南方双季稻区不同复种方式对稻田综合温室效应的影响
2020-03-11伍思平肖小军吴自明曾勇军
伍思平 眭 锋 肖小军 张 俊 吴自明 曾勇军 黄 山
(1江西农业大学教育部/江西省作物生理生态与遗传育种重点实验室,江西 南昌 330045;2 江西省红壤研究所/国家红壤改良工程技术研究中心/农业农村部江西耕地保育科学观测实验站,江西 南昌 331717;3中国农业科学院作物科学研究所,北京 100081)
甲烷(CH4)和氧化亚氮(N2O)是导致全球变暖的重要温室气体[1-2]。大气CH4和N2O 浓度已经由工业革命前的715 μL·m-3和270 μL·m-3分别增加到1 803 μL·m-3和324 μL·m-3,并且以每年0.2% ~0.3%的速度增长[2]。全球农业生态系统每年温室气体排放总量为5.1~6.1 Pg,占人为活动所引起的温室气体排放总量的10%~12%[3]。我国是世界上最大的稻米生产国[4]。同时,稻田也是大气温室气体的重要排放源[2,5]。因此,研究如何协同实现水稻增产和稻田温室气体减排具有重要意义。
南方双季稻区是我国重要的粮油生产基地[6-7]。双季稻区具有长达5 个多月的冬闲季,期间种植紫云英或油菜,能够利用该地区冬闲季充足的光温水资源,提高双季稻系统的周年生产力[8-9]。在不同长期定位试验中,胡安永等[10]研究表明,稻田旱作期间CH4和N2O 排放量表现为冬小麦高于休闲和紫云英处理;与冬闲处理相比,冬小麦处理显著增加稻田全球变暖潜势,冬闲和紫云英处理间总变暖潜势无显著差异。相比于冬水田,水旱轮作有利于降低CH4排放,但会促进N2O 的排放[11]。张啸林[12]研究表明,双季稻-油菜轮作体系中CH4累积排放量显著高于单季稻-小麦轮作体系。由于农村劳动力的大量转移、种粮收益低等原因,双季稻区中稻的面积有增大趋势[13]。但是,前人研究多集中在双季稻系统下不同冬闲处理对稻田温室气体排放的影响[14-15],对单、双季稻系统的比较研究较少。为此,本研究于2016-2017 年在南方双季稻区开展田间试验,设置油菜-一季中稻、冬季绿肥(紫云英)-早稻-晚稻,冬闲-早稻-晚稻3 种轮作方式,监测不同复种方式的作物产量、CH4和N2O 气体排放,以期为南方双季稻田作物丰产和温室气体减排提供科学依据。
1 材料与方法
1.1 试验地概况
试验于2016 年10 月-2017 年10 月在江西省南昌市进贤县红壤研究所(116°17′E,28°35′N)进行。该试验地属于亚热带季风气候,2016-2017 年平均降水量1 588 mm。供试土壤为第四纪红色粘土发育的潴育型水稻土。试验前0 ~20 cm 耕作层土壤理化性质为pH 值5.9、有机质31.8 g·kg-1、碱解氮160.8 mg·kg-1、有效磷56.0 mg·kg-1、速效钾126.8 mg·kg-1。
1.2 试验设计
本试验采用完全随机区组设计,各小区面积均为48 m2。设置3 个处理:油菜-一季中稻(记作R-MR),油菜种子于2016 年10 月12 日(中稻收获后)人工均匀撒播入稻田,播种量为6.0 kg·hm-2,在2017 年5 月10 日人工收获后,将油菜秸秆切成约10 cm 小段原位还田,种植中稻;冬季绿肥(紫云英)-早稻-晚稻(记作GM-ER-LR),紫云英种子于2016 年10 月12 日(双季晚稻收获前10 d 左右)人工均匀撒播入稻田,播种量为30 kg·hm-2,在2017 年4 月28 日将紫云英原位翻耕入稻田,种植双季早稻和晚稻;冬闲-早稻-晚稻(记作WF-ER-LR),冬季休闲,种植双季早稻和晚稻。每个处理3 次重复。
供试品种:油菜品种为湘油1035,由湖南农业大学提供;冬种紫云英品种为余江大叶,为当地繁种;中稻品种为常规稻金农丝苗,由广东省农业科学院水稻研究所超级稻育种研究室提供;早、晚稻品种相同,均为常规稻中早39,购自江西进贤惠农种业有限公司。早稻收获后将秸秆切成约10 cm 小段原位还田,中稻和晚稻收获后秸秆均匀平铺在原位土壤表层。早、中、晚稻季秧苗均采用大田水育秧方式,早稻播种日期为为4 月8 日,秧龄28 d 移栽,栽插规格为25.0 cm×11.0 cm,基本苗为4 株/穴;中稻播种日期为5 月11日,秧龄25 d 移栽,栽插规格为25.0 cm×15.0 cm,基本苗4 株/穴。晚稻播种日期为7 月6 日,秧龄20 d移栽,栽插规格为25.0 cm×11.0 cm,基本苗为3 株/穴。油菜季和中稻季氮肥、磷肥和钾肥用量相同,分别为纯氮150 kg·hm-2、P2O567.5 kg·hm-2和K2O 120 kg·hm-2。其中油菜季以包膜尿素作为氮肥,按基肥∶苗肥∶薹肥=5∶2∶3 施用,中稻季以常规尿素作为氮肥,按基肥∶分蘖肥∶穗肥=5∶2∶3施用;油菜季和中稻季均以钙镁磷肥作为磷肥,做基肥一次性施用;油菜季和中稻季均以氯化钾作为钾肥,油菜季按基肥∶薹肥=5∶5施用,中稻季按基肥∶穗肥=5∶5施用。早晚稻季均以常规尿素作为氮肥,施用量为纯氮120 kg·hm-2,磷肥和钾肥施用量和施用方式与中稻季一致。
在油菜季、紫云英季和休闲季田间水分自然排干;在早、中、晚稻季田间水分管理模式为秧苗移栽后保持3 cm 左右水层,分蘖末期排水烤田,复水后采用干湿交替灌溉方式,直至水稻收获前10 d 左右稻田水分自然排干。早、中、晚稻季开始排水晒田日期分别为5 月30 日、7 月5 日和8 月12 日,复水日期分别为6 月14日、7 月20 日和9 月1 日,试验期各处理水层深度动态变化如图1 所示。本试验始于2015 年,气体监测从2016 年油菜季开始。

图1 试验周期内田间水层深度和气温动态变化Fig.1 The dynamic change of water depth and air temperature during the experimental period
1.3 测定项目与方法
1.3.1 温室气体排放 采用静态暗箱-气相色谱法监测田间温室气体的排放通量[16]。暗箱规格为50 cm×50 cm×50 cm,暗箱由不锈钢金属材质制成,当水稻或油菜高度>50 cm 时,需额外增添一个或两个相同尺寸的双向开口的箱体,此时箱体总高度为100 或150 cm。箱体表面包裹海绵和锡箔纸,避免气体采集过程中太阳照射导致暗箱内温度升高过快。暗箱内顶部安装一个电压为12 Ⅴ的小型风扇,外接便携式蓄电池,气体采集过程中,使暗箱内气体充分混匀。油菜季、紫云英季和冬闲季温室气体采集时,在油菜和紫云英种植小区内,选取出苗均匀、长势基本一致的地点,固定一个上部留有3 cm 深的凹槽,下部15 cm 深的底座,将底座直接插入土壤。在早稻季、中稻季和晚稻季,稻田翻耕前底座移出稻田,秧苗移栽后,底座再次埋入稻田。早晚稻季底座内秧苗均为8 穴,中稻季底座内秧苗为6 穴。田间气体采集时,暗箱与底座或暗箱与暗箱连接处的凹槽内注水以隔绝空气。水稻生长季,稻田温室气体每7 d 采集一次,油菜季、紫云英季和冬闲季每15 d 采集一次。气体采集时,同步记录田间气温动态变化(图1)。田间气体采集时间为上午9:00-11:00,分别在0、10、20、30 min 用50 mL 针孔注射器来回抽取箱内气体多次后,采集50 mL 气体转入100 mL 预先抽真空采气袋,迅速带回实验室用Agilent 7890b 气象色谱仪(美国安捷伦科技有限公司)测定样品CH4和N2O 气体浓度。CH4和N2O 检测器分别为氢火焰离子化检测器(flame ionization detecter,FID)和电子捕获检测器(electroncapture detector,ECD)。温室气体测定时仪器具体参数设定参照文献[17]。温室气体排放通量计算公式如下:

式中,F 为温室气体排放通量(CH4,mg·m-2·h-1,N2O,μg·m-2·h-1);ρ表示标准状态下的气体密度(kg·m-3);h 为采气暗箱的净高度(cm);dc/dt 为单位时间内暗箱内温室气体的排放速率;T 为采气过程中暗箱内的平均温度(℃);273 为气态方程常数。
根据测定CH4和N2O 气体浓度与气体采集时间的关系曲线计算温室气体排放通量(拟合曲线相关系数大于0.90);当测定的CH4或N2O 气体浓度较低时,气体浓度与时间拟合曲线相关系数大于0.80 亦可。加权平均得到试验周期内CH4累积排放总量[Ac(CH4),kg·hm-2]和N2O 累积排放总量[Ac(N2O),g·hm-2]。
1.3.2 综合温室效应、籽粒能量产出和全球增温潜势强度 综合温室效应在100 年时间尺度上,单位质量的CH4和N2O 的综合温室效应(global warming potential,GWP)分别为CO2的25 倍和298 倍[2],其计算公式如下:

采用全球增温潜势强度(global warming potential intensity,GWPI)表示温室气体的排放强度。GWPI 表示生产单位等效籽粒能量产出(grain energy yield,GEY)的稻谷对气候变暖的潜势(以每kg 稻谷产量的综合温室效应CO2当量计算)[2,18],GEY 和GWPI 计算公式如下:


水稻籽粒能源转换因子为14.5 MJ·kg-1,油菜籽粒能源转换因子为29.9 MJ·kg-1。采用作物的等效籽粒能量产出计算GWPI,而不是作物的绝对产量,以消除油菜和水稻的物种差异。
1.3.3 产量 在油菜和水稻成熟期,各小区人工收割10 m2,机械脱粒,称重,取1 kg 烘干测定含水量,计算干重再统一换算成14%的含水率,测定各小区的实际产量(kg·hm-2)。
1.4 数据处理
采用Origin 9.0 软件作图,运用SPSS 18.0 进行数据处理,采用LSD 法于P=0.05 水平上进行显著性检验。
2 结果与分析
2.1 CH4 排放通量季节变化
在本试验周期内,各处理CH4排放通量的季节变化范围为-0.05 ~53.83 mg·m-2·h-1(图2)。在油菜季、紫云英季和冬闲季,CH4排放通量均趋近于零,而在早、中、晚稻季,CH4排放通量动态变化幅度较大。在R-MR 的中稻季,水稻移栽后CH4排放通量逐渐增加,在6 月29 日监测到排放最高峰,中期排水晒田后迅速下降,后期覆水及干湿交替灌溉模式中未监测到较大排放峰。在GM-ER-LR 和WF-ER-LR 的早稻季,水稻移栽后CH4排放通量迅速增加,均在5 月26 日监测到排放最高峰,中期排水晒田后亦迅速下降,复水后施用穗肥,在6 月23 日监测到较大排放高峰,直至水稻收获前一周,CH4排放通量趋近于零。而在GMER-LR 和WF-ER-LR 的晚稻季,水稻移栽后淹水条件下,前三次CH4排放通量均监测到较大排放高峰,中期排水晒田及后期干湿交替灌溉模式中,CH4排放通量趋近于零。GM-ER-LR 和WF-ER-LR 试验周期内CH4累积排放总量均显著高于R-MR,而GM-ER-LR与WF-ER-LR 间无显著差异(表1)。

图2 不同复种方式对稻田CH4 排放动态的影响Fig.2 Effects of different multiple cropping pattern on CH4 emission fluxes

表1 不同复种方式对稻田CH4 累积排放总量的影响Table 1 Effects of different multiple cropping regimes on total CH4 emissions /kg·hm-2
2.2 N2O 排放通量季节变化
由图3 可知,各处理N2O 排放规律不一,R-MR在3 月23 日、GM-ER-LR 及WF-ER-LR 在2 月6 日均监测到排放最高峰。3 个处理在水稻生育前中期N2O 排放通量趋近于零,均表现为大气N2O 较弱的源或汇。R-MR 临近中稻收获前仅监测到一次排放高峰(9 月3 日),GM-ER-LR 和WF-ER-LR 在晚稻季复水后施用穗肥及后期干湿交替灌溉模式下,N2O排放通量动态变化幅度很大,两处理均在9 月17 日监测到排放最高峰。R-MR 试验周期内N2O 累积排放总量显著高于GM-ER-LR 和WF-ER-LR,分别提高51.8%和97.7%,而GM-ER-LR 与WF-ER-LR 间无显著差异(表2)。

图3 不同复种方式对稻田N2O 排放动态的影响Fig.3 Effects of different multiple cropping pattern on seasonal variations of N2O emission fluxes
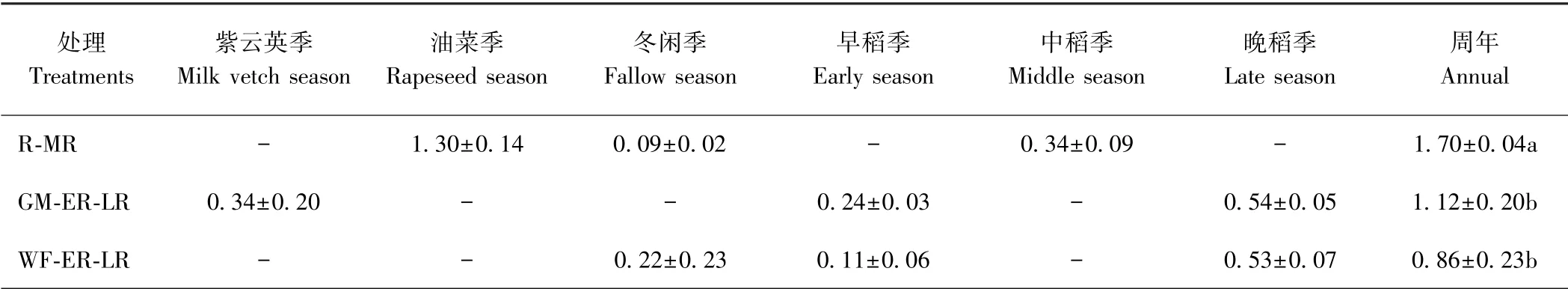
表2 不同复种方式对稻田N2O 累积排放总量的影响Table 2 Effects of different multiple cropping pattern on total N2O emissions /(g·hm-2)
2.3 稻田GWP、GEY、GWPI 和周年总产量
GM-ER-LR 和WF-ER-LR 试验周期内稻田GWP、GEY 和GWPI 均显著高于R-MR(表3)。与WF-ERLR 相比,GM-ER-LR 周年总产量提高1.2%,而GWP增加4.3%,GM-ER-LR 和WF-ER-LR 两处理间GWPI无显著差异,分别较R-MR 提高100%和50%。
3 讨论
3.1 不同复种方式对稻田CH4 和N2O 排放的影响
本研究结果表明,稻田CH4排放主要集中在水稻季(早、中、晚稻季);而在非水稻季(油菜季、紫云英季和冬闲季),稻田CH4排放通量均趋近于零,这与前人研究结果相同[19-21]。究其原因,主要是在水稻季,水稻前期淹水条件下,秸秆有机物还田为产甲烷菌提供本底基质导致CH4排放速率加剧[21-22],且水稻分蘖前期地上部和地下部生物量较小,通气组织不发达,对稻田CH4氧化能力有限[23];而在非水稻季,田间管理为水分自然排干,甲烷氧化菌占绝对主导地位,即使能产生CH4,大部分也被氧化了[22]。本研究中,与RMR 相比,GM-ER-LR 和WF-ER-LR 显著增加了试验周期内稻田CH4累积排放总量,主要是因为R-MR水稻周年累积淹水天数(60 d)低于GM-ER-LR 和WF-ER-LR(83 d),稻田淹灌环境下有利于产甲烷菌的存活从而促进稻田CH4的排放[24-25]。且R-MR 油菜秸秆还田量低于GM-ER-LR 紫云英加早稻秸秆还田量和WF-ER-LR 早稻秸秆还田量,有机物外源添加显著提高稻田CH4累积排放总量[21,26-27]。此外,R-MR 稻季周年施氮量(150 kg·hm-2)低于GM-ER-LR和WF-ER-LR 中双季早晚稻氮素施用总量(240 kg·hm-2),CH4排放通量随施氮量的增加而增加[28]。本研究中双季晚稻的日排放量较RM-R 的中稻低,主要是南方双季晚稻区稻田整体气温较低,抑制了产甲烷菌的活性[29-30]。

表3 不同复种方式对稻田综合温室效应、温室气体排放强度和产量的影响Table 3 Effects of different multiple cropping pattern on the GWP,GEY,GEPI and crop yield
在非水稻季,3 种轮作方式中稻田N2O 排放通量动态变化幅度均较大,并且在中、晚稻季生育后期也监测到较大幅度的N2O 排放通量。其原因可能是本试验在非水稻季稻田采取水分自然排干及在R-MR 油菜季施用氮素,且在水稻季生育后期采取干湿交替灌溉模式。这种水分管理模式和氮素施用均可能会引起土壤微生物硝化与反硝化过程加剧,促进稻田N2O 气体排放[17,24,31]。而在早稻季和中稻季前中期均未监测到N2O 排放高峰,原因可能是在南方双季稻区此季节经常出现连续阴雨天气,当稻田土壤孔隙含水量较高时(>90%),土壤孔隙中O2利用率极低,影响土壤微生物硝化-反硝化进程[21-22],从而抑制稻田N2O 产生及排放。本试验中R-MR 试验周期内N2O 累积排放总量显著高于GM-ER-LR 和WF-ER-LR。在R-MR 试验周期内N2O 累积排放主要是在油菜季,占稻田周年N2O 累积排放总量的76.5%。黄太庆等[32]研究表明,在非水稻季,油菜施氮处理N2O 季节累积排放量显著高于冬闲处理,本研究结果与之相似,即油菜施氮处理为土壤硝化和反硝化微生物提供足够的N 源。此外,本研究还发现,GM-ER-LR 紫云英季N2O 累积排放总量高于WF-ER-LR 冬闲季,这与前人研究结果类似[30],主要原因是紫云英根系较发达,会显著提高土壤中有机碳含量和土壤碳库管理指数,从而促进N2O排放量;其次,化学氮肥的施用,也是促进稻田N2O 排放的主要因素。
3.2 不同复种方式对稻田GWP、GEY、GWPI 的影响
本研究结果表明,GM-ER-LR 和WF-ER-LR 周年GEY 显著高于R-MR。尽管R-MR 中稻季产量高于GM-ER-LR 和WF-ER-LR 的双季早稻或晚稻季产量,且油菜能量转换因子>水稻能量转换因子,但其周年GEY 累积总量仍低于GM-ER-LR 和WF-ER-LR。此外,GM-ER-LR 周年总产量最高,较WF-ER-LR 提高1.2%。在本试验3 种复种方式中,GM-ER-LR 周年总产量提高主要体现在早稻季。主要原因可能是GMER-LR 在非水稻季种植紫云英,紫云英翌年直接翻入土壤,有利于土壤培肥,促进作物养分吸收从而提高产量[8]。GM-ER-LR 和WF-ER-LR试验周期内稻田GWP 显著高于R-MR,其GWPI 较R-MR 也显著增加,这可能与GM-ER-LR 和WF-ER-LR 中CH4排放量高有很大关系。本试验中GM-ER-LR 和WF-ER-LR稻田N2O 累积排放总量极低,但CH4累积排放总量显著高于R-MR。前人研究发现,在稻作系统中GWP 的大小主要由CH4累积排放总量决定[22,24,33],本研究结果与之类似。GM-ER-LR 和WF-ER-LR 虽然较显著增加周年总产量,但也显著提高稻田GWP 和GWPI,为全球气候变暖带来一定的风险。本研究仅为1 年田间试验,研究期限较短,今后会继续对不同轮作方式的双季稻田温室气体排放进行长期监测。
4 结论
本研究结果表明,与R-MR 相比,GM-ER-LR 和WF-ER-LR 均显著提高试验周期内CH4累积排放总量,但也显著降低了N2O 累积排放总量。与R-MR 相比,GM-ER-LR 和WF-ER-LR 均显著提高周年GEY,但也增加了试验周期内稻田GWP。综上,与R-MR 相比,虽然在南方双季稻区GM-ER-LR 和WF-ER-LR 均增加了粮食产出,但也会增加稻田的温室效应和温室气体排放强度。因此,鉴于双季稻区中稻面积有增大趋势,综合环境效益和经济效益,油菜-一季中稻的增产减排效果更好,可为南方双季稻低碳高效栽培提供理论依据。
