亚硝酸钠诱导的草鱼肝细胞凋亡中内质网应激IRE1通路的作用研究
2020-03-04陈思琪谢丽霞姚朝瑞李大鹏
陈思琪 谢丽霞 姚朝瑞 李大鹏 汤 蓉
(华中农业大学水产学院, 池塘健康养殖湖北省工程实验室, 农业部淡水生物繁育重点实验室, 武汉 430070)
在生态系统中, 亚硝酸盐是氮循环的一个天然组成部分, 现已成为水产养殖系统中一个潜在性问题[1]。在高密度集约化养殖系统中, 由于放养密度增大以及过度投饵造成的高蛋白残饵和高含氮排泄物不断沉积于池底, 超过养殖水体代谢能力, 硝化细菌的消化平衡遭到破坏, 进而导致亚硝酸盐的积累,成为水产养殖过程中面临的重要胁迫因子[2]。水体中亚硝酸盐是由氨通过硝化细菌硝化作用形成的,其可以在水生系统中积累到很高的浓度[1]。已有研究表明, 养殖水体中高浓度亚硝酸盐和氨氮是诱发鱼病主要的环境因子[3], 亚硝态氮和氨氮引起的水质恶化受到了养殖界和相关学者的高度重视。
水体中亚硝酸盐浓度升高会对水生动物产生多种生理干扰, 如生长抑制、组织损伤、内脏功能紊乱和内分泌紊乱等[4]。亚硝酸盐暴露对鱼、虾和蟹等养殖对象均具有一定的毒性作用, 影响生长发育, 诱发组织病理变化, 降低抵抗力, 甚至导致死亡[5]。水体中的亚硝酸盐通过鳃上皮细胞进入鱼体, 主要将血红蛋白氧化成高铁血红蛋白, 从而导致血液载氧能力逐渐降低, 造成组织缺氧、神经麻痹、摄食量降低、鳃组织出现病变、呼吸困难、骚动不安或反应迟钝, 严重时则使生物发生暴发性死亡[6,7]。此外, 水体中亚硝酸盐浓度超过一定浓度会引起应激, 从而诱导动物细胞凋亡[8]。目前关于亚硝酸盐对细胞凋亡的研究报道主要集中在个体水平研究,通过细胞水平揭示亚硝酸盐引起细胞凋亡的研究相对较少, 其潜在的作用机制还有待阐明。细胞凋亡存在于机体的整个生命过程, 参与了各种生理及病理过程。现已有研究发现内质网也参与细胞凋亡的途径。内质网是蛋白质折叠和转运的专用细胞器, 对细胞内稳态和细胞外刺激的变化高度敏感[9]。环境应激引起的未折叠、错折叠、突变蛋白积累均可作为应激信号干扰内质网稳态并引起内质网应激(Endoplasmic reticulum stress, ER Stress)。为了应对这种环境应激, 机体内质网会作出未折叠蛋白反应(Unfolded protein response, UPR)的保护性或适应性策略, 以此恢复内质网的稳态[10], 如果内质网应激的不良作用过大, 细胞开始凋亡。UPR有三个通路: IRE1、PERK和ATF6。IRE1是一个内质网I型跨膜糖蛋白, 在胞内段具有激酶活性和RNA酶活性。在哺乳动物细胞中有 IRE1α和IRE1β 两种亚型, IRE1α普遍存在于所有的组织细胞中, 而 IRE1β 主要存在于肠上皮细胞中[11]。内质网应激能导致IRE1腔内结构域与BiP解离, IRE1寡聚化, 自身磷酸化激活其RNA酶活性[12]。两者的底物都是X-box 结合蛋白 1(XBP1) mRNA, 其编码碱性含有亮氨酸锌指结构的转录因子。IRE1的RNA酶活性切割 XBP1 mRNA, 使其成为成熟的 mRNA,其编码的 XBP1 蛋白能增强分子伴侣蛋白 BiP 等的转录活性[13]。在持续性的内质网应激条件下曾观察到IRE1信号通路关闭, 而PERK信号通路仍然发挥作用直至细胞死亡。因此有研究认为UPR的IRE1通路分支主要参与存活反应[14], 而近来研究更是表明IRE1通路是决定细胞生存或者凋亡的关键途径[15]。
草鱼(Ctenopharyngodon idella)属鲤形目鲤科雅罗鱼亚科草鱼属。因其生长迅速, 饲料来源广,是我国淡水集约化系统的主要物种之一, 目前正遭受着由氨氮和亚硝酸盐等水污染引起的环境压力。水产养殖中亚硝酸盐含量的高低决定着养殖水质的好坏, 亚硝酸盐含量高对养殖动物带来很大的危害, 当前水产养殖集约化高密度养殖导致很多池塘亚硝酸盐含量超标。肝脏作为鱼类体内主要的解毒器官, 其在应对外界污染物胁迫中起到关键性作用, 同时也是亚硝酸盐主要靶器官之一[16—18]。本实验以草鱼肝细胞系L8824为实验对象, 进行亚硝酸钠的急性暴露, 通过检测细胞凋亡相关基因表达、细胞凋亡和细胞中内钙离子浓度变化, 探讨亚硝酸钠暴露对草鱼肝细胞的凋亡影响以及内质网应激通路中IRE1通路在其中发挥的作用, 以阐明亚硝酸钠导致草鱼肝细胞凋亡中可能的致毒机制, 以期为草鱼的科学化养殖提供可靠的科学理论依据。
1 材料与方法
1.1 材料
草鱼肝细胞系(L8824)购自中国典型培养物保藏中心; M199培养基、胎牛血清、0.25%胰蛋白酶-EDTA消化液(Trypsin-Ethylene diamine tetraacetic acid, Trypsin-EDTA)购自Gibco公司; CCK8试剂盒购自上海翊圣生物科技有限公司; Annexin-FITC/PI细胞凋亡试剂盒购自南京建成生物工程研究所;STF-083010和2-APB购自美国APExBIO公司。
1.2 细胞培养
L8824培养于含10%胎牛血清的M199培养液中, 置于28℃、5% CO2恒温恒湿培养箱中培养。细胞汇合达80%—90%时, 用0.25%胰蛋白酶消化,离心, 常规传代。
1.3 CCK-8法检测细胞活力
将处于对数生长期状态良好的细胞以5×105/mL密度接种于96孔板, 每孔100 μL, 培养箱中预培养24h后分别换为不同浓度的亚硝酸钠(0、5、20和50 mg/L) M199培养基分别孵育12h和24h, 每组4个平行, 在培养结束后, 每孔加入10 μL CCK8试剂, 孵育2.5h后用酶标仪测定各孔450 nm处吸光度(A值); 用IRE1α抑制剂STF-083010 (50 μmol/L)[19]与20 mg/L NaNO2共同处理草鱼肝细胞24h, 每组4个平行。在培养结束后, 每孔加入10 μL CCK8试剂, 孵育2.5h后用酶标仪测定各孔450 nm处吸光度(A值)。
1.4 Annexin-FITC /PI 双染流式细胞术检测细胞凋亡
将细胞接种于六孔板中, 待细胞汇合达80%—90%时, 将培养基分别换为不同浓度的亚硝酸钠(0、5、20 和50 mg/L) M199培养基分别孵育12h和24h, 用胰蛋白酶消化, 制成细胞悬液, 离心取细胞沉淀, 用PBS洗涤, 4℃ 300×g离心5min, 2次, 弃上清液, 将细胞重悬浮于100 μL Binding Buffer。加入5 μL Annexin-FITC和10 μL PI, 轻轻混匀, 避光室温反应10min, 加入400 μL Binding Buffer, 1h内进行检测; 将细胞接种于六孔板中, 待细胞汇合达80%—90%时, 用IRE1α抑制剂STF-083010(50 μmol/L)与20 mg/L亚硝酸钠共同处理草鱼肝细胞24h, 用胰蛋白酶消化, 制成细胞悬液, 离心取细胞沉淀, 用PBS洗涤, 4℃ 300×g离心5min, 2次, 弃上清液, 将细胞重悬浮于100 μL Binding Buffer。加入5 μL Annexin-FITC和10 μL PI, 轻轻混匀, 避光室温反应10min, 加入400 μL Binding Buffer, 1h内进行检测。每个处理组取200 μL加入培养皿中, 在荧光显微镜下观察细胞凋亡。
1.5 反转录聚合酶反应(RT-RCR) 检测mRNA表达量变化
取对数生长期L8824, 接种于6孔板。每孔2 mL培养液, 每组3个平行。按照上述处理方法, 对细胞进行处理, 培养完成后收集各组细胞。Trizol法提取细胞总RNA, 测定其浓度、纯度并按试剂盒说明书步骤反转录cDNA。引物为擎科生物公司设计合成, 引物名称序列扩增片段长度见表1。qPCR反应配置为蒸馏水7.2 μL, SYBR Green qPCR Master Mix 10 μL, 上游引物(10 μmol/L) 0.4 μL, 下游引物(10 μmol/L) 0.4 μL, 样品cDNA溶液2 μL, 总体积20 μL。

表1 引物序列Tab. 1 Primers used in the study
1.6 荧光显微镜观察细胞内钙离子浓度变化
取生长状态良好的对数生长期的L8824细胞接种于六孔板中, 接种浓度约为1×106个细胞/孔, 依照上述处理方法, 对细胞进行处理。在培养结束后,除去培养基。用Hank’s平衡盐溶液(HBSS)溶液洗涤3次, 加入FLuo-4 AM工作液(5 μmol/L), 37℃培养箱孵育30min, 除去FLuo-4 AM工作液, 用HBSS溶液洗涤细胞3次, 加入HBSS溶液覆盖细胞, 37℃培养箱孵育30min, 用荧光显微镜检测。
1.7 数据分析
各处理组设置3个重复, 数据以“平均数±标准差”, 采用SPSS19.0软件进行单因素方差分析, 差异显著表示为P<0.05。
2 结果
2.1 亚硝酸钠对L8824细胞活力的影响
在亚硝酸钠暴露12h后, 在暴露浓度为5 mg/L时, 细胞活力下降并不显著, 此后随着暴露浓度的增加, 与对照相比, 细胞活力均显著性下降, 亚硝酸钠暴露24h后, 随着暴露浓度的增加, 与对照相比,细胞活力均显著性下降, 暴露浓度为50 mg/L时细胞活力下降最为显著(表2)。
2.2 亚硝酸钠诱导L8824细胞凋亡
在亚硝酸钠暴露12h后,jnk、bax、caspase9、caspase3表达量在最高浓度50 mg/L时均显著上升,bcl-2表达量显著下降。亚硝酸钠暴露24h后,jnk表达量在20 mg/L时显著性升高,bax、caspase9、caspase3表达量在20和50 mg/L时均显著性上升, 而bcl-2表达量在20 mg/L时显著下降, 其中,bax表达量在20 mg/L时达到最大值, 而caspase9、caspase3表达量在50 mg/L时达到最大值。
由图1可见, 亚硝酸钠暴露浓度为20 mg/L时,随着暴露时间的增加,jnk、caspase9、caspase3表达量均显著上升, 而bcl-2表达量显著下降。亚硝酸钠暴露浓度为50 mg/L时, 随着暴露时间的增加,bax、caspase9 和caspase3表达量均显著性上升。
在亚硝酸钠暴露12h和24h后, 与对照相比, 暴露浓度在5 mg/L时L8824 细胞凋亡率没有显著性变化, 在高浓度20和50 mg/L暴露时, L8824 细胞凋亡率均显著性上升。在亚硝酸钠暴露浓度为20和50 mg/L时, 随着时间的增加, L8824细胞凋亡率均显著性上升(图2)。
2.3 亚硝酸钠诱导草鱼肝细胞L8824内质网应激
亚硝酸钠暴露12h后,ire1α、xbp1s、grp78表达量在20和50 mg/L时均显著性上升, 其中,ire1α表达量在50 mg/L时达到最大值, 而xbp1s、grp78表达量在20 mg/L时达到最大值。在亚硝酸钠暴露24h后,ire1α、xbp1s和grp78表达量在5、20和50 mg/L时均显著性上升, 其中ire1α和grp78表达量在50 mg/L时达到最大值, 而xbp1s表达量在20 mg/L时达到最大值(图3)。
2.4 亚硝酸钠诱导细胞内钙离子紊乱
由图4可见, 亚硝酸钠暴露12h和24h后, 对照组无明显绿色荧光, 亚硝酸钠浓度从5到50 mg/L, 细胞内绿色荧光开始变多且趋于明亮, 钙离子荧光强度相对于对照均显著性上升。亚硝酸钠暴露相同浓度时, 随着时间的增加, 细胞内绿色荧光开始变多且趋于明亮, 钙离子荧光强度相对于对照均显著性上升, 呈现浓度与时间依赖性。

表2 不同浓度亚硝酸钠对L8824 细胞活力的影响Tab. 2 The effect of sodium nitrite on the viability of L8824 cells
2.5 IRE1α抑制剂STF-083010通过抑制IRE1通路减少亚硝酸钠诱导的细胞凋亡
在20 mg/L亚硝酸钠暴露组中加入IRE1α抑制剂STF-083010 (50 μmol/L)作用24h, 与对照相比, STF-083010对L8824细胞活力没有显著性影响, 亚硝酸钠单处理组细胞活力显著下降。与亚硝酸钠单处理组相比, STF-083010处理组细胞活力显著上升(表3)。

图1 亚硝酸钠对L8824细胞jnk、bcl-2、bax、caspase9和caspase3表达量的影响Fig. 1 Effects of sodium nitrite on the expression of jnk, bcl-2,bax, caspase9 and caspase3 in L8824 cells

图2 亚硝酸钠对L8824细胞凋亡的影响Fig. 2 The effect of sodium nitrite on apoptosis of L8824 cells
由图5可见, 与对照相比, 20 mg/L亚硝酸钠单处理组中ire1α、xbp1s、grp78、jnk、bax、caspase9和caspase3的表达显著上升,bcl-2表达量显著下降。与亚硝酸钠单处理组相比, STF-083010处理组ire1α、jnk、bax和caspase3的表达显著下降,bcl-2表达量显著上升,caspase9表达量无显著性变化。与对照相比, 20 mg/L亚硝酸钠处理组凋亡率显著上升; 与亚硝酸钠单处理组相比, STF-083010处理组细胞凋亡率显著下降(图6A)。在荧光显微镜下观察, 与对照相比, 亚硝酸钠处理组荧光强度显著增强; 与亚硝酸钠单处理组相比, STF-083010处理组荧光强度显著降低(图6B)。
2.6 2-APB和STF-083010减弱亚硝酸钠诱导的细胞内钙离子紊乱
与对照相比, 20 mg/L亚硝酸钠处理组绿色荧光变多且强度显著增强, 与亚硝酸钠单处理组相比,2-APB组和STF-083010组绿色荧光变少且强度显著减弱(图7)。
3 讨论
3.1 亚硝酸盐诱导L8824细胞发生凋亡

图3 亚硝酸钠对L8824细胞ire1α, xbp1s和grp78表达量的影响Fig. 3 Effects of sodium nitrite on the expression of ire1α, xbp1s and grp78 in L8824 cells

图4 亚硝酸钠对L8824细胞内钙离子浓度的影响Fig. 4 The effect of sodium nitrite on the intracellular calcium content in L8824 cells
亚硝酸盐作为氨转化为硝酸盐过程中的中间产物, 其对鱼、虾和蟹等水生生物均具有较强的毒性作用, 是养殖水域中诱发暴发性疾病的重要因素。关于亚硝酸盐对鱼类的致毒机理, 国内外已在多种鱼类中开展了相关研究[20,21]。亚硝酸盐进入水生动物的体内主要通过鳃的吸收作用, 在淡水鱼和甲壳动物中, 其体液离子浓度相对于外界水环境处于高渗状态, 因此必须通过鳃的主动吸收以弥补机体离子的流失[22]。亚硝酸盐进入水生生物体内主要通过离子交换机制。在淡水鱼类的鳃上皮上面存在一种Cl-/离子交换机制, 一般在碳酸酶(CA)作用下。H2CO3被水解为H+和, 而H+-ATPase主动运输为排出胞外的浓度梯度。在Cl-/离子交换机制作用下,得以排出细胞膜外而环境中存在的Cl-被主动吸收进入鳃上皮细胞[23]。而当水生生物处于较高浓度的亚硝酸盐暴露下, Cl-/离子交换机制受到影响, 一部分Cl-吸收能力被的吸收代替, 从而造成了水生生物体内亚硝酸盐的堆积。亚硝酸盐通过阴离子交换机制进入鳃上皮以后, 在细胞外液中往往能够积累到很高的浓度[24]。而随着亚硝酸钠不断进行积累, 其导致血液中高铁血红蛋白的毒性越发的明显, 失去部分氧气运输的能力造成鱼体最终窒息死亡[25]。此外, 亚硝酸盐暴露往往能够引起鱼体的氧化应激, 因此鱼类中抗氧化酶活性水平的变化往往作为不同水生生物应激反应的生物标志物[26]。在对罗非鱼(Tilapia nilotica)的研究中发现,当水环境中的亚硝酸盐浓度高于罗非鱼自身的耐受范围时, 其丙二醛的含量随着氧自由基积累而增加[27]。而鱼体内为了防止氧自由基的积累, 常常依赖于由超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶等抗氧化酶类组成的抗氧化系统的作用, 而长期暴露可能会对抗氧化系统产生一定的抑制作用。鱼类肝脏在调控基础代谢、降解与排泄外源有毒物质中发挥重要作用, 并且可以作为鱼体健康状况的指示性参数, 肝脏作为亚硝酸钠的主要解毒器官之一, 肝细胞可以将亚硝酸盐转化为硝酸盐以达到解毒目的[28]。简而言之, 亚硝酸盐作为强氧化剂, 进入机体后会削弱其血红细胞运氧能力,并引起自由基增多、生理紊乱、组织缺氧, 体内多种生理机能的障碍和下降, 产生氧化应激、扰乱内分泌、造成组织损伤、引起细胞凋亡, 从而影响正常的生理机能[29,30]。目前有关于亚硝酸钠对肝脏细胞凋亡的研究还比较局限, 亚硝酸钠诱导肝脏细胞凋亡可能存在的分子机制还有待阐明。本实验结果发现亚硝酸钠暴露下凋亡相关基因,jnk、bax、caspase9、caspase3表达量显著性上升, 亚硝酸钠暴露12h和24h后L8824细胞凋亡率显著上升。以上结果表明, 急性亚硝酸钠暴露能导致草鱼肝细胞凋亡。Al-Gayyar 等[31]在研究黑藻油对亚硝酸钠引起的大鼠细胞凋亡的影响中发现亚硝酸钠引起大鼠肾组织损伤, 并引起了大鼠肾组织发生细胞凋亡;Al-Rasheed 等[32]研究槲皮素对亚硝酸钠所致缺氧大鼠肝、肺、肾、心肌组织的保护作用中, 发现亚硝酸钠会下调大鼠组织中bcl-2表达量, 诱导发生细胞凋亡, 与本实验结果相一致。

表3 亚硝酸钠和STF-083010共同作用对L8824细胞活力的影响Tab. 3 Effects of sodium nitrite and STF-083010 on the viability of L8824 cells
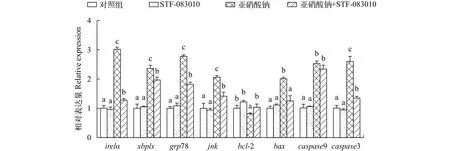
图5 亚硝酸钠和STF-083010及其配伍对L8824细胞ire1α、xbp1s、grp78、jnk、bcl-2、bax、caspase9和caspase3表达量的影响Fig. 5 Effects of sodium nitrite, STF-083010 and their combination on the expression of ire1α, xbp1s, grp78, jnk, bcl-2, bax, caspase9 and caspase3 in L8824 cells
3.2 亚硝酸盐诱导L8824细胞发生内质网应激
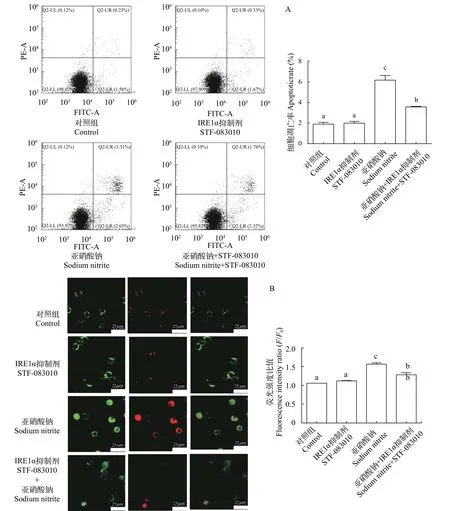
图6 亚硝酸钠和STF-083010及其配伍对L8824细胞凋亡的影响Fig. 6 Effects of sodium nitrite, STF-083010 and their combination on the cell apoptosis of L8824 cells
UPR是由一个内质网分子伴侣GRP78和3个内质网应激感受蛋白所介导的, 分别是PERK (PKR-like ER kinase)、ATF6 (Activating transcription factor 6)和IRE1。IRE1被认为是UPR信号中最后一个被激活的分子, IRE1一旦被激活, 首先通过剪接XBP1诱导UPR, 随后诱导P58IPK的表达恢复蛋白质合成, 使细胞恢复到正常状态。但如果应激继续加重, IRE1 就会通过激活JNK激酶, 削弱Bcl-2的抗凋亡功能, 从而诱导内质网膜上Bax和Bak构象变化并寡聚化最终导致内质网膜完整性的破坏和细胞内钙离子浓度增加, 激活Caspase9 最终激活Caspase3 诱导细胞凋亡[33]。在本实验中ire1α和grp78表达量在高浓度亚硝酸钠暴露12h和24h后显著性上升, 但xbp1s在亚硝酸钠暴露24h后表达量显著下降, 这表明随着暴露时间的增加, UPR中IRE1-XBP1通路被抑制, 原因可能是因为随着暴露时间的增加,IRE1减少对XBP1的剪接作用, 内质网应激减弱,IRE1激活JNK, 诱导细胞凋亡。Dickhout 等[34]通过检测到GRP78和GPR94的蛋白表达量上调发现过氧化亚硝酸盐在人血管内皮引起了内质网应激。同时, 荧光显微镜检测细胞内钙离子浓度发现, 不同浓度和不同时间的亚硝酸钠暴露都引起了不同程度的钙离子紊乱, 内质网分子伴侣在内质网应激中发挥作用需要钙离子的参与, 因此钙离子紊乱会诱导发生内质网应激[35]。而在本实验中内质网分子伴侣grp78 mRNA 表达量在亚硝酸钠暴露后显著上升支持了这个结论, GRP78是钙离子的缓冲伴侣, 能够影响内质网依赖的钙离子稳态[36,37], 亚硝酸钠通过引起内质网释放钙离子诱导细胞内钙离子水平升高, 阻碍了内质网折叠蛋白的功能, 从而诱导细胞发生内质网应激[38]。以上结果表明, 亚硝酸钠能够通过激活IRE1通路引起草鱼L8824细胞发生内质网应激。
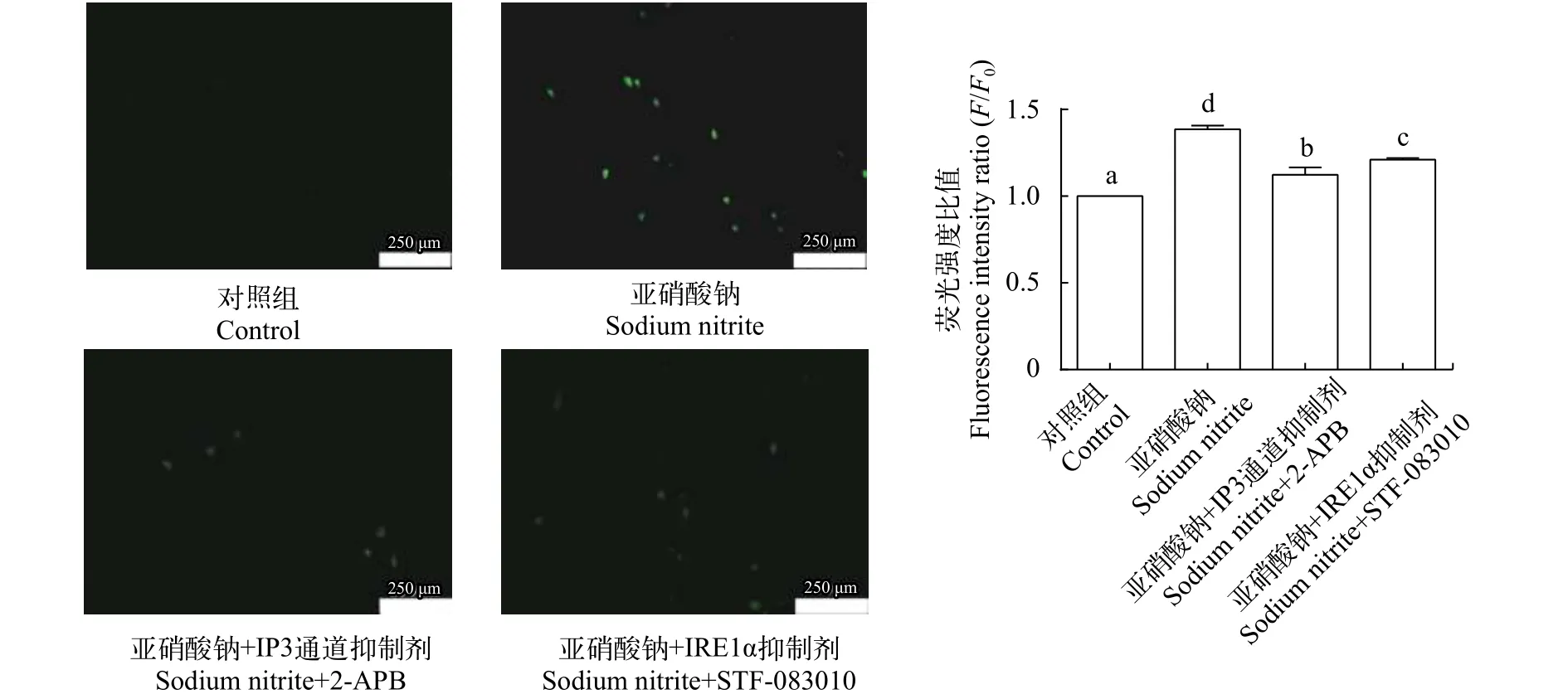
图7 2-APB和STF-083010 对亚硝酸钠暴露L8824细胞内钙离子浓度的影响Fig. 7 Effects of 2-APB and STF-083010 on the intracellular calcium content under sodium nitrite exposure in L8824 cells
3.3 IREI通路在亚硝酸盐诱导L8824细胞发生凋亡中发挥重要作用
近年来大量研究表明内质网应激是诱导细胞凋亡的重要途径[39], 为研究IRE1 通路在亚硝酸钠诱导L8824细胞发生凋亡的作用, 本实验用IRE1α抑制剂STF-083010与20 mg/L亚硝酸钠共同作用于L8824细胞24h, 结果发现, 在IRE1被抑制后, 细胞凋亡率显著下降,jnk、bax、caspase3表达量也显著下降, 而抗凋亡分子bcl-2表达量显著上升, 说明JNK诱导的细胞凋亡也被抑制。同样地, Kato 等[40]发现在深部组织损伤的大鼠肾小管上皮细胞中, 抑制IRE1通路可显著降低细胞凋亡; Huang 等[41]研究发现乙型脑炎病毒诱导BHK-21细胞凋亡实验中, 抑制IRE1通路也可显著降低BHK-21细胞凋亡。此外, 在本实验中荧光显微镜检测细胞内钙离子浓度结果显示, 加入STF-083010显著减弱了钙离子紊乱, 减少了内质网应激[38]。由此可见, 亚硝酸盐和其他的致凋亡因子相似, 可以通过抑制IRE1通路降低其诱导的细胞凋亡。
综上所述, 高浓度亚硝酸钠会诱导L8824细胞发生凋亡, 激活内质网应激IRE1通路, 显著降低细胞活力, 并引起细胞质内钙离子紊乱; 加入了IRE1抑制剂后, 细胞凋亡和细胞质内钙离子浓度显著降低, 细胞活力显著升高。以上结果表明, 内质网应激IRE1通路在高浓度亚硝酸钠诱导的L8824细胞凋亡中发挥了重要作用。
