乳酸菌在肠道中介导MyD88特异性诱导RegⅢγ表达
2020-03-13沙万里刘东旭尹柏双李国江吉林农业科技学院吉林吉林市320吉林省预防兽医学重点实验室吉林吉林市320
沙万里,闫 满,刘东旭,尹柏双,李国江 (.吉林农业科技学院,吉林 吉林市320;2.吉林省预防兽医学重点实验室,吉林 吉林市320)
哺乳动物肠道内大约有1014个细菌[1],在健康动物中处于动态平衡状态,同时,固有免疫防御在防止微生物入侵中又起到至关重要的作用[2-3]。研究发现微生物能够刺激机体诱导胰岛再生源蛋白(regenerating islet-derived protein,Reg)的产生,该蛋白与细胞增殖[4]、抑制炎症[5]和细胞凋亡有关[6],其中Reg蛋白中的RegⅢ在防止共生菌和致病菌穿透肠道屏障方面至关重要[7]。抗菌肽RegⅢγ作为抑菌蛋白,其抗菌作用具有一定的选择性,它能够特异性的与肽聚糖结合,由于革兰阳性菌胞外肽聚糖较厚,因此更易与RegⅢγ结合并促使其对细胞壁的穿孔作用,同时RegⅢγ也能结合少部分的革兰阴性菌如鼠伤寒沙门菌,绿脓杆菌等[8]。
研究发现在肠上皮细胞表面和微生物间,分隔出大约50μm 的距离,RegⅢγ是维持这个空间结构的重要因素,通过对入侵微生物的抑制及对组织损伤的修复以维持肠道内环境的稳定[9]。RegⅢγ在肠道组织损伤后大量增加[10],但是关于肠道中RegⅢγ被调节表达的作用机理并不十分明确。研究发现当小鼠灌服鼠类柠檬酸杆菌后,IL-22 水平显著增高进而诱导肠道RegⅢγ大量表达以抑制鼠类柠檬酸杆菌感染肠道[11]。此外在肠道中除了IL-22能够诱导RegⅢγ表达外,Toll样受体(TLR)信号通路中的MyD88途径对RegⅢγ的调节也起到重要作用,当小鼠缺乏MyD88 基因时,小鼠肠道RegⅢγ表达量显著低于正常小鼠肠道中RegⅢγ的表达量,从而更易导致微生物入侵肠道的几率大大增加[12]。因此肠道微生物主要通过激活IL-22或TLR-My D88信号通路诱导抗菌蛋白RegⅢγ的产生,从而抑制细菌的侵入。然而作为益生菌的乳酸菌定植肠道是否同样诱导RegⅢγ的表达,以及其主要诱导途径尚不清楚。
本研究前期结果表明,混合乳酸菌具有刺激肠道上皮细胞使其分泌抗菌肽RegⅢγ的能力,可诱导小鼠肠道树突状细胞的分化成熟[13],但乳酸菌在肠道RegⅢγ表达通路中是否通过My D88信号通路尚不明确。本研究将通过灌胃小鼠混合乳酸菌,进一步探索RegⅢγ在肠道中的表达与组织修复过程中MyD88的特异性诱导途径。
1 材料与方法
1.1 材料小鼠:4~5周龄雌性SPF Balb/c小鼠(北京华阜康生物科技有限公司),饲养在独立通风饲养器中,食物和水都经过高压灭菌,成长到8~10周龄后,进行免疫试验。细菌菌株:Lactobacillus acidophilusATCC 4356,Lactobacillus rhamnosus
GG ATCC53103 由美国模式培养物研究所提供,Lactobacillus plantarumA 菌株接种于MRS培养基中37℃厌氧条件下培养。Escherichia coliBL21由吉林农业大学动物科技学院吉林省动物微生态制剂工程研究中心提供,菌株接种于LB 培养基中,37℃需氧条件下培养。
1.2 方法
1.2.1 乳酸菌灌胃方案Lactobacillus acidophilus
ATCC 4356、Lactobacillus rhamnosusGG ATCC-53103、Lactobacillus plantarumA 经MRS培养基37℃厌氧条件下培养,调制总量为3×109CFU/m L(低计量)和3×1010CFU/m L(高剂量)混合乳酸菌,磷酸盐缓冲液(PBS)清洗后备用。每只小鼠灌胃3×109和3×1010CFU/m L 菌体。连续灌胃2周,对照组自由饮水。2周后处死小鼠,取小肠与结肠。
1.2.2 RNA 的提取及Quantitative Real-time PCR
按照TaRa Ka Mini BEST Universal RNA Extraction Kit说明书方法对小鼠十二指肠与结肠进行总RNA 提取,参照TaRa Ka Prime ScriptTMRT Reagent Kit反转录试剂盒对完整的RNA 片段500 ng RNA 进 行cDNA 反 转 录 合 成,使 用SYBR 染 料及Step One Plus实时荧光定量PCR 系统对cDNA进行荧光定量。引物序列如下:
RegⅢγ-F:CCGTGCCTATGGCTCCTATTG,
RegⅢγ-R:GCACAGACACAAGATGTCCTG;
My D88-F:TCATGTTCTCCATACCCTTGGT,
My D88-R:AAACTGCGAGTGGGGTCAG;
IL-22-F:ATGAGTTTTTCCCTTATGGGGAC,
IL-22-R:GCTGGAAGTTGGACACCTCAA;
IL-12p40-F:TGGTTTGCCATCGTTTTGCTG,
IL-12p40-R:ACAGGTGAGGTTCATGTTTCT;
IL-23p19-F:CAGCAGCTCTCTCGGAATCTC,
IL-23p19-R:TGGATACGGGGCACATTATTTTT;
IL-17a-F:TTTAACTCCCTTGGCGCAAAA,
IL-17a-R:CTTTCCCTCCGCGCATTGACAC。
内参基因 gapdh-F:CACTCACGGCAAATTCAACGGCAC,gapdh-R:GACTCCACGACATACTCAGCA。Step One Plus软件通过2-ΔΔCt值法计算试验中各组基因的相对表达量。
1.2.3 RegⅢγ蛋白免疫荧光试验 小鼠肠道经福尔马林固定18 h后进行石蜡包埋,制备厚度为4μm的石蜡切片,经脱蜡后用10%小牛血清进行阻断。阻断后样品滴加兔抗鼠RegⅢγ一抗(1∶500稀释度,Bioss)4℃孵育过夜,然后滴加二抗(山羊抗兔Ig G,1∶250稀释度,Bioss)室温染色30 min,最后滴加4′,6-二脒基-2-苯基吲哚(DAPI,SIGMA)室温染色20 min。使用荧光显微镜(OLYMPUS IX51)观察各组样品荧光强度。
1.2.4 ELISA 试验 取小鼠小肠与结肠,37℃培养24 h[14],然后使用My D88、IL-22、IL-17 ELISA 试剂盒(R&D)检测肠道小肠与结肠中MyD88、IL-22、IL-17蛋白表达。
1.2.5 组织学评估 取小鼠小肠与结肠组织,制成石蜡切片,并对切片进行HE 染色以分析小肠和结肠中炎症情况。
1.2.6 统计学分析 使用GraphPad Prism 软件对试验数据进行统计分析,并通过T检验确定试验数据是否具有统计意义。P<0.05,数据具有统计意义。
2 结果
2.1 乳酸菌定植小肠与结肠中RegⅢγ表达水平
考虑到胃肠道的不同区域由于其细胞型、微生物成分等存在差别[14],因此本试验检测乳酸菌定植后小肠与结肠中RegⅢγ的表达情况,以确定乳酸菌在不同的肠道区域是否都能诱导RegⅢγ的表达。小鼠口服灌胃混合乳酸菌(3×109CFU/m L)2 周后,小鼠从单独的隔离器中取出,获取小鼠肠道组织以检测RegⅢγ的表达。通过RT-qPCR 我们成功地在小鼠小肠与结肠中检测到RegⅢγ mRNA 表达。结果显示混合乳酸菌在小肠与结肠中其诱导RegⅢγ表达的能力基本相同(图1)。免疫荧光显示(图2)小鼠服用混合乳酸菌后其小肠与结肠RegⅢγ 蛋白荧光强度明显高于对照组小鼠,与RTqPCR 结果一致。因此研究表明乳酸菌定植肠道后不受肠道区域情况限制都能够诱导RegⅢγ的表达。

图1 SPF小鼠和乳酸菌定植小鼠小肠及结肠中RegⅢγm RNA 的表达 ∗∗.P≤0.01;∗∗∗.P≤0.005。下同

图2 SPF小鼠3种混合乳酸菌定植小鼠小肠及结肠中RegⅢγ蛋白表达
2.2 乳酸菌定植数量与RegⅢγ表达水平的关系为了确定RegⅢγ表达水平是否与乳酸菌定植量相关,使用高、低2 种剂量(1×1010和1×109CFU/m L)的乳酸菌灌胃小鼠。2种剂量的乳酸菌连续在小鼠肠道定植2周后小鼠肠道中乳酸菌定植量呈现显著性差异。通过RT-qPCR 检测小鼠小肠与结肠中RegⅢγ的表达水平,结果显示服用高剂量乳酸菌的小鼠与低剂量组和对照组相比其小肠与结肠组织中RegⅢγ m RNA 表达水平显著增加(图3)。免疫荧光分析结果表明,与低剂量组和对照组相比,高剂量组的小肠与结肠中RegⅢγ蛋白表达也显著增强(图4)。
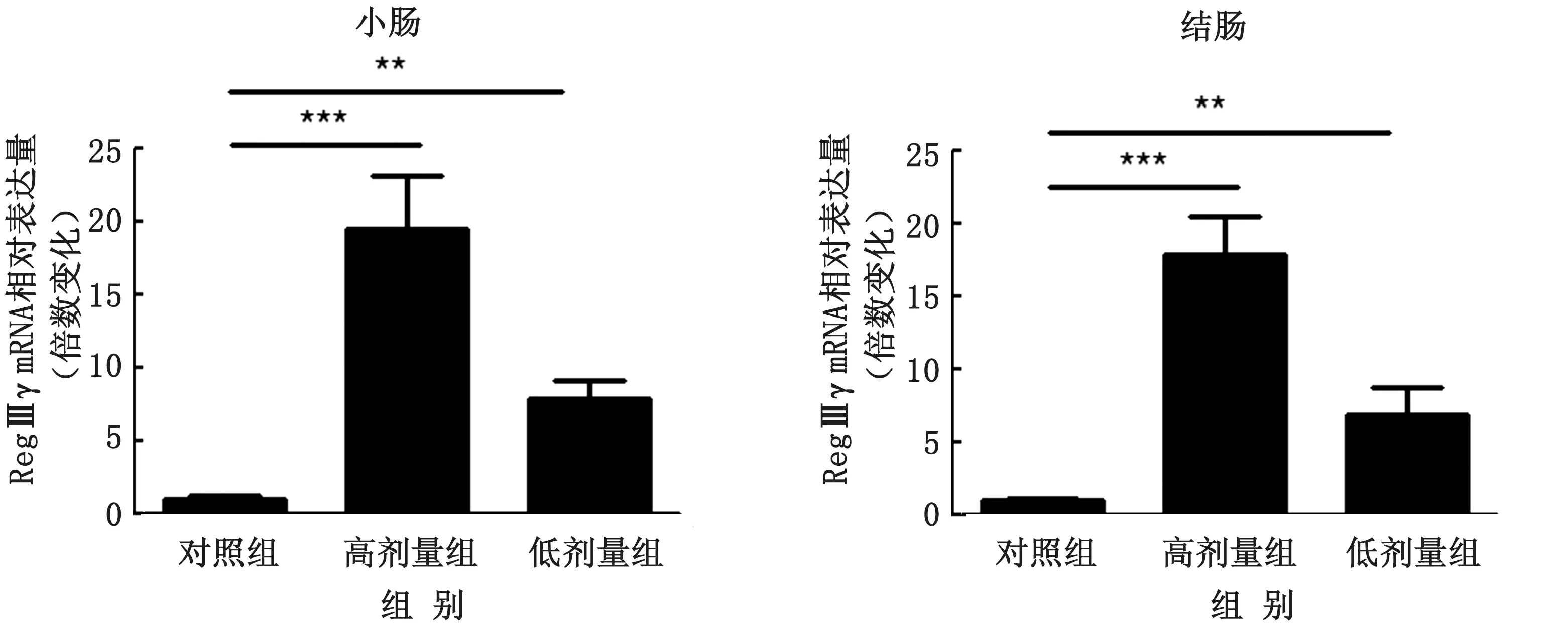
图3 SPF小鼠和不同剂量乳酸菌定植的小鼠小肠及结肠中RegⅢγ的表达
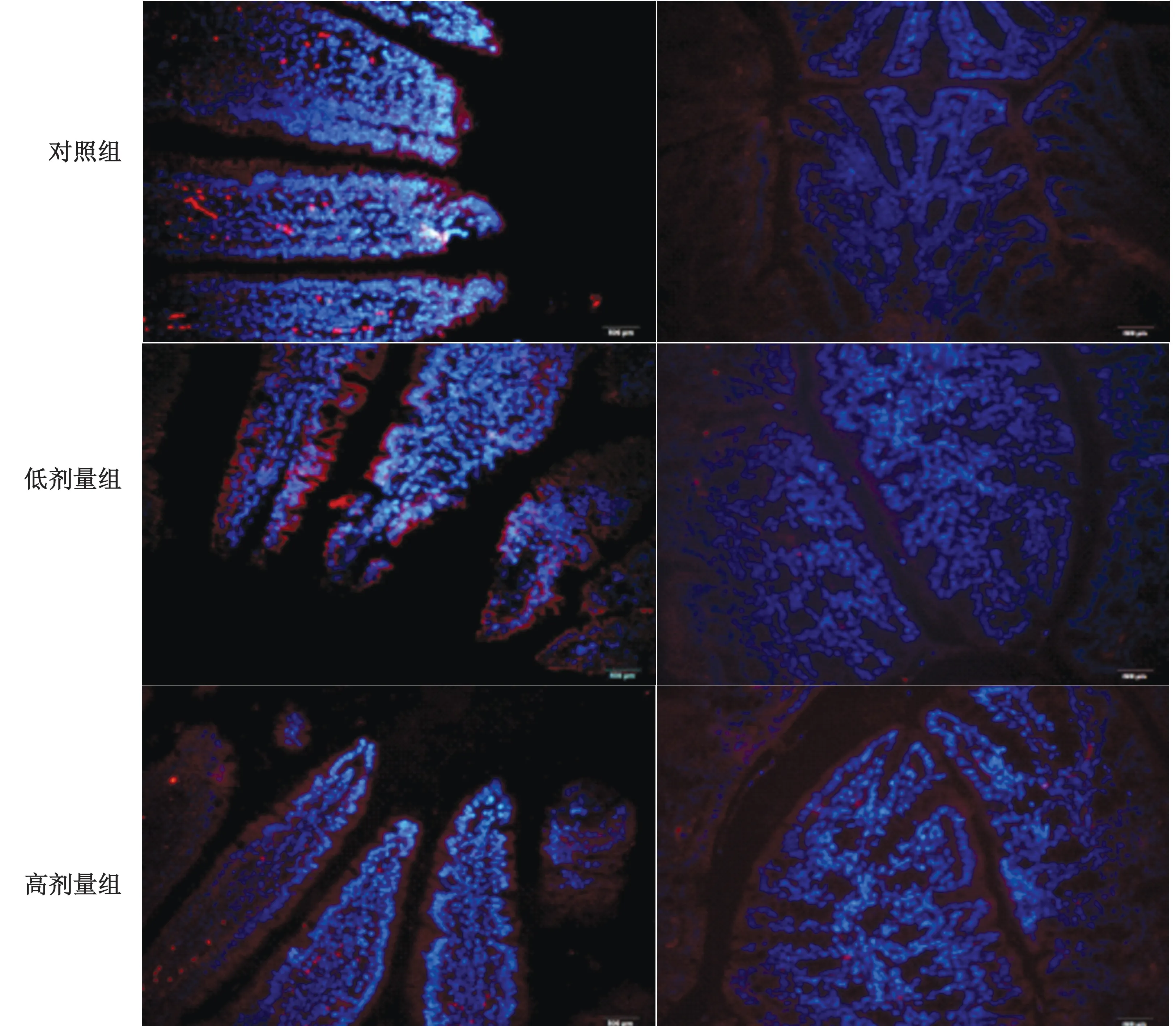
图4 SPF小鼠不同剂量3种混合乳酸菌定植小鼠小肠及结肠中RegⅢγ蛋白表达
2.3 乳酸菌诱导肠道RegⅢγ表达的主要途径为了检测乳酸菌诱导RegⅢγ表达的主要途径,小鼠口服灌胃混合乳酸菌,取小鼠小肠及结肠组织并通过RTqPCR检测肠道MyD88及IL-22、IL-17等Th17细胞因子反应情况。结果显示乳酸菌的定植能够诱导小鼠小肠与结肠中MyD88显著表达,同时在一定程度上还能诱导Th17细胞反应的发生。乳酸菌能够诱导IL-23、IL-22及IL-17的产生,但是随着乳酸菌剂量的增多Th17细胞反应并不明显增强,高剂量组与低剂量组IL-23、IL-22及IL-17表达水平基本维持在相近水平,但是整体均低于MyD88表达水平,同时ELISA结果显示MyD88蛋白水平高于Th17细胞因子水平与RT-qPCR结果一致(图5~6)。这表明乳酸菌主要通过MyD88特异性诱导途径介导RegⅢγ表达,同时对Th17细胞起到双向调控作用以维护肠道健康,HE染色结果验证了乳酸菌的定植虽然在一定范围内诱导Th17细胞反应但各组肠道均没有出现炎症症状(图7)。
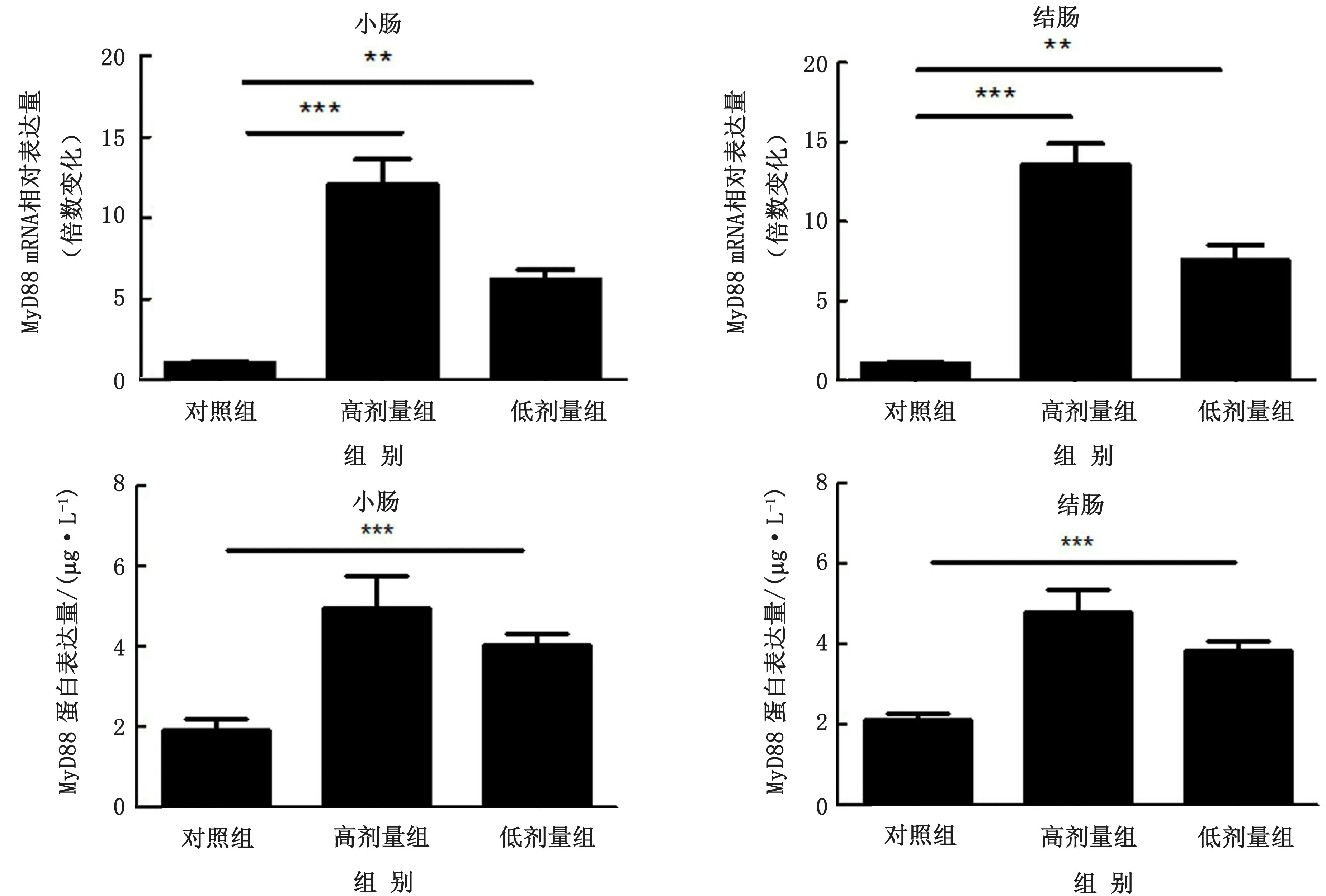
图5 SPF小鼠和不同剂量乳酸菌定植的小鼠小肠及结肠中MyD88的表达

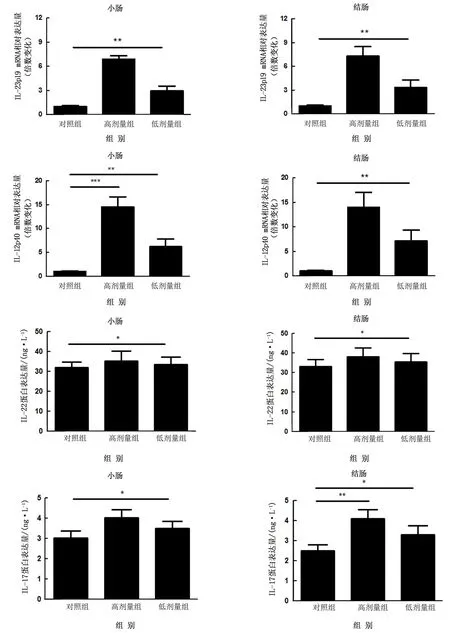
图6 SPF小鼠和不同剂量混合乳酸菌定植的小鼠小肠及结肠中Th17细胞因子的表达
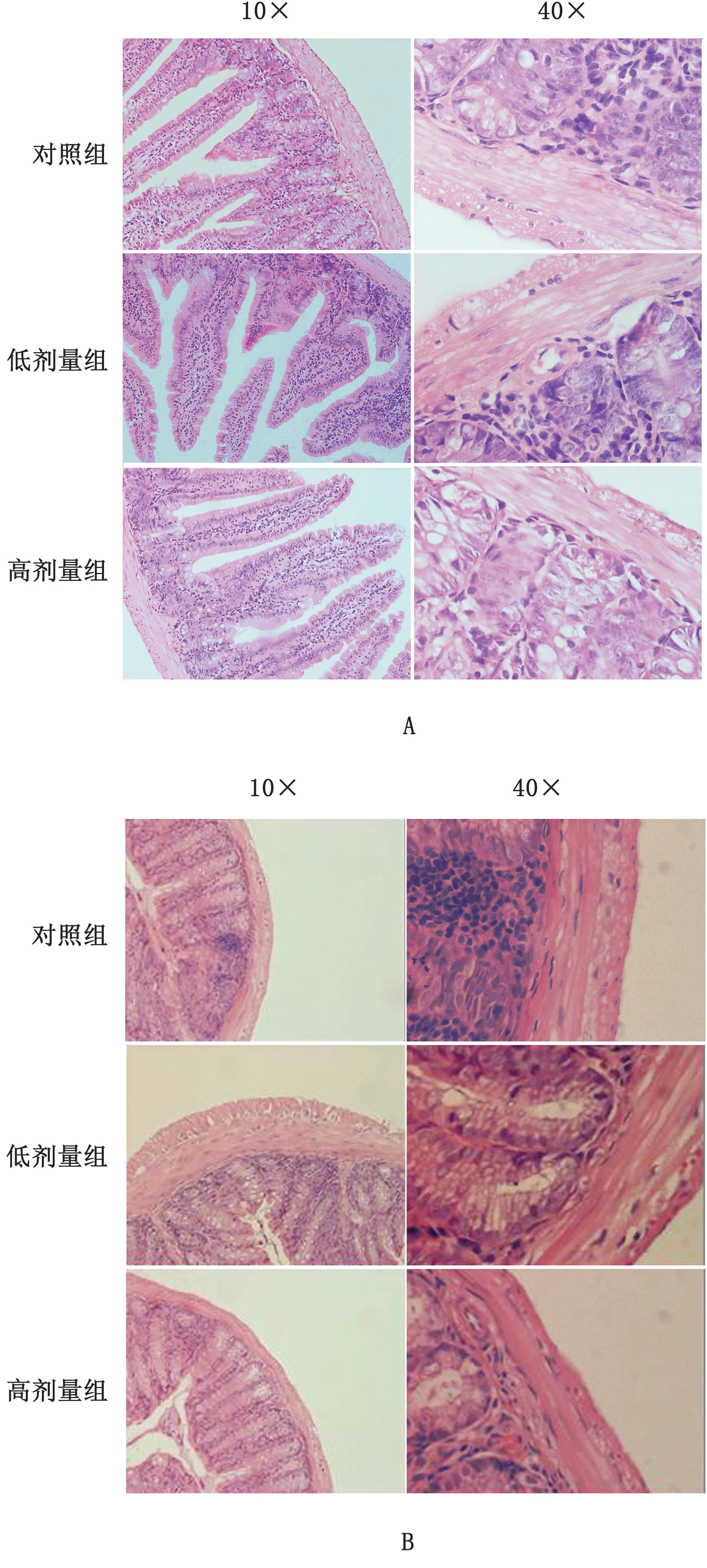
图7 SPF小肠及结肠H&E染色切片 A.小肠;B.结肠
3 讨论
抗菌肽RegⅢγ蛋白通过调节宿主肠道上皮与微生物之间的空间位置关系继而产生潜在的免疫防御以抵御病原微生物的入侵,因此RegⅢγ在维护肠道内环境稳定中起到重要作用。当前研究发现RegⅢγ不仅能够杀灭入侵的微生物,同时肠道微生物还能够影响肠道中RegⅢγ的表达水平[15]。
本研究发现RegⅢγ表达水平与乳酸菌定植及剂量相关。SPF级BALB/c小鼠服用由3种乳酸菌组成的混合益生菌在肠道定植后能够诱导小肠与结肠中RegⅢγ的表达,同时不同乳酸菌定植量诱导的RegⅢγ表达水平也显著不同,服用高剂量混合乳酸菌的小鼠与低计量乳酸菌组相比其小肠与结肠中RegⅢγ表达水平显著增高。这表明肠道中RegⅢγ的表达水平与乳酸菌的定植量相关,同时服用高剂量乳酸菌诱导RegⅢγ表达水平的增高也说明乳酸菌发挥益生作用的前提首先是要大量的定植。研究表明,肠道中潘氏细胞作为小肠特征性细胞能够分泌抗菌肽RegⅢγ,并且潘氏细胞在小肠中从十二指肠到回肠细胞数量呈逐渐增多现象,而在结肠中由于黏液层较厚不存在或少量存在潘氏细胞[16]。本研究结果却发现乳酸菌在小肠与结肠中其对RegⅢγ的诱导能力大致相同,结合已有研究证实的双歧杆菌主要通过激活TLR2/My D88信号途径继而诱导RegⅢγ的表达、肠道中RegⅢγ的表达受IL-22的调控以及小鼠注射鞭毛蛋白后当肠道固有层中的树突状细胞识别到抗原后迅速产生IL-23继而诱导肠道固有淋巴细胞分泌IL-22,IL-22促使上皮细胞修复的同时诱导RegⅢγ 的表达等基础研究[17]。本研究发现小鼠服用高、低剂量混合乳酸菌后在小肠与结肠中My D88 表达水平显著增多,并且高剂量组My D88 表达水平高于低剂量组,表明乳酸菌能够激活MyD88信号途径,同时IL-23、IL-22及IL-17等Th17细胞因子表达水平出现一定的增高,但是肠道结构并没有被破坏,也没有产生炎症反应,这表明乳酸菌能够双向调控Th17反应,既在一定程度上诱导Th17细胞反应又严格控制过度的Th17反应,其原因可能为机体适应性免疫成分控制RegⅢγ剧烈表达从而使益生菌在肠道定植。因此,乳酸菌诱导RegⅢγ的表达主要是受My D88途径的调节,
鉴于混合乳酸菌对RegⅢγ 的调节主要通过My D88途径特异性诱导表达,同时混和乳酸菌对Th17反应具有双向调控作用,既诱导其一定的表达又控制过度的炎症反应,其中Th17中的细胞因子IL-22对RegⅢγ的表达也具有一定的促进作用,与乳酸菌的定植对肠道炎症性疾病的发生具有预防作用的研究结论吻合[18]。根据以上发现推断,乳酸菌可能通过增加肠道中RegⅢγ的表达以加强肠道上皮的屏障功能以阻止病原菌的侵入及预防肠道炎症性疾病的发生。同时本研究提出这样的假设,乳酸菌通过刺激肠道上皮固有免疫反应来维护肠道健康从而实现了抑制炎症,因此推测乳酸菌通过介导My D88特异性诱导RegⅢγ表达以维护肠道内环境的稳定,可能对炎症性肠道疾病的研究提供一项新的治疗策略.
