小鼠胚胎早期发育过程中OCT4作用的研究进展
2020-03-30钟蓓高晗
钟蓓 高晗
摘要:OCT4属于 POU 转录因子。目前对于 OCT4 基因的研究认为在鼠和人体内,OCT4 主要表达于胚胎干细胞及生殖细胞中,对于维持胚胎干细胞的多能性和自我更新有十分重要的作用,对于胚胎早期发育具有重大意义。但是目前 OCT4 在小鼠早期胚胎中的结合位点还不清晰,应进一步深入研究 OCT4 基因构建 OCT4 在小鼠早期胚胎发育过程中的调控网络,为揭开胚胎早期发育调控机制的面纱提供重要参考。
关键词:小鼠;OCT4;早期胚胎发育;胚胎干细胞;调控网络
中图分类号: Q813文献标识码:B文章编号:2095-9737(2020)02-0007-03
小鼠是解析人类基因功能最重要的模式生物,是研究人类疾病发病机制的理想模型。小鼠和人类有类似的生理解剖构造和胚胎发育过程,染色体组成和人类的类似:小鼠有 19条常染色体(人类有22条),还有X和Y性染色体,而且小鼠许多基础生物学以及行为学过程与人类类似,同时对其基因组序列的分析表明,99%的人类编码基因都可以在小鼠基因组中找到对应的同源基因,同时染色体上基因之间的排列关系也在小鼠和人类基因组中高度保守[1]。这些数据为运用小鼠基因敲除技术和转基因技术研究人类基因功能提供了重要的理论基础。
OCT4是指位于人类基因组第 6 号染色体上的POU5F1 基因编码的OCT4A 亚型。它属于POU家族蛋白Class 5变体1,因此其基因名被称为POU5F1。OCT4是POU家族转录因子的一个八进制结合子群,它通过一个称为POU域的二部DNA结合域与八聚体motif(consensus sequence ATGCAAAT)结合[2]。POU 域由两个螺旋-转角-螺旋 DNA 结合域组成,通过可变连接区域连接。在 POU 家族中,POU 特异性结构域(POUS)和 POU 同源结构域(POUHD) 都是保守的,POU 家族蛋白可以與被调控基因启动子或增强子中的顺式作用元件结合发挥转录调控作用。1989 年,由 Schler 等人首次在小鼠原始生殖细胞、卵母细胞和体外培养的胚胎干细胞中发现[3-4]。OCT4 的早期研究是描述性的,并着眼于确定 OCT4 在胚胎以及胚胎来源的多能细胞系中的功能作用。
1 OCT4 基因在胚胎发育中作用的研究进展
OCT4 转录因子最初作为卵母细胞中的母体因子具有活性,并且整个植入前期在胚胎中保持活性。OCT4 表达与未分化的表型和肿瘤相关。OCT4 被发现是一种特异于早期胚胎发生的转录因子,它是小鼠胚胎植入前发育过程中的关键调控因子,它在 20 多年前的三项独立研究中被鉴定出来,这些研究使用不同胚胎阶段的核提取物和 ECC 系的核提取物来鉴定在胚胎早期发育中具有特定功能的转录因子。利用其与八聚体基序的特异性结合能力,OCT4 在不同的 ECC 系、未受精卵母细胞、早期胚胎和原始生殖细胞(PGCs)中均有表达,但在成体组织中未见表达。因此,OCT4 是一种转录因子,在早期胚胎发生中起重要作用。其表达仅限于发育后期的生殖系[3-5]。
在小鼠中,成熟卵母细胞中存在 OCT4 mRNA。在八细胞期,OCT4 的表达达到一个更高的水平。随后,OCT4 的表达被限制在内细胞团(ICM)中。在胚胎发育后期,OCT4 的表达仅维持在原始生殖细胞(PGCs)中[5]。
研究者检测了人胚泡中 PGCs 的表达。分离17例人胚泡的 ICM 和 TE,分别用RT-PCR[6-7]检测 OCT4mRNA 水平。结果表明,全能 ICM 细胞中 OCT4 的平均表达量是分化的TE细胞的30倍。这些研究表明,OCT4 在小鼠和人类细胞中的表达模式非常相似。
在小鼠中,POU5F1 基因与 17 号染色体上的主要组织相容性复合体(MHC)非常接近[8]。准确的位置被映射在 MHC 的 Q 和 T 区域之间。人类 POU5F1 基因定位于 6 号染色体,与小鼠POU5F1 有 87%的序列相同,显示出高度的进化保守性。这两个基因都由 5 个外显子和 4 个内含子组成。在小鼠中,POU5F1 基因转录从一个不包含 TATA-box 序列的近端启动子的GC富集区域的几个位点启动。启动子包括由 RA 诱导的孤核受体靶向的反应元件,以及特异性蛋白 1 (SP1)的结合位点,因此被认为在 RA 诱导的分化过程中发挥作用。即 OCT4 的表达是通过 POU5F1 基因内的多个调控元件来调控的。
OCT4 在早期细胞的命运决定中发挥作用。其中两个最具启发性的例子是:OCT4与CDX2在囊胚中的表达相互负调控,OCT4对FGF4表达的调控。CDX2是调节滋养外胚层谱系的关键转录因子,其在ESCs中的过表达通过抑制OCT4表达诱导滋养外胚层谱系分化。
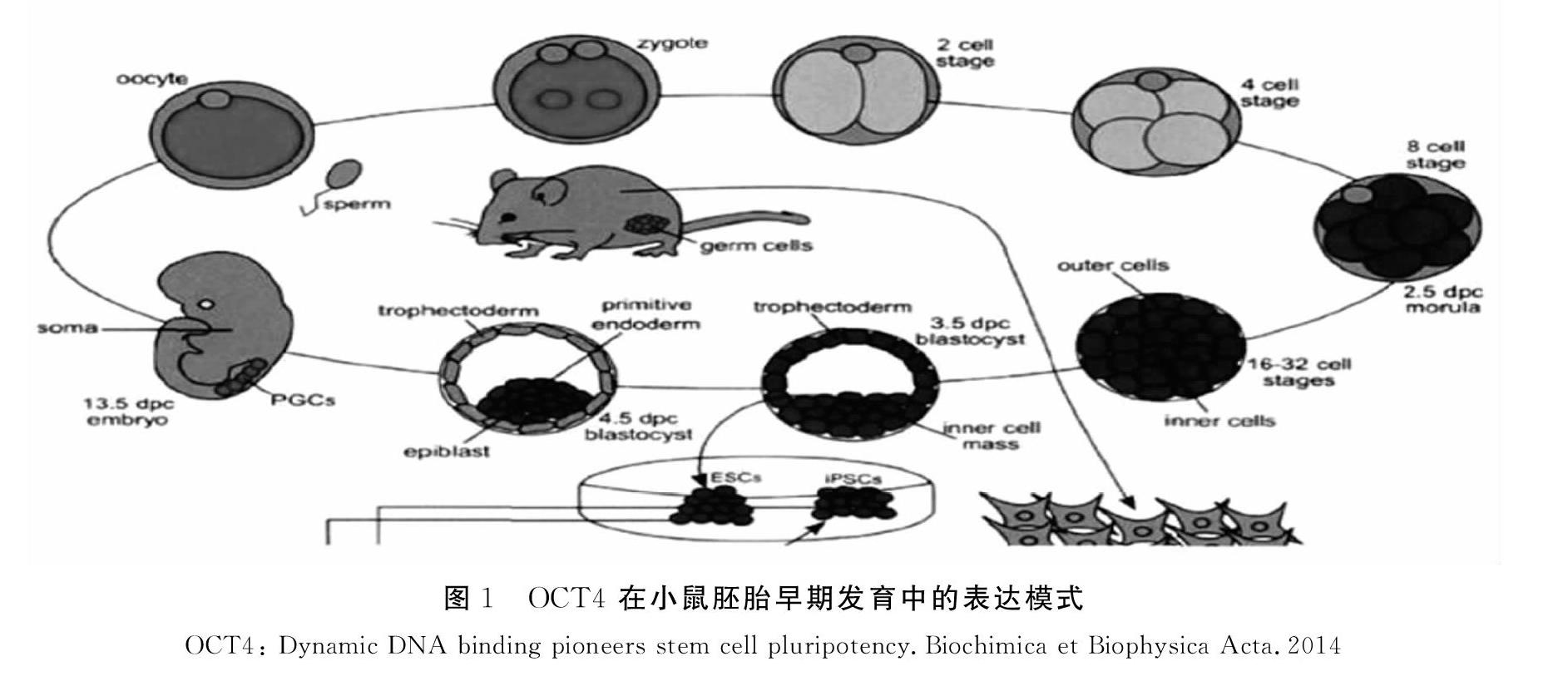
研究表明,小鼠卵母细胞中存在母源OCT4,随着受精和合子基因激活合子来源的OCT4才开始表达于4~8细胞期胚胎,并均匀的分布于早期胚胎各卵裂球细胞内直至在囊胚时期其表达开始局限于ICM,而从TE细胞中消失。对于着床后的卵圆柱期胚,我们只能在其原始外胚层中检测到OCT4的表达,在之后7~8天的胚胎中,能够在神经外胚层中检测到OCT4的表达。自8.5天起,就只能在原始生殖细胞中检测到其表达。因此,OCT4在原肠胚形成时期中对谱系定型起重要作用。在目前已有的研究中,研究人员们完成了对 OCT4 在小鼠胚胎早期发育中的表达模式的确定,见图1。
2 OCT4 基因在胚胎干细胞中作用的研究进展
已有的研究表明,OCT4是将体细胞重新编程为诱导多能干细胞所需要的四个重要的转录因子之一,是该过程中唯一不能被同一蛋白家族其他成员替代的因子[9]。OCT4可以与Sox2形成异二聚体,因此这两种蛋白质将DNA结合在一起。人OCT4基因通过可变剪切可以翻译成三种蛋白亚型,分别被命名为OCT4A、OCT4B和OCT4B1。OCT4A是最早被发现和研究的亚型,通常没有特别强调不同亚型的情况下提到的OCT4就是指OCT4A。OCT4A和OCT4B的DNA结合结构域和C-末端反式激活结构域相同,二者的差异主要是N-末端反式激活结构域外显子不同。OCT4B1和OCT4B使用相同的外显子,只是OCT4B1比OCT4B多了一个隐藏在OCT4A和OCT4B内含子序列中的外显子。OCT4A通常在早期胚胎细胞和胚胎干细胞等多能性细胞中表达,定位在细胞核中,与多能性调控密切相关。由于ECC与胚胎干细胞(胚胎干细胞是由内细胞团衍生而来的细胞)的相似性,ECC被广泛用作研究胚胎发育的模型。研究表明ECCs中OCT4表达受视黄酸(RA)诱导分化抑制。其在ESCs中的表达水平受严格的调控机制控制,它也是胚胎干细胞核心转录调控网络的一部分。通过Kazutoshi Takahashi 和 Shinya Yamanaka[10]的实验表明,OCT4 是诱导体细胞多能性(重编程)所需的四个转录因子中最小的一个,与之前描述的 OCT4 在 ESCs 中维持多能性的作用相比,OCT4 在诱导和建立诱导多能干细胞多能状态的背景下的研究也极为重要。目前,许多研究将重点放在研究 OCT4 参与调控多能性和早期分化过程的基因表达程序的分子机制。研究认为,OCT4 是多能细胞中核心转录调控网络的一部分。它的功能是单独或与其他转录因子协同识别和结合 DNA 调控区域。
在成熟时,OCT4 的表达仅限于发育中的生殖细胞。小鼠 OCT4 靶向破坏已产生无多能ICM 的胚胎,提示维持多能性需要 OCT4。此外,对 OCT4 表达量的定量分析表明,高水平的 OCT4 表达使胚胎干细胞向胚外中胚层或内胚层谱系发展,而低水平的 OCT4 表达使胚胎干细胞向胚外中胚层或内胚层谱系发展滋养层细胞;正常OCT4 水平的ES 细胞仍保留多能性。因此,有人提出 OCT4 是干细胞多能性和分化的关键调控因子。
已有的研究说明 OCT4 的多功能性,其既可以作为负责胚胎干细胞分化基因的抑制因子,也可以作为已知保留胚胎干细胞多能性基因的反激活因子。因此,OCT4可以认为是决定胚胎干细胞细胞自我更新和分化命运的主要因素,其在维持胚胎干细胞的多能性和自噬性方面发挥作用。
3 讨论
随着干细胞研究的热潮,现如今领域内对于 OCT4 基因的研究越来越深入。目前对于OCT4 基因的研究认为在鼠和人体内,OCT4 主要表达于胚胎干细胞及生殖细胞中,对于维持胚胎干细胞的多能性和自我更新有十分重要的作用,对于胚胎早期发育具有重大意义。但是目前 OCT4 在小鼠早期胚胎中的结合位点还不清晰,作为哺乳动物早期发育的关键调控因子之一,我们认为应进一步深入研究 OCT4 基因,通过 Low-Input Chip-seq 技术检测OCT4 在小鼠早期胚胎中的结合位点,可构建 OCT4 在小鼠早期胚胎发育过程中的调控网络,这对于揭开胚胎早期发育调控机制的面纱具有重要作用。
参考文献:
[1] 王凯.生命科学研究中常用模式生物[J].生命科學研究,2010,14(2):156-165.
[2] D. Tantin, Oct transcription factors in development and stem cells: insights and mechanisms[J]. Development,2013,140: 2857-2866.
[3] H.R. Schler, A.K. Hatzopoulos, R. Balling,et al. A family of octamer-specific proteins present during mouse embryogenesis: evidence for germline-specific expression of an Oct factor[J].EMBO J, 1989,8: 2543-2550.
[4] M.J. Lenardo, L. Staudt, P. Robbins, et al. Repression of the IgH enhancer in teratocarcinoma cells associated with a novel octamer factor[J]. Science,1989,243:544-546.
[5] K. Okamoto, H. Okazawa, A. Okuda, et al. A novel octamer binding transcription factor is differentially expressed in mouse embryonic cells[J].Cell,1990,60:461-472.
[6] M. Pardo, B. Lang, L. Yu, et al. An expanded Oct4 interaction network: implications for stem cell biology, development, and disease[J].Cell Stem Cell, 2010,6:382-395.
[7] D.L. van den Berg, T. Snoek, N.P. Mullin, et al. An Oct4-centered protein interaction network in embryonic stem cells[J].Cell Stem Cell, 2010,6:369-381.
[8] H.R.Schler,G.R.Dressler, R. Balling, et al. Oct4:a germline-specific transcription factor mapping to the mouse t-comple[J]. EMBO J,1990,9:2185-2195.
[9] D. C.Ambrosetti,C.Basilico,L.Dailey.Synergistic activation of the fibrolast growth factor 4 enhancer by sox2 and Oct-3 depends on protein-protein interactions facilitated by a specific spatial arrangement of factor binding sites[J].Mol.Cell.Biol,1997,17:6321-6329.
[10] K. Takahashi, S. Yamanaka. Induction of pluripotent stem cells from mouse embryonic and adult fifibroblast cultures by defifined factors[J]. Cell,2006(126):663-676.
