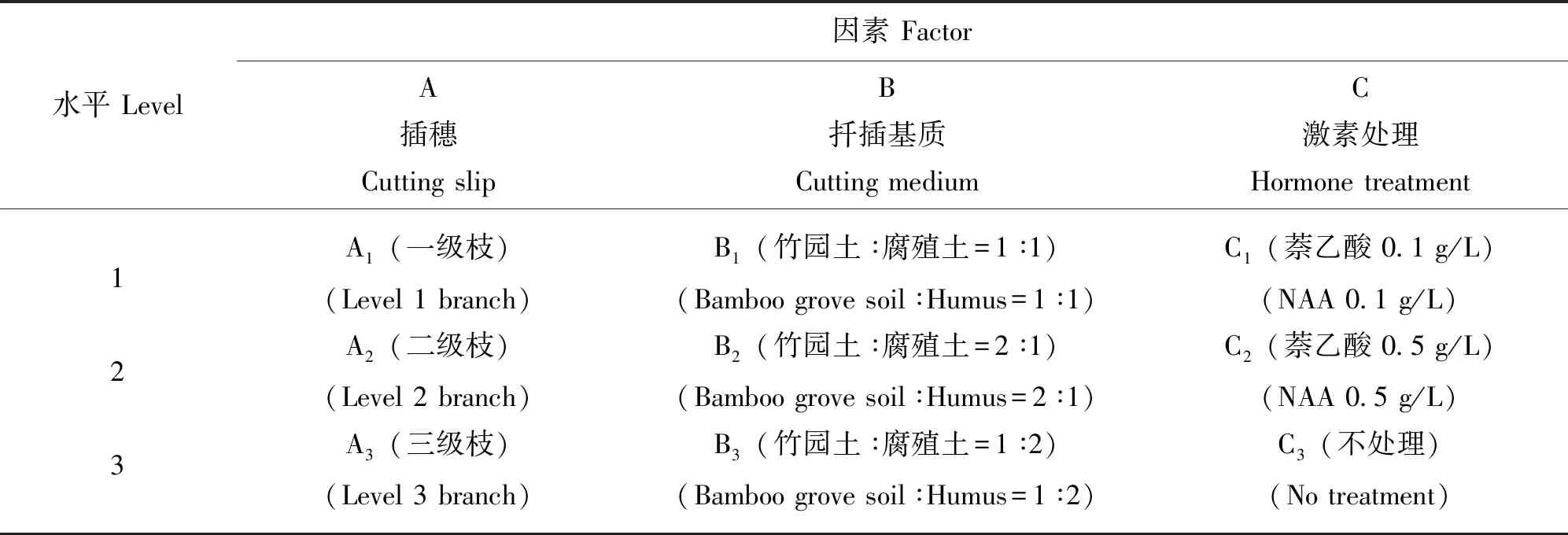
表1 正交试验因素水平表
2 结果与分析
2.1 扦插繁殖正交试验结果与分析
正交试验结果表明(表2,图2),A 因素中K3值等于 217最大, 即插穗因素中的最优水平是 3 水平(枝茎0.7 cm),B、C因素中的最大K值依次为K3= 154、K2=187, 即扦插基质、激素处理两个因素中的最优水平依次为 3 水平(竹园土∶腐殖土=1∶2)、2 水平(萘乙酸 0.5 g/L), 最佳因素水平搭配是A3B3C2。因此,采用三级枝条作为插穗、以竹园土∶腐殖土=1∶2作为扦插基质、0.5 g/L 萘乙酸处理切口是驼峰藤扦插生根成苗的最佳方案。
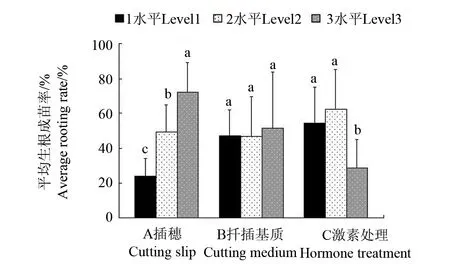
图2 各因素水平平均生根成苗率

表2 L9( 33) 正交试验及其统计分析结果1)
1)生根成苗率(K)=(生根成苗株数/插穗个体数)×100%;K1、K2、K3为各列中1、2、3水平的生根成苗率合计;RK表示全距(K1平均值、K2平均值、K3平均值中最大值减最小值)
1) Planting percent (K) =(Total number of rooted cuttings / Total number of cuttings)×100%;K1was total of rooting rate in each column of Level 1 and so forthK2,K3;RK= Range ( Difference between the maximum and the minimum betweenK1,K2andK3average values)
2.2 野外回归对比试验结果与分析
回归引种的对比试验结果表明(表3),原生森林生态居群回归引种植株定植1个月后的成活率是100.0%,定植6个月后的植株成活率96.7%,株高252.0 cm,基径0.78 cm,主茎分枝数3.7,定植9个月后的植株开花率45.5%,结实率32.7%;次生开发阴香林生态居群相应的成活率是97.8%,定植6个月后的植株成活率87.8%,株高189.6 cm,基径0.63 cm,主茎分枝数1.9,定植9个月后的植株开花率5.0%,结实率1.7%;次生开发蒲桃林生态居群相应的成活率是100.0%,定植6个月后的植株成活率86.7%,株高 163.6 cm,基径0.59 cm,主茎分枝数1.9,无开花结实植株。差异显著性分析的结果表明原生森林生态居群的回归引种成活率及生长发育明显优于次生开发阴香林和次生开发蒲桃林生态居群。也进一步明确植被稀疏、郁闭度小的原生森林生态环境有利于驼峰藤的生长和保存,是驼峰藤的适生生境;而植被茂密、郁闭度较大、光照略弱的次生开发阴香林生态群落及次生开发蒲桃林生态群落不利于驼峰藤的生长发育和自然繁衍。驼峰藤插穗扦插成苗及在原生森林生境下回归引种植株的生长过程如图3所示。
1) 居群1: 次生开发阴香林居群; 居群2: 次生开发蒲桃林居群; 居群3: 原生森林生态居群。不同字母表示显著差异(P<0.05)
1)Population 1: Ecological population of secondary forest ofCinnamomumburmanni; Population 2: Ecological population of secondary forest ofSyzygiumjambos; Population 3: Ecological population of primeval forest. Different letters indicate significant differences (P<0.05)

图3 驼峰藤扦插成苗及原生境下回归引种的植株
3 结论与讨论
驼峰藤扦插过程中剪取枝条的等级和激素浸泡浓度对扦插成活存在显著的影响,而扦插基质的配比对扦插成活也有影响,但未达显著水平。驼峰藤扦插繁殖的最佳方案为剪取三级枝条作为插穗,保留一叶片和两个节,以竹园土∶腐殖土=1∶2作为扦插基质,以0.5 g/L的萘乙酸(NAA)处理切口进行扦插繁殖。近年来,许多学者对影响濒危植物繁殖成活率的插穗规格、外源激素和扦插时间等因素进行了大量研究[16-18],不同学者所得出的研究结果不尽相同。本研究将扦插繁殖获得的驼峰藤再生植株回归自然,定期对其进行监测,调查统计其成活率和生长势,结果表明,回归 6 个月后,部分植株因受到自然环境的淘汰,有叶发黄、枝枯死等现象,回归过程中其成活率随着时间的延长而降低,但成活的驼峰藤植株生长良好,基茎变大,每个植株主茎长出1~4个分枝。其中原生森林生态居群回归种植的植株明显优于次生开发阴香林和次生开发蒲桃林生态居群,9个月后即可开花结实,植株开花率达45.5%,结实率达32.7%,因此回归引种必须以植物群落及生态环境为基础。有研究表明,特定的植物群落组成及其形成的特殊生态环境的变化会影响植物的生长率和开花[19-22]。此方案的获得为驼峰藤的迁地保护和回归引种等就地保护击开了一个突破点,可有效地保护和保存驼峰藤的遗传种质资源。但是,回归植株的种群繁衍、管理监测及遗传性状稳定性等需要继续深入研究。
珍稀濒危植物因物种进化史、所处生态环境、物种本身种群结构及遗传特性等而不能适应人类的干扰和生态环境的迅速变化[23-24]。开展珍稀濒危植物的野外回归,是一项困难的工作,需要考虑一系列科学和社会经济问题,成功的案列并不多[25]。珍稀濒危植物保护的主要方式是就地保护、迁地保护和野外回归。开展珍稀濒危植物的繁育技术攻关和基础研究,再进行市场化生产和野外回归,具有重要的科学和实践价值,可实现对濒危植物的有效保护。基于本研究的试验结果和驼峰藤的濒危现状,提出以下保育建议:1)为了提高自然回归成活率增强植株生长势,建议选择与原生地相似的适宜生境,在回归初期,加强监测管理,为植物的顺利生长提供充足的养分和空间;2)植物是生物多样性的重要组成部分,特别是珍稀濒危植物,在对其保护和回归时,需要综合考虑社会、经济、生态等问题,把植物回归与自然生态、人类社会等相联系,以产生良好的社会、生态和经济效益。
