历山典型植被土壤碳代谢指纹特征与差异
2020-03-11张勇强李智超厚凌宇宋立国孙启武
张勇强,李智超,厚凌宇,宋立国,孙启武
(中国林业科学研究院林业研究所,国家林业和草原局森林培育重点实验室, 林木遗传育种国家重点实验室,北京 100091)
土壤微生物与地上植被关系紧密,微生物通过多种方式对地上植被产生作用,地上植被也能够影响微生物[1-2]。森林是一个复杂的生态系统,其生物多样性显著,植物的空间异质性明显,不同经纬度、海拔、土壤类型、水热条件等环境因素的综合影响,决定了区域内的植物种类及微生物群落的异质性,构成复杂的系统,针对这些系统的微生物群落多样性及差异的研究意义重大。丰富的土壤微生物群落参与土壤中的养分的循环与转化,为植物的生长及发育提供能量基础[3]。植物群落对微生物多样性影响的不确定因素较多,而微生物也影响植物群落稳定性。因此有必要利用分子生物学手段,从整体上研究土壤微生物群落代谢多样性及其对土壤中各碳、氮源的利用情况[4-5]。
BIOLOG 板采用31 种底物诱导微生物代谢,通过显色反应来测算土壤微生物群落代谢功能多样性,用来描述土壤微生物群落功能的变化,具有简便、快速、灵敏和分辨力强等特点[6]。可利用其得到的吸光度平均变化值(AWCD)评价土壤微生物对碳源利用的整体情况及利用活性,通过计算多样性指数来评估群落中物种的丰富度、物种的优势度和物种的均一性[7-9],反映土壤微生物群落的组成及其稳定性,进一步了解土壤微生态的健康情况[9]。BIOLOG 作为简易的定量分析土壤及环境碳代谢指纹特征的技术措施,已广泛运用于农田土壤、污染土壤和林地土壤,但对林地土壤的研究主要集中于单一树种或人工林的研究,很少针对同一山体沿海拔梯度分布的典型天然植被土壤,且对于不同海拔土壤微生物分布格局的研究尚在起步阶段[10]。本研究旨在探讨典型森林土壤微生物群落代谢特征与差异,解析地上植被和土壤微生物群落的相互关系、不同植被类型下土壤微生物对各碳源的利用情况及微生物碳代谢多样性与土壤性质的相关性,同时探究土壤微生物的代谢多样性差异与海拔梯度分布格局、植被的关系,旨在为研究同一山体典型天然植被土壤碳代谢指纹特征与差异提供思路。
1 材料与方法
1.1 研究区域概况
历山拥有山西省重要的森林资源,有华北“绿肺”之称,该山处于中条山脉东段,动植物资源丰富,是研究典型天然植被土壤碳代谢指纹特征与差异的理想场所。该地是亚热带向暖温带的过渡区,气候适宜,为温带大陆性季风气候,年均温8~12 ℃,年降水量约700 mm主要集中于夏季,主要土壤类型为褐土,无霜期150~180 d,年均相对湿度70%。历山的主峰为海拔2 321.8 m 的舜王坪,四周草场覆盖,海拔与地理位置形成了历山从山顶至山脚丰富的植被带[11-12]。
1.2 样地布设及取样方法
以植被特征为依据,选择具有代表性的亚高山草甸(C1),华山松纯林(C2),红桦、榆树、五角枫、辽东栎阔叶混交林(C3),榆树、核桃楸、青皮椴、辽东栎阔叶混交林(C4),白桦、青皮椴、核桃楸、华北落叶松、油松针阔混交林(C5),白皮松、油松针叶林(C6),油松纯林(C7)7 种不同海拔典型森林(草甸)植被,每种植被选择3 块20 m×20 m 的样地,样地概况见表1。每块样地调查植物种类,采用S 型取样法取样方内5 点的0~20 cm 土壤样品,混合装入土袋,带回实验室放4 ℃冰箱中待测。
1.3 实验方法
用含有31 种碳源的BIOLOG-ECO 板,进行土壤微生物群落碳代谢实验。采用Classen 等[13]的方法制备生态板接种液。首先称取相当于30 g烘干土重的鲜土加入到盛有270 mL 浓度为0.85%的NaCl 溶液的三角瓶中180 r/min 震荡30 min,取3 mL 上清液加入到27 mL NaC1 溶液,混匀后再取3 mL 上清液加入到27 mL NaCl 溶液,土壤溶液最终稀释至10-3。用排枪吸取150 μL 的稀释液加入ECO 板的各孔中。将接种好的微孔板放在28 ℃的恒温培养箱中。分别于24、48、72、96、120、144、168 h 在Micro Log TM Version 4.20.05微孔板读数仪(Biolog Inc,USA)上进行测定,对波长90 nm(颜色+浊度)和750 nm(浊度)的数值进行处理计算。
1.4 数据处理
土壤微生物群落的功能多样性采用通过72 h的测定数据计算。微生物群落利用碳源的总能力用平均颜色变化率(AWCD)表示,计算方法如下:
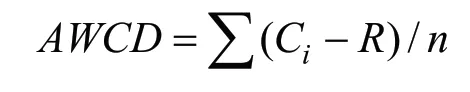
式中:Ci为第i个碳源孔的两波段光密度差值,R为对照孔的吸光度值,n为培养基碳源种类数Shannon-Wiener 指数(H′)表示被利用的碳源的总数,对稀疏种敏感,可用于评估物种的多样性[14],计算方法如下:
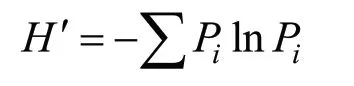
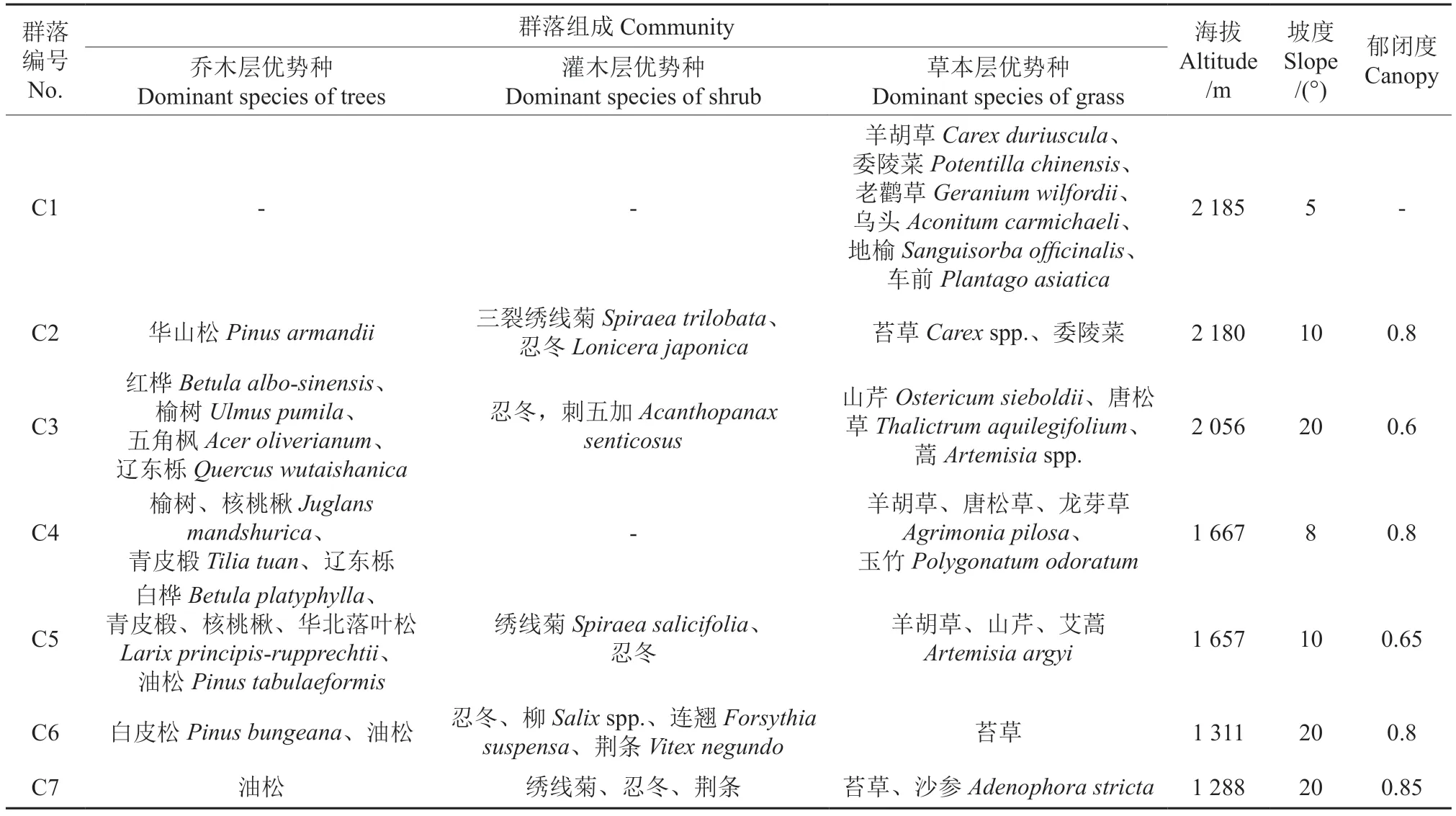
表1 样地基本情况Table 1 Basic condition of plots
式中:Pi表示第i个非对照孔中的吸光值与所有非对照孔吸光值总和的比值,即Pi=(Ci-R)/ Σ(Ci-R)[15]。
Pielou 指数(J)[16]可用于评估物种均匀度,计算方法如下:

式中:S为被利用的碳源总数。
Simpson 指数(D)[17]可用于评估物种丰富度,计算公式如下:

McIntosh 指数(U)是均一性的量度,是基于群落物种多维空间距离的多样性指数,计算公式如下:
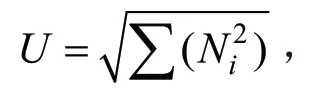
式中:Ni是第i孔的相对吸光值;
采用Excel 2003 统计并处理数据,R 软件对各时段AWCD 值及72 h 多样性指数进行单因素方差分析确定其显著性,对各典型植被进行主成分分析并计算得分,确定其土壤微生物主要利用碳源,用GraphPad PRISM®软件绘图。
2 结果与分析
2.1 土壤微生物群落对碳源的利用特性及不同碳源变化特征
2.1.1 土壤微生物对所有碳源利用的变化特征
土壤微生物对单一碳源的利用能力可以通过AWCD 值反映[18]。由图1 可知,随着培养时间的增加,所有植被类型AWCD 值不断增加,即微生物利用碳源量不断增加。培养时间24 h 以内,所有植被类型下AWCD 值最低,表明土壤微生物活性较低,碳源基本未被利用。24~48 h 内,AWCD 值变化的斜率最大,表明在这段时间内微生物对碳源的利用速率最高。72 h 以后,AWCD的变化速率逐渐降低,微生物对碳源的利用逐渐区域稳定。168 h 后,AWCD 值基本到达最大值,各植被类型下土壤微生物群落对碳源利用的大小顺序为:C5 >C1 >C4 >C3 >C6 >C7 >C2,且差异显著(P<0.05)。7 种植被类型中,白桦、青皮椴、核桃楸、华北落叶松、油松针阔混交林中土壤微生物群落代谢活性最高(1.265),其次为亚高山草甸(1.195),华山松最低(0.876)。
2.1.2 不同植被类型下土壤微生物对不同碳源利用的强度分析
BIOLOG 生态板的31 种碳源底物分为6 大类:碳水化合物、氨基酸、羧基酸、多聚类、酚酸类、胺类,其中,碳水化合物12 种,氨基酸6 种,羧酸类5 种,多聚类4 种,酚酸类2 种,胺类2 种。图2为6类碳源在7种典型植被下利用的整体情况,可以看出各类碳源在不同植被类型下差异显著。碳水化合物类是7 种植被利用的主要碳源,其次为氨基酸类或羧酸类,多聚物类、酚类和胺类的代谢较少。

图1 不同植被类型土壤微生物群落AWCD 随时间变化Fig.1 Changes of soil AWCD in different vegetation types with time
2.2 微生物代谢多样性
根据第72 h 的AWCD 值计算Shannon-Wiener多样性指数(H′)、McIntosh 多样性指数(U)、Simpson 优势度指数(D)和Pielou 均匀度指数(J)。由表2 可知,Shannon-Wiener 多样性指数(H′)C4、C1、C5 较高,均大于4,C3、C6 次之,在3~4之间,C7、C2 较小,均小于3,较高的Shannon-Wiener 指数代表微生物种类多且分布均匀,7 种不同植被类型下微生物群落多样性在榆树、核桃楸、青皮椴、辽东栎阔叶林最高,亚高山草甸,白桦、青皮椴、核桃楸、华北落叶松、油松针阔混交林中较高,红桦、榆树、五角枫、辽东栎阔叶林,白皮松、油松针叶林中次之,在油松、辽东栎针阔混交林较低,华山松纯林中最低。McIntosh 多样性指数(U)反映了碳源利用种类数的不同,能以此区分不同碳源的利用程度,Pielou 均匀度指数(J)反映了全部微生物种群个体数目的分配状况,二者与Simpson 优势度指数大致呈相同的大小规律,而Simpson 优势度指数(D)可反映群落中最常见物种,与前三者大小规律相反。
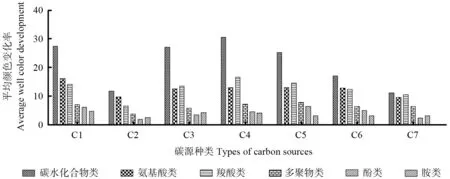
图2 土壤微生物对六类碳源的利用情况Fig.2 Utilization of six carbon sources by soil microorganisms
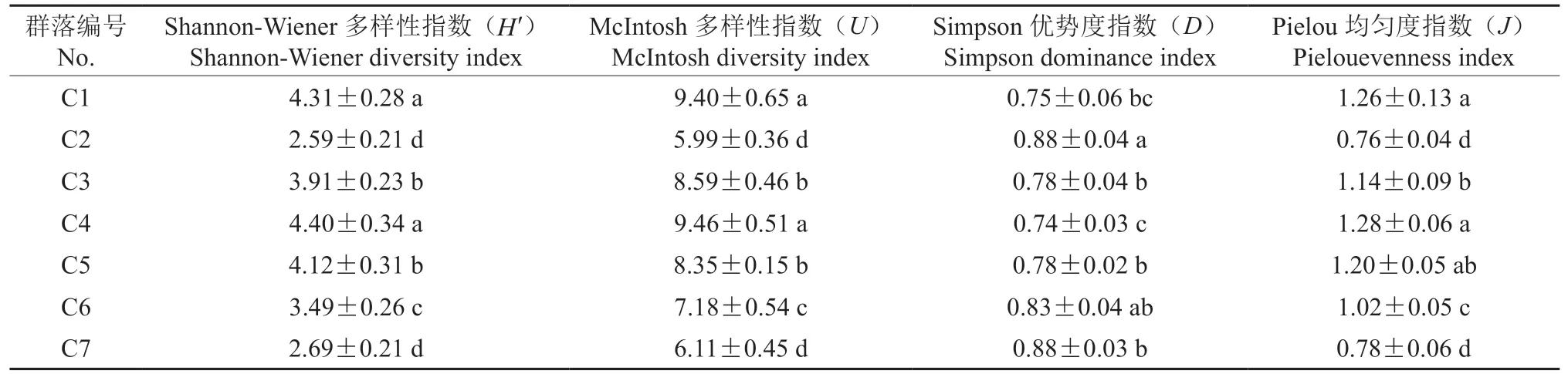
表2 不同植被类型土壤微生物群落多样性指数Table 2 Diversity indices for soil microbial communities of different vegetation types
2.3 土壤微生物群落碳代谢功能主成分分析
分别对7 种典型森林植被土壤微生物对碳源的利用进行主成分分析,提取前两个成分,并通过R软件程序包计算微生物对碳源利用情况得分,选择得分前5 的碳源对应微孔列于表3。微平板各孔所对应的碳源类型及种类如表4 所示。主成分分析结果显示:在不同植被类型下,土壤微生物主要利用的碳源类型差别明显,没有某一单一碳源在7 种植被类型中处于绝对优势的地位,不同于图2 不同类碳源的变化规律,表明图2 结果中不同类碳源代谢优势地位不是某一碳源代谢的结果,可能是由于该类别碳源种类多造成的。对于碳源代谢多样性的研究,表3 比图2 意义更大。
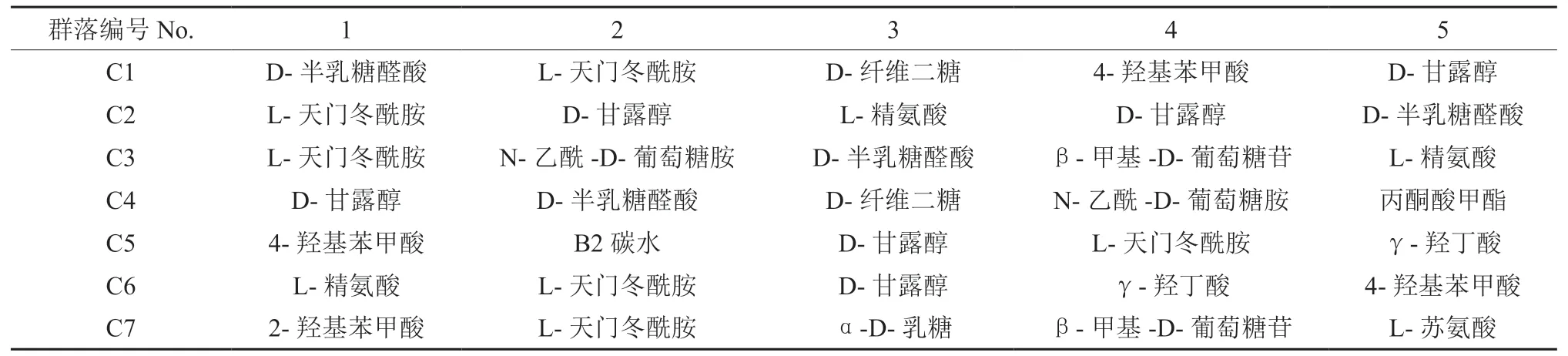
表3 主成分分析得分排名Table 3 Ranking of principal component analysis scores

表4 31 种碳源分类及对应微孔Table 4 Classification of 31 carbon sources and corresponding micropore
3 讨 论
3.1 典型植被土壤碳代谢差异
典型植被由于植物种类不同,所处环境不同,微生物群落具有较大差异,研究土壤微生物群落代谢多样性,对揭示植物-土壤-微生物之间的关系意义深远[19]。众多研究结果表明,土壤微生物群落代谢活性主要与枯落物数量和组成有关,不同植物产生不同种类不同数量的枯落物,影响土壤性质,从而造成土壤微生物的差异[20-22]。在吴则焰等[23]对武夷山青冈和米槠两种天然林及马尾松和杉木两种人工林土壤微生物多样性研究中,指出天然林土壤微生物群落多样性高于人工林,阔叶林高于针叶林。朱平等[24]在对祁连山垫状植被、高寒草甸、沼泽化草甸和高寒灌丛的研究中指出,沼泽化草甸和高寒草甸的土壤微生物对主要碳源的利用率高于灌丛草甸和垫状植被,分析认为,草地土壤表层根系密集,凋落物和根系分泌物为微生物提供丰富能源[23,25],枯落物数量较大且易分解,土壤含水率高,更适合微生物生长。邓娇娇等对不同利用方式土地土壤微生物的研究[26]指出,土壤微生物多样性阔叶林林>灌木林>针叶林>耕地,对黑土区水土保持林的研究[28]指出,土壤微生物多样性表现同样为阔叶林>针叶林。Hackl 等[29]对不同类型天然林土壤微生物群落组成研究中也指出,针叶凋落物含有大量木质素和丹宁酸,难以分解,需要特定的微生物负责降解这些顽固的有机化合物,因此导致土壤中微生物群落多样性较差。综合以上的结果及本文的研究,我们认为土壤微生物代谢多样性在典型森林(含草甸)土壤下应该具有以下规律:针阔混交林(以阔叶林为主)>草甸>阔叶林>针叶林。本研究中海拔1657 m 左右的针阔混交林,由于枯落物组成复杂且数量较多,水热条件较好,土壤微生物群落多样性较高,而海拔2180 m 左右的华山松纯林,虽然存在很厚的枯落物层,但枯落物种类单一,加之由于海拔的升高导致的水热条件变差,微生物及酶活性降低,其土壤微生物群落碳代谢活性最低。
3.2 土壤微生物代谢指纹特征
植物的组成及多样性与土壤微生物的生产及代谢活动是相互影响,相辅相成的[30],土壤微生物对各碳源的利用情况可以反映土壤碳源转化及土壤微生物代谢多样性[31]。综合以上研究结果,在BIOLOG 所研究的31 种碳源中,微生物对碳水化合利用最高,与之前研究一致[23,26,27],而通过对不同碳源得分利用率的得分排名在之前的研究中则很少体现,这一结果可以在一定程度上反映不同植物对微生物分解后产物的利用情况,为不同植被类型代谢所需营养物质种类及数量提供参考。通过对每种典型植被土壤微生物利用碳源情况分析,了解各类碳源在不同环境下微生物代谢的重要程度,以针对性的补充植物生长发育所必须的碳源,对森林植物的生长发育意义重大。
3.3 土壤微生物代谢多样性
土壤微生物群落结构多样性的主导因素是与土壤本身的理化性质和植被状况有关[32-34],生态学中采用不同的生态学指数反映群落内部物种总数和物种相对多度,多样性指数和丰富度指数代表微生物种类多且主要物种丰富,而均匀度指数反映土壤群落中普遍存在的物种。本研究中阔叶林中多样性指数和丰富度指数最高,其次为亚高山草甸和针阔混交林,但与其他人研究结果不同的是三种植被下均匀度指数较小,表明这三种植被类型中微生物种类多但分布不均匀,没有较突出的优势种,利于枯落物的分解和转化,促进养分的循环和积累。华山松纯林中多样性指数指数均最低,均匀度指数最大,表明其土壤中微生物种类较少,单一微生物种突出,不利于枯落物的分解转化,从而导致华山松体内养分积累慢,不利于其生长,这也是一些高海拔华山松树高和胸径受到限制的原因之一。
3.4 微生物多样性的影响因素
湿度、温度、光照等众多环境因素均受海拔变化的影响,研究海拔梯度对微生物多样性保护、生物代谢多样性分布特征及驱动因素的意义重大。Bryant JA 等[35]在对海拔梯度与微生物关系的研究中初次提出:土壤微生物多样性的海拔格局与植物多样性分布差异显著,与植物多样性的单峰模式不同,土壤细菌物种丰富度随海拔升高呈现单调递减模式。Shenc 等[36]在对长白山不同海拔植被土壤微生物研究指出,微生物多样性随海拔并没有明显的规律。Singh 等对日本富士山[37]北坡土壤的研究显示,土壤微生物沿海拔梯度具有单峰的分布格局,而对韩国汉拿山[38]的研究显示,土壤细菌多样性在中海拔较低,而低海拔和高海拔相对较高,表明山地生态系统中土壤微生物丰富度和多样性并没有一致的分布格局。此外,也有一些最新的研究利用高通量测序技术揭示了沿海拔下降的土壤微生物多样性分布模式[39,40]。根据目前的研究,土壤微生物多样性的海拔分布并未明确倾向于某种分布模式,本研究中土壤微生物的代谢多样性同样也没有与海拔梯度有某种确切联系,反而与植物的种类及多样性呈正相关关系,植物种类越丰富,其微生物代谢功能及多样性越强。
3.5 本研究局限及下一步研究方向
通过BIOLOG 技术,我们讨论了历山典型植被土壤碳代谢指纹特征与差异,虽对不同海拔植被类型都做了研究,但微生物碳代谢的影响因子并没有量化。海拔对微生物多样性的影响是众多环境因子共同作用的结果,由于时间的局限性,研究并没有对各植被类型的环境因子做出长期监测,以分析究竟哪一或哪些因子对微生物多样性的影响更为显著。另外,研究采取的方法较为简单,但同样存在局限性,其局限在于并未对微生物物种在某一水平上进行鉴定,无法比较微生物物种多样性在不同植被类型间的差异及海拔对不同物种微生物多样性的影响。针对以上局限性,我们下一步的研究方向为在各植被类型下设立观测点,以年为周期长期监测各环境因子,并在监测后对土壤进行重新采样,采用高通量测序等技术[41]对土壤微生物物种多样性进行鉴定。
4 结 论
本文调查研究了亚高山草甸(C1),华山松纯林(C2),红桦、榆树、五角枫、辽东栎阔叶混交林(C3),榆树、核桃楸、青皮椴、辽东栎阔叶混交林(C4),白桦、青皮椴、核桃楸、华北落叶松、油松针阔混交林(C5),白皮松、油松针叶混交林(C6)、油松纯林(C7)7 种典型森林土壤中的微生物多样性,结果显示,7 种典型森林土壤微生物对31 种碳源的综合利用能力顺序如下:C5 >C1 >C4 >C3 >C6 >C7 >C2,白桦、青皮椴、核桃楸、华北落叶松、油松针阔混交林土壤中微生物群落利用碳源的能力最强,华山松纯林最低。各多样性指数也反映出同样的规律。
各植被类型下土壤微生物群落对各碳源的利用的主成分分析研究结果显示,不同植被类型下土壤微生物代谢碳源程度各异,综合31 种碳源的利用,碳水化合利用率最高,其次依次为氨基酸、羧酸类、多聚类、酚酸类,胺类碳源的利用率最小。
土壤微生物代谢多样性与海拔梯度并无规律性关系。研究发现,土壤微生物代谢多样性与地上植物种类及多样性密切相关,地上植物种类和数量越丰富,其土壤中微生物代谢活动越强,代谢多样性越高。
