重组新城疫病毒 IL29 抑制人胃癌细胞系BGC的增殖与迁移
2020-03-04尹超云陈振威张安伟张旋风张日亭步雪峰
尹超云, 张 垚,陈振威,张安伟,张旋风,张日亭,步雪峰
(1.江苏大学附属医院 血管外科, 江苏 镇江 212000; 2.江苏大学 医学院,江苏 镇江 212013; 3.江苏大学附属人民医院 普外科, 江苏 镇江 212002)
胃癌(gastric cancer)是中国常见的消化道肿瘤之一,居于第2位[1]。肿瘤的转移是影响肿瘤预后重要的原因之一,而肿瘤细胞的增殖与迁移能力是肿瘤细胞转移的关键环节。IFN λ 家族是一组新型人白细胞介素(interleukin,IL),又名Ⅲ型 IFN,包 括 IL-29 (IFN-λ1)、IL-28A(IFN-λ2) 和 IL-28B(IFN-λ3),具有抗增殖、抗肿瘤以及免疫调节等生物学活性[2]。有研究表明,表达IL-29 的重组新城疫病毒LoSota 株(recombinant Newcastle disease virus LoSota strain expressing IL-29, rL-IL29) 抑制肺腺癌A549 细胞的增殖及迁移与AKT途径有关[3],对于IL-29 是否能抑制胃癌细胞增殖与迁移尚不清楚。在多种癌细胞中发现ERK(extracellular signal regulated kinase,ERK)的被异常激活,是肿瘤发生的重要因素之一。研究表明胃癌患者血清中磷酸化ERK蛋白水平升高,且随着胃癌分期的增加其表达量也增加[4]。同时ERK与AKT在肿瘤转移的通路中具有重要的作用。本实验探讨IL-29对胃癌BGC细胞增殖和迁移的影响及其相关机制。
1 材料与方法:
1.1 材料
新城疫病毒(Newcastle disease virus,NDV) (哈尔滨兽医研究所);rL-IL29(本实验组构建);人胃癌细胞系BGC(中国科学院上海生科院细胞资源中心);RPMI 1640培养基(Gibco公司),胎牛血清(维森特生物技术南京公司);兔抗IL-29、MMP2(72、66 ku)、p-AKT抗体(美国博士德生物公司);兔抗P-ERK1/2、HRP标记的抗鸡(Abbkine公司);HRP标记羊抗兔IgG、羊抗鼠IgG(均上海康为世纪公司);Alexa Flour 488标记羊抗鼠IgG、Cy3标记羊抗鼠IgG(KLP公司);CCK-8试剂盒(南京厚载生物科技公司)。
1.2 方法
1.2.1 细胞的分组及处理:将BGC细胞置于37 ℃、5% CO2的饱和湿度培养箱中,用含10%胎牛血清的RPMI 1640(加100 U/mL青-链霉素)培养。取对数增殖的BGC细胞随机种于6孔板内,培养24 h,待BGC细胞覆盖6孔板孔80%左右时将6孔板随机分为PBS组(即对照组)、rL-IL29组及NDV组。用1640培养基将IL-29及NDV稀释至10 000倍感染6孔板内BGC细胞,培养24 h后为下一步实验做准备,每组样本重复3次。
1.2.2 CCK8法检测BGC细胞活力:在96孔板每孔中加入BGC细胞悬液100 μL (约5 000~10 000个细胞),边缘孔用PBS填充,在恒温培养箱中孵育12 h。将IL-29及NDV病毒稀释倍数为为原液的10-2、10-3、10-4、10-5和10-6,每孔加入上述浓度10 μL的稀释病毒液,培养箱孵育24 h。再在每孔加入10 μL CCK-8溶液,培养箱孵育4 h。设置无胃癌细胞的空白孔(培养基和CCK-8溶液),有胃癌细胞的对照孔(不加培养基和CCK-8溶液),每组设定3个复孔。测定450 nm各孔的吸光度值。细胞活力=(实验组a-空白组a)/(对照组a-空白组a)
1.2.3 病毒滴度测定: 将细胞增殖状态良好的BGC 细胞消化计数后稀释至1×105个/mL,加入96孔板,100 μL/孔(37 ℃,5% CO2),培养 24 h 后加病毒。在1.5 mL离心管中,设置6个稀释度,第一管加90 μL培养液,然后加入10 μL病毒原液,混匀后吸取10 μL加入第2个离心管中,以此类推,培养24 h后观察其死亡(或阳性)数,然后计算出各稀释点引起死亡累积数的死亡率。所求得的值就写为EID50。其算法为:lgEID50=高于 50%死亡的稀释倍数的对数+稀释系数的对数×距离比距离比=(高于50%的死亡率-50%) /(高于50%的死亡率-低于50%的死亡率)
1.2.4 克隆形成实验检测细胞增殖:将6孔板孔分为对照组、IL-29 组及NDV 组细胞,按每孔1 500 个细胞的标准接种对数增殖的BGC细胞, 每组设立3 个复孔。培养24 h 后予以换液,每隔2~3 d 换1 次培养基,培养10 d后,结晶紫染液进行染色,计数并拍照。按公式计算: 克隆形成率(%)=(克隆数/接种细胞数)×100%(以50个细胞以上为一个集落)。
1.2.5 Western blot检测IL-29、NDV、MMP2、p-ERK和p-AKT蛋白:将1.2.1中准备好的6孔板里的细胞提取蛋白。进行SDS-PAGE电泳,再转膜,5% BSA封闭1 h,分别加入鸡抗NDV蛋白抗体(1∶800)、兔抗IL-29、MMP2、p-AKT(1∶300)、兔抗P-ERK(1∶400)及β-肌动蛋白抗体(1∶2 000),4 ℃孵育过夜。加入羊抗鼠或羊抗兔或羊抗鸡 HRP-IgG(1∶2 000)室温孵育1 h,TBST洗膜,ECL显色扫描,LANE.1D软件进行条带吸光度值定量分析。
1.2.6 免疫荧光检测p-ERK的表达: 24孔板内对数增殖的BGC细胞经病毒处理培养24 h 后,在室温下经4%多聚甲醛固定4 h,PBS洗3次,0.5% Triton×100室温下处理细胞30 min,PBS洗3次,经5% BSA封闭1 h,p-ERK一抗4 ℃孵育过夜。PBS洗3次,Cy3标记的兔二抗(1:200)染色1 h,PBS洗3次;Hoechst 33342 染色细胞核10 min,PBS洗3次后置于荧光显微镜下观察。
1.2.7 Transwell小室法检测细胞侵袭:用无血清 DMEM 配制成约1×105个/mL的BGC细胞悬液,取 200 μL滴入24孔板Transwell小室的上室,下室分别加入含10%胎牛血清的1640培养基稀释的PBS(对照组)、NDV及rL-IL29(1∶1 000)各500 μL,每组均设立2个复孔,37 ℃、5% CO2的培养箱内培育 24 h。用棉签擦去 Transwell 上室聚碳酸酯膜表面的细胞,PBS洗2遍,将上室置于4%多聚甲醛中固定15min。PBS洗2遍后,结晶紫染色20 min,再用PBS洗2遍后,倒置相差显微镜下计数10个不同视野的细胞数,计算平均值。
1.2.8 划痕实验检测细胞迁移:24孔板中的BGC细胞用含10%胎牛血清的1640 培养基进行培育,待细胞覆盖率达80% ~ 90% 时,用无菌10 μL 的加样枪头在细胞层中缓缓的纵向划一条直线,同时保持划痕的宽度均匀一致; 再用PBS 洗3遍,用1640培养基将病毒rL-IL29、NDV分别稀释到10 000倍,将稀释后rL-IL29、NDV病毒稀释液加入到24孔板中作为rL-IL29组和NDV组,同时用1640培养基作为对照组。分别于划痕后0、24 h 在倒置显微镜下观察划痕间距。
1.3 统计学分析
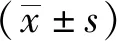
2 结果
2.1 CCK8检测病毒对BGC活力的影响
随着病毒的稀释倍数降低时,胃癌细胞的活力降低,当病毒稀释液浓度为10-2、10-3时细胞活力<80%。选用细胞活力>80%时的滴度[5],故后续实验病毒稀释液选择10-4作为后续实验的工作浓度,此时La Sota系NDV病毒液及rL-IL29 病毒液的滴度都在鸡胚半数感染量 (50% egg infective doses,EID50) 为109.8 EID 50 /mL左右(P<0.05)(图1)。
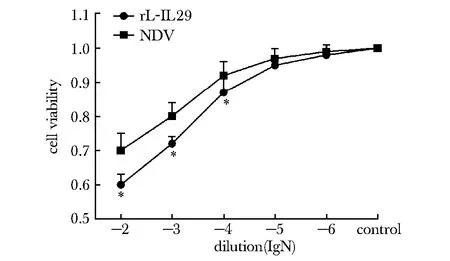
*P<0.05 compared with the NDV group图1 不同浓度的rL-IL29及NDV病毒对BGC胃癌细胞活力的影响Fig 1 Effects of different concentrations of rL-IL29 and NDV viruses on the cell vitality in gastric cell
2.2 克隆形成实验检测rL-IL29感染对BGC细胞增殖的影响
将两种稀释后的病毒感染胃癌细胞BGC培养10 d 后,rL-IL29、NDV病毒抑制了胃癌细胞BGC的增殖,同时rL-IL29组的胃癌细胞较NDV及PBS组受到明显的抑制 (P<0.05)(图2)。
2.3 NDV、IL-29、p-ERK、MMP2和pAKT蛋白的表达
NDV蛋白在rL-IL29 组及NDV组稳定表达,rL-IL29组的 IL-29 蛋白表达较NDV 组及PBS组明显升高(P<0.05),p-ERK蛋白在rL-IL29组、NDV组的相对表达量较PBS组明显降低(P<0.05),且rL-IL29组的p-ERK蛋白表达量低于NDV组(P<0.05);MMP2及pAKT得到了相似的结论(图3)。
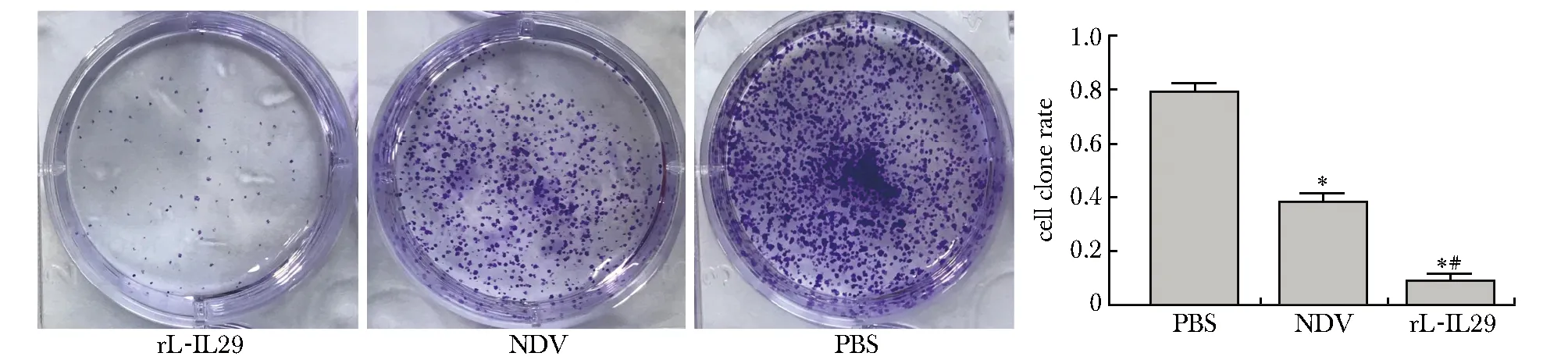
*P<0.05 compared with the PBS group;#P<0.05 compared with the NDV group图2 rL-IL29 感染对BGC细胞增殖的影响Fig 2 Effect of rL-IL29 infection on proliferation in gastric cell line
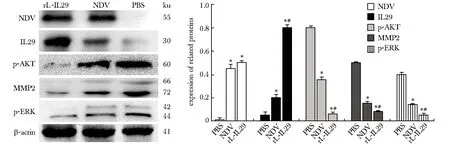
*P<0.05 compared with the PBS group;#P<0.05 compared with the NDV group图3 感染 rL-IL29 后对 BGC 细胞迁移相关蛋白的影响Fig 3 Effect of rL-IL29 on the migration-related protein in gastric cell line
2.4 P-ERK的免疫荧光检测
P-ERK蛋白在rL-IL29组的蛋白表达量明显低于NDV组及PBS组(P<0.05),p-ERK蛋白在NDV组的蛋白表达量明显低于PBS组(P<0.05)(图4)。
2.5 rL-IL29 抑制 BGC 胃癌细胞侵袭能力:
rL-IL29 组穿过Transwell 小室的胃癌细胞数明显少于 NDV 组及 PBS组(P<0.05),NDV 组的胃癌细胞穿过 Transwell 小室的细胞数明显少于 PBS 组(P<0.05) (图5)。
2.6 rL-IL29 抑制BGC细胞的迁移能力
48 h 后rL-IL29 组 BGC 细胞的迁移距离明显低于 NDV 组和 PBS 组,NDV 组 BGC 细胞的迁移距离明显低于 PBS组 (P<0.05)(图6)。
3 讨论
新城疫病毒(NDV) 具有很好的溶瘤作用且对人的致病性极低,可以促进 NK 细胞产生IFN-γ从而上调 NK 细胞 TRAIL 蛋白表达,致使肿瘤细胞的凋亡与坏死[6]。 IL-29 在人体多种细胞中均有表达,同时它的活性最强, IL-29 通过与 IL-28R1 结合发挥生物学效应[7]。IFN λ 可以明显抑制鼠的皮下瘤肺转移的作用[8]。本实验结果表明,在rL-IL29转染BGC胃癌细胞24H后,IL-29 蛋白表达明显高于其他组,可以表明rL-IL29稳定表达IL-29。 同时从细胞克隆实验、细胞划痕实验及Transwel可以看出rL-IL29明显抑制了胃癌细胞的增殖与迁移作用,且 rL-IL29的抑制效果较 NDV更明显。
ERK是一类丝/苏氨酸激酶,在细胞的增殖、迁移、分化和凋亡等多种过程发挥了重要作用,当受到细胞外信号刺激时,可激活ERK信号传导通路,参与了细胞外信号传入细胞内的经典途径[9]。有研究表明异常激活的磷酸化的ERK在胃癌、肺癌等多种恶性肿瘤中均能发现,同时ERK通路也涉及恶性肿瘤的发生、发展及恶化过程[10]。本研究中rL-IL29组ERK蛋白磷酸化水平(p-ERK)较NDV组及PBS组下调,说明rL-IL29可以下调p-ERK从而抑制了ERK的信号传导通路。基质金属蛋白酶(matrix metalloproteinases,MMPs) 是一类细胞外蛋白水解酶,可以将细胞外基质中的各种蛋白成分降解[11]。MMP2 作为其家族中的一员,在基底膜的降解以及肿瘤细胞迁移、浸润中起着至关重要的作用[12],MMP2 表达强度与胃癌分期、淋巴结转移呈正相关[13]。AKT 是 PI3K/AKT信号通路中的原癌基因,PDK1可以激活PI3K/AKT, 活化的 AKT可以调节其下游的多条通路,从而促进细胞增殖、抗凋亡以及迁移[14]。本实验中发现rL-IL29可以下调MMP2和p-AKT的水平,从而进一步证明rL-IL29可以抑制BGC胃癌细胞的迁移,同时也表明rL-IL29的抑制肿瘤细胞的机制与下调MMP2和p-AKT蛋白有关。
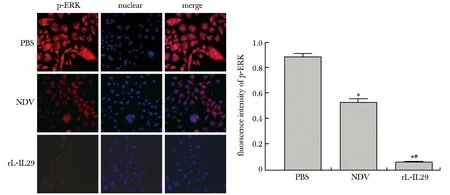
*P<0.05 compared with the PBS group;#P<0.05 compared with the NDV group图4 感染 rL-IL29 后对 BGC 细胞p-ERK蛋白的影响Fig 4 Effect of rL-IL29 on p-erk protein in gastric cell line
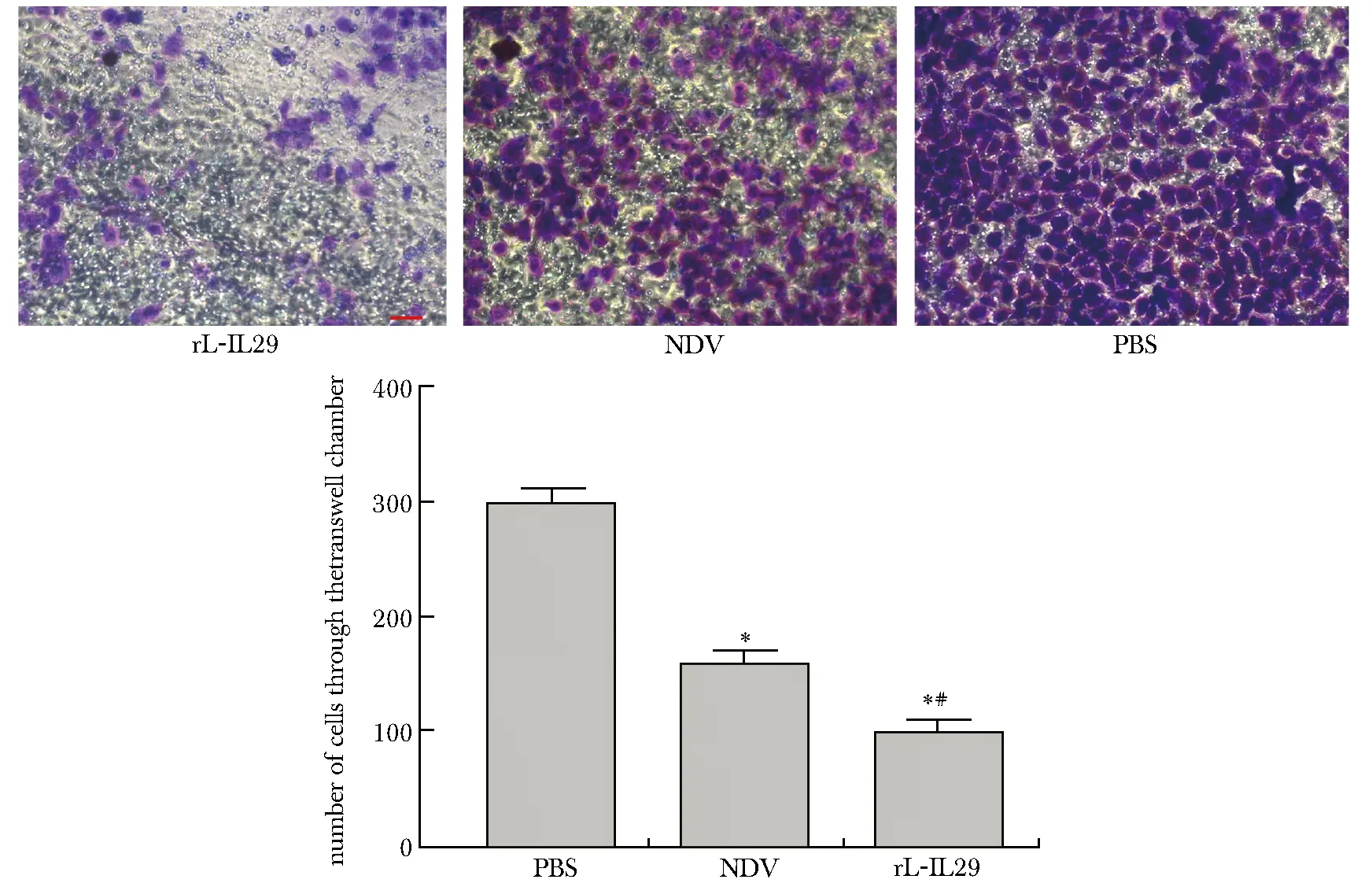
*P<0.05 compared with the PBS group;#P<0.05 compared with the NDV group图5 感染 rL-IL29 后对 BGC 细胞侵袭性的影响Fig 5 Effect of rL-IL29 infection on the invasiveness in gastric cell line

*P<0.05 compared with the PBS group;#P<0.05 compared with the NDV group图6 感染 rL-IL29 后对 BGC 细胞迁移的影响Fig 6 Effect of rL-IL29 infection on migration in gastric cell line
综上,本研究表明 rL-IL29 可能通过阻碍ERK, PI3K /Akt信号通路,下调MMP2蛋白的表达,从而抑制 BGC胃癌细胞的增殖与迁移,但两条通路的具体的作用机制仍需进一步研究。
