一株近玫色锁掷酵母的分离鉴定及其产胞外乳糖酶性质的研究
2020-02-25苑广伟李镇标孙玉红王兆磊
苑广伟, 林 梅, 李镇标, 孙玉红, 王兆磊, 王 磊*
1.华南农业大学材料与能源学院,广州 510642;2.广东唯新生物科技有限公司,惠州 516057;3.深圳唯新生物科技有限公司,深圳 518116
近玫色锁掷酵母(Sporidioboluspararoseus)是一类担子菌门真菌,属掷孢酵母科,菌落一般呈红色或浅红色。近年来研究发现,锁掷酵母能产色素、不饱和脂肪酸、多糖等营养代谢物质,已作为食品添加剂应用于动物饲料中,其在医药、环保等领域也越来越受到人们关注,应用范围也愈加广泛[1]。目前利用其生产天然类胡萝卜素的研究日趋火热,LIU C等人通过酸碱处理法、酶溶法、化学试剂处理法、高压匀浆法和细胞自溶法五种破壁方法处理近玫色锁掷酵母,达到破坏细胞壁释放类胡萝卜素的目的,并通过SK-HEP-1细胞实验确定其产生的类胡萝卜素具有抗氧化作用[2];CHAIYASO T等人研究发现SporidioboluspararoseusKX709872具有产α-淀粉酶和葡糖糖苷酶的活性,将其用于生物发酵,结果表明其有效的将餐厨垃圾转化为生物质和脂类,达到变废为宝的作用[3];DU C等人的研究表明Sporidioboluspararoseus的代谢产物可缓解血脂异常的情况,可用于改善肥胖人员的脂类代谢,降低疾病的发生[4],但针对近玫色锁掷酵母(Sporidioboluspararoseus)所产乳糖酶的研究还未深入。
乳糖酶(Lactase)又称β-D-半乳糖苷半乳糖水解酶(β-D-galactoside-galactohydrolase,EC.3.2.1.23),简称为β-半乳糖苷酶(β-galactosidase)[5,6]。乳糖酶能催化乳糖水解生成葡萄糖和半乳糖[7]。β-半乳糖苷酶在自然界中广泛地存在,哺乳动物的肠、大脑,特别是婴儿的肠道中都含有β-半乳糖苷酶[8,9];桃、苹果和咖啡豆等植物中也含有β-半乳糖苷酶[10];微生物界中的细菌、酵母、霉菌和放线菌等都具有产β-半乳糖苷酶的能力[11]。β-半乳糖苷酶可用于乳产品的生产加工[12],β-半乳糖苷酶通过水解乳糖生产低聚半乳糖,而低聚半乳糖是一种功能性物质,是人体肠道中胃肠道微生物菌落所利用的营养源。低聚半乳糖有利于维持和恢复胃肠道菌群,同时促进其生长增殖,其具有调节肠道健康,改善免疫功能、降低代谢紊乱的潜力[13];β-半乳糖苷酶作为酶制剂可用于治疗乳糖不耐受症状及多种儿童消化不良症,特别是婴幼儿腹泻病[14];β-半乳糖苷酶还能与其他抗生素联用,例如与杜迪和思密达联用治疗轮状病毒肠炎,能明显地缩短治疗疗程,提高治愈度;同时β-半乳糖苷酶还可以用于免疫分析、对微量激素药物的定量分析,以及病毒等的测定[15];β-半乳糖苷酶的水解从而有效解决乳清的利用和排放问题,同时还生成乳清糖浆等制品,从而达到节约资源,减少环境污染的可能[16]。
近年来,微生物来源的β-半乳糖苷酶被广泛研究,其在食品和制药工业中的许多应用而受到越来越多的关注;酵母菌是生产β-半乳糖苷酶的主要微生物菌株[17]。乳酸克鲁维酵母(Kluyveromyceslactis)作为生产乳糖酶的优先来源已被广泛研究;SONG C等人对嗜冷性酵母产生的β-半乳糖苷酶进行研究,表明了其应用于乳业生产中的潜力[5];NAKAGAWA等人从日本北海道的土壤样品中分离出一株假单胞菌可以产生耐低温耐酸的β-半乳糖苷酶[18]。在本研究中,从土壤中分离得到一株近玫色锁掷酵母XWSP1(SporidioboluspararoseusXWSP1),并发现其具有产β-半乳糖苷酶的能力,故初步优化其产β-半乳糖苷酶的培养条件和酶学性质,为后续研究提供参考。
1 材料与方法
1.1 材料
1.1.1菌株来源
SporidioboluspararoseusXWSP1,分离自广州徐闻农垦丰收农场土壤。
1.1.2主要试剂和仪器
Tryptone,OXOID公司),Yeast extract,OXOID公司;β-半乳糖苷(ONPG)生化鉴定管,广州环凯微生物科技有限公司;邻硝基苯基-β-D-吡喃半乳糖苷,麦克林;E.Z.N.A.TMSP Fungal DNA Kit,OMEGA BIO-TEK;TaKaRa LA Taq®,TaKaRa公司;10×Loading Buffer,TaKaRa公司;DL 2 000 bp DNA Marker,TaKaRa公司;DL 15 000 bp DNA Marker,TaKaRa公司。超低温冷冻离心机,德国Eppendorf公司;紫外可见分光光度计,尤尼柯仪器有限公司;pH计,奥豪斯仪器有限公司;PCR仪,苏州东胜兴业科学仪器有限公司;核酸测序仪,美国Applied Biosystems公司;电泳仪,美国Bio-rad公司;核酸成像系统,美国UVP公司。
1.1.3种子培养基
马铃薯葡萄糖琼脂培养基(potato peptone agar,PDA,g/L):马铃薯200 g,葡萄糖20 g,NaCl 3 g,KH2PO41.5 g,维生素B10.008 g,琼脂粉15 g。
1.1.4发酵培养基
YPD液体培养基(Yeast Extract Peptone Dextrose Medium,g/L):Tryptone 20 g,Yeast extract 15 g,葡糖糖20 g。
1.2 产乳糖酶菌株的分离、筛选
采用种子培养基进行土壤真菌的分离纯化,取10 g土壤样品,添加90 mL无菌水,置于恒温摇床上28 ℃、180r/min恒温振荡1 h,使土样均匀地分散于无菌水中,制成10-1土壤悬液。采用稀释涂布平板法和平板划线法进行土壤真菌的分离纯化。取10-1土壤悬液,通过梯度稀释,使悬液稀释至10-3;取100 uL不同稀释梯度的悬液涂布于PDA固体培养基中,每个稀释梯度做三个平行组;将涂布完毕的PDA平板置于生化培养箱中28 ℃恒温培养2 d~4 d,观察培养基上菌落生长情况,并通过平板划线法根据菌落的形态特征挑取不同的单菌落进行划线,从而获得纯化菌株。
将已纯化的菌株以2%接种量、1×106CFU/mL接种浓度接种至发酵培养基中,28 ℃、180 r/min恒温振荡培养30 h获得菌液;将菌液稀释至1×106CFU/mL,取50 μL稀释后菌液加入β-半乳糖苷(ONPG)生化鉴定管中(广州环凯微生物科技有限公司),三个平行组,混匀,28 ℃恒温静置培养48 h,将生化鉴定管颜色变黄色的菌株进行进一步纯化和鉴定。
1.3 菌株分子鉴定
采用E.Z.N.A.TMSP Fungal DNA Kit试剂盒法提取真菌基因组DNA。并以ITS1(TCCGTAGGTGAACCTGCGG)和ITS4(TCCTCCGCTTATTGATATGC)为引物进行PCR扩增,PCR扩增条件为:94 ℃预变性5 min;然后94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min,循环35次;72 ℃延伸10 min。PCR产物经琼脂糖凝胶电泳分析后,交由广州擎科生物科技有限公司进行核酸测序。ITS序列提交至GenBank数据库中进行BLAST比对分析。
1.4 粗酶液的制备
取不同培养条件下的近玫色锁掷酵母发酵液,经12 000 r/min,4 ℃离心5 min,除去菌体,得到的上清液即为粗酶液,用于后续胞外乳糖酶酶活测定。
1.5 乳糖酶酶活力测定方法
采用吉斯特-布洛卡兹法[19],以邻硝基苯基-β-D-吡喃半乳糖苷(ONPG)为底物进行测定,取0.5 mL粗酶液待测液于5 mL离心管中,并加入2.0 mL 10 mmol/L ONPG溶液并震荡均匀,30 ℃水浴中反应10 min后取出,加入0.5 mL 1 mol/L Na2CO3(已预冷)溶液显色,于分光光度计420 nm处比色。以加热灭活的粗酶液同样处理作为空白。在上述条件下,每分钟催化ONPG生成1 μmol/L邻硝基苯酚(ONP)所需的酶量定义为一个酶活力单位(1 U)。用已知浓度的邻硝基苯酚(ONP)溶液绘制标准曲线,根据标准曲线得到待测液中邻硝基苯酚(ONP)浓度。从而计算出胞外乳糖酶酶活力。
酶活力计算公式:U/mL=(V1·N·c)/(V2·t)
注:c:根据标准曲线计算出的ONPG浓度(mmol/L);N:为粗酶液稀释倍数;V1:反应总体积(mL);V2:粗酶液体积(mL);t:反应时间(min)
1.6 发酵条件优化
以培养时间、接种量、培养温度、培养基组分(碳源、氮源)等条件优化菌株产乳糖酶的能力。设置三个发酵培养重复组,以1×106CFU/mL接种浓度、2%接种量接种于发酵培养基中,28 ℃、180r/min培养72 h,分别测定培养6 h、12 h、18 h、24 h、30 h、36 h、42 h、48 h、54 h、60 h、66 h和72 h发酵液的胞外乳糖酶酶活。根据测定结果,选择最优培养时间,探索不同接种量、培养温度对酶活力的影响。在以上优化的基础上,设置不同培养基组分,分别以不同碳源取代原始培养基的葡萄糖,确定最优碳源。然后改变培养基中蛋白胨、酵母粉的量,最终确定最适产酶发酵条件。
1.7 pH、金属离子对乳糖酶酶活的影响
在pH分别为5.0、5.5、6.0、6.5、7.0、7.5和8.0的乳糖酶反应体系中,测定pH对胞外乳糖酶活性的影响。根据测定结果,选择最适pH,在反应体系中分别加入终溶度为1 mmol/L的金属离子K+、Na+、Mg2+、Ca2+、Mn2+、Cu2+和Zn2+,探索不同金属离子对胞外乳糖酶活性的影响。
2 结果与分析
2.1 菌株筛选结果
筛选出一株具有乳糖酶活性的菌株,命名为XWSP1,酶活筛选结果如图1所示,与CK(加入等量ddH2O)相比,该菌株使β-半乳糖苷(ONPG)生化鉴定管中液体颜色变为深黄色,表明该菌株具有产乳糖酶的能力。

注:实验组1、2、3为加入XWSP1发酵液的三个平行实验组,CK为加入ddH2O对照组
图1 菌株XWSP1在ONPG生化管中的颜色变化
2.2 菌株XWSP1形态特征
菌株XWSP1在种子培养基上形态特征如图2所示:菌落呈淡红色,向上凸起,表面光滑、湿润、容易挑取,菌落质地均匀,颜色均一。
图2 XWSP1菌落形态
2.3 菌株XWSP1的分子鉴定
通过试剂盒法提取的该菌基因组DNA片段长度在15 kb以上,符合基因组DNA长度的大小,如图3所示。通过ITS1/ITS4引物对进行PCR扩增,该菌PCR产物电泳条带如图4所示,其序列大小在500 bp~750 bp之间,与ITS序列条带大小相符。

注:M为15 000 bp marker;1为菌株XWSP1基因组DNA

注:M为2 000 bp maker;ITS为菌株XWSP1的PCR条带
图4 ITS PCR产物的琼脂糖凝胶的电泳分析
将ITS序列结果在GenBank数据库中进行BLAST比对分析,该菌株与近玫色锁掷酵母SporidioboluspararoseusstrainAUMC 7791(JQ425362.1)同源性最高,达到99%。故命名为:近玫色锁掷酵母XWSP1(SporidioboluspararoseusXWSP1)。进一步利用MEGA7.0软件Neighbor-joining法构建系统进化树,如图5所示,该菌株与SporidioboluspararoseusstrainAUMC 7791(JQ425362.1)聚为一支,进化距离最近。

图5 XWSP1及同源菌株的ITS序列系统进化分析
2.4 菌株XWSP1产乳糖酶发酵培养基的优化
2.4.1培养时间的优化
由图6可知,菌株XWSP1产胞外乳糖酶酶活力与培养时间有关,随着培养时间的增加,酶活力逐渐增加,当培养至54 h,酶活力最高,达到0.019U/mL,此时菌密度亦达到稳定期;当培养时间多于54 h时,酶活力又逐渐降低,推测引起酶活力下降的原因是菌株发酵所需营养物质缺乏,菌株代谢物积累,导致菌株生长代谢受限;故培养时间54 h为产酶最适的培养时间。

图6 菌株培养时间与胞外乳糖酶酶活力关系图
2.4.2接种量的优化
由图7可知,菌株XWSP1以4.0%接种量进行液体培养时,胞外乳糖酶酶活力最高,达到0.050 U/mL当接种量低于或高于4.0%时,酶活力均有所下降。故4.0%的接种量是该菌产酶最适的接种量。
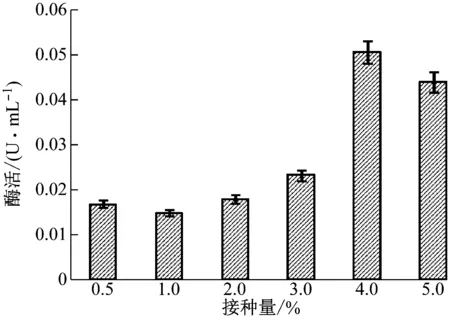
图7 菌株接种量对胞外乳糖酶酶活力的影响
2.4.3培养温度的优化
由图8可知,当菌株XWSP1的培养温度为28 ℃时,其胞外乳糖酶酶活力最高,当培养温度低于23 ℃或高于28 ℃时,其胞外乳糖酶的酶活力明显下降。故28 ℃是该菌发酵及产胞外乳糖酶的最适培养温度。

图8 培养温度对胞外乳糖酶酶活力的影响
2.4.4培养基组分的优化
由图9可知,当以半乳糖、乳糖替代葡萄糖提供碳源时,菌株XWSP1发酵后的胞外乳糖酶酶活力明显升高。以半乳糖做碳源其胞外乳糖酶酶活力是葡萄糖的2.77倍;而以可溶性淀粉、蔗糖、果糖替代葡萄糖提供碳源时,胞外乳糖酶酶活力明显降低。故半乳糖为菌株XWSP1发酵产胞外乳糖酶最适培养基碳源组分。
由图10可知,与仅添加蛋白胨或酵母粉相比,以蛋白胨和酵母粉同时添加至培养基中,菌株XWSP1的产酶能力较好;当添加蛋白胨15 g/L、酵母粉20 g/L时,菌株XWSP1产酶能力最佳;仅有蛋白胨或酵母粉一种组分时,菌株XWSP1的产酶能力较差。
2.5 乳糖酶酶学性质分析
2.5.1pH对酶活的影响
由图11可知,在反应温度不变的条件下,胞外乳糖酶在pH为5.0~8.0范围内均维持较高的酶活力,其中pH为7.0时酶活力最高,pH大于7.0或小于7.0时酶活下降;该实验中不同pH反应条件(pH为5.0~8.0)下的酶活力均高于0.2 mg/mL β-半乳糖苷酶/GAL标准品的酶活。
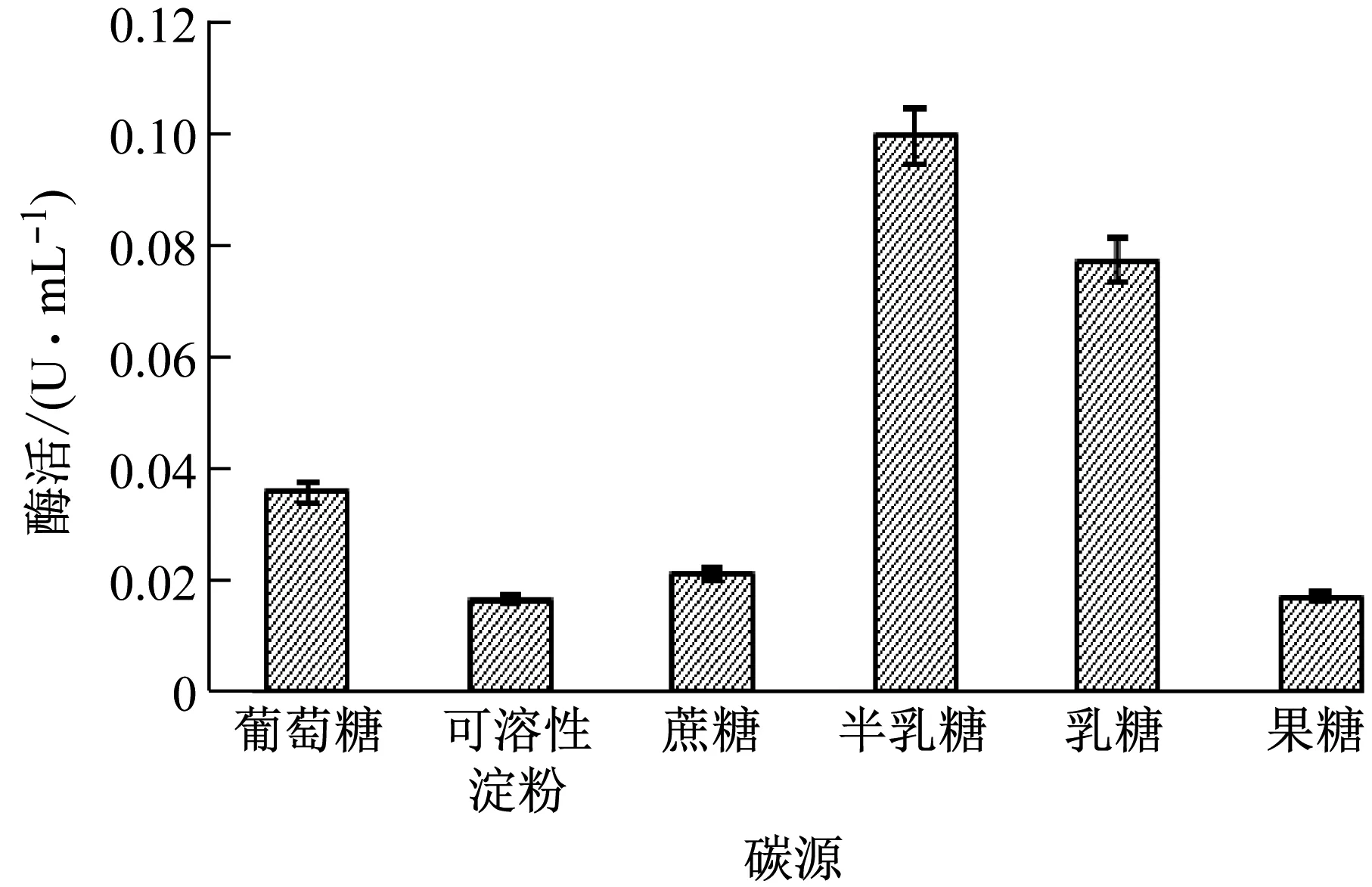
图9 培养基碳源对乳糖酶酶活力的影响
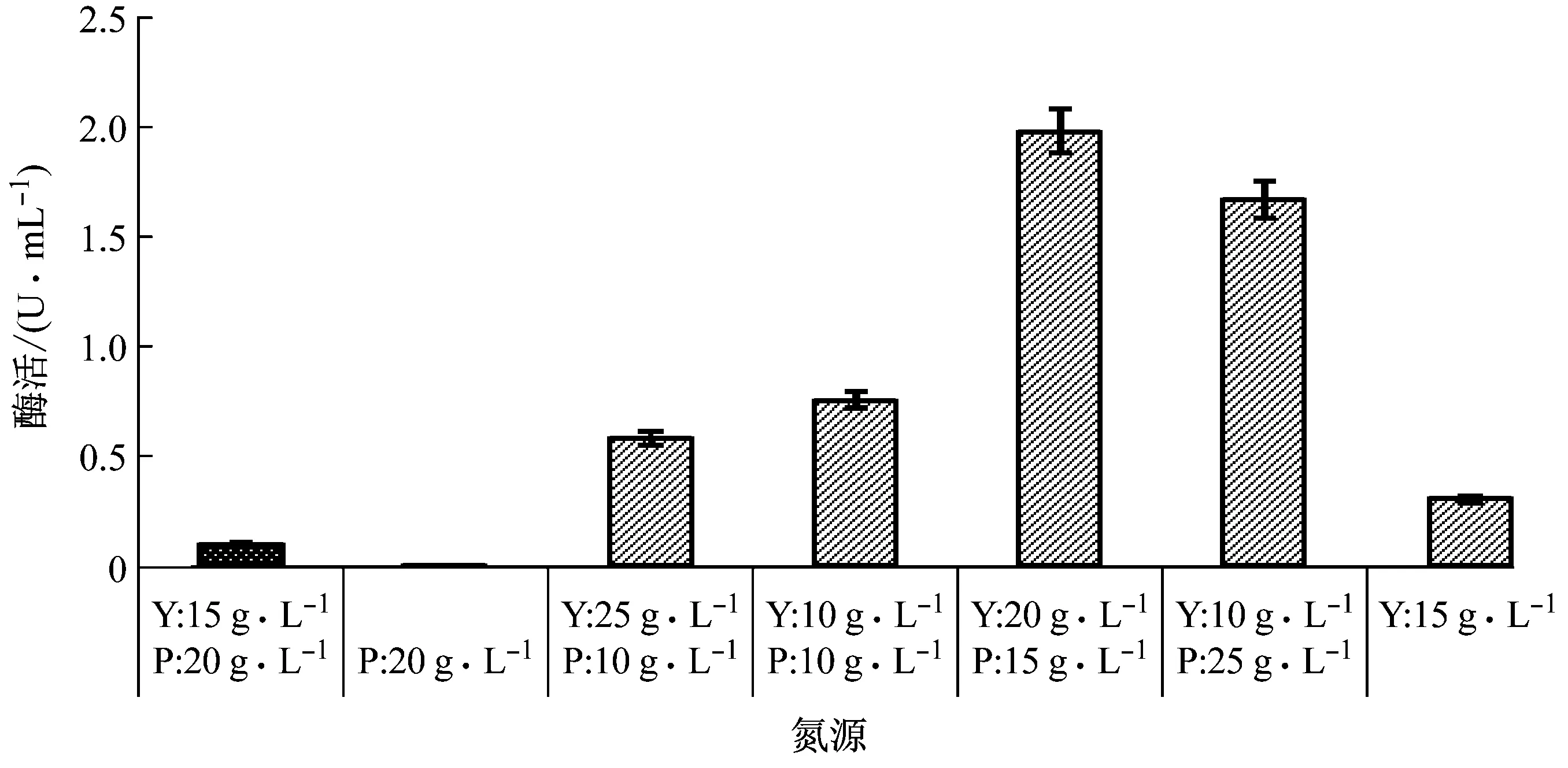
注:P为蛋白胨,Y为酵母粉
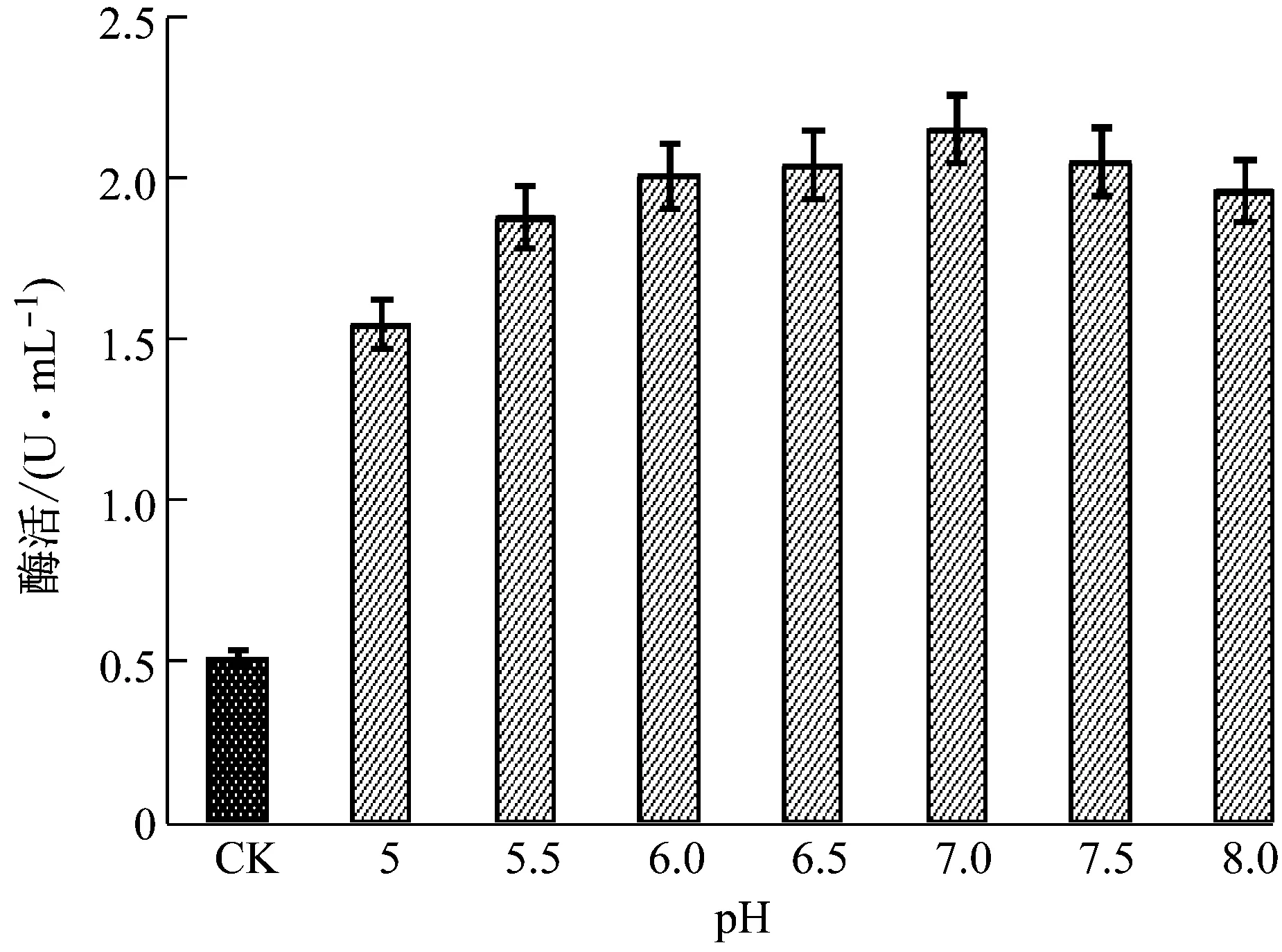
注:CK:β-半乳糖苷酶/GAL标准品,其标准酶活力为110 U/mg,本研究所用β-半乳糖苷酶溶液浓度为0.2 mg/mL。
2.5.2金属离子对酶活的影响
由图12可知,不同金属离子对胞外乳糖酶酶活力起到不同的作用,Ca2+、Mn2+、Mg2+和Cu2+对酶活有抑制作用,其中Cu2+对酶活有抑制作用最强,K+、Zn2+和Na+对胞外乳糖酶酶活力几乎没有影响。

图12 金属离子-胞外乳糖酶酶活力关系图
3 讨论与结论
酵母菌是目前工业化生产乳糖酶的主要菌种来源[19],但针对近玫色锁掷酵母所产乳糖酶的研究较少。本研究从土壤中分离出一株产乳糖酶菌株,经鉴定该菌株在分类学上与近玫色锁掷酵母属聚为一族,故命名为:近玫色锁掷酵母XWSP1(SporidioboluspararoseusXWSP1),该菌株的保藏编号为CCTCC NO: M2019119。近玫色锁掷酵母XWSP1在蛋白胨 15 g/L、酵母粉 20 g/L、半乳糖20 g/L和初始pH7.0的培养基中,以1×106CFU/mL接种浓度、4%接种比例,28 ℃ 180 r/min恒温振荡培养培养54 h时,其分泌的胞外乳糖酶具有最高的酶活;近玫色锁掷酵母XWSP1所产胞外乳糖酶最适反应pH为7.0,Ca2+、Mn2+、Mg2+和Cu2+对酶活有不同程度的抑制作用,其中Cu2+对酶活抑制作用最强。
不同培养条件和反应条件对乳糖酶酶活具有一定的调节作用。GHOSH M等人研究表明CandidapseudotropicalisB57产乳糖酶最适温度为30 ℃、最适pH为7.0[20];ITOH T等人研究发现当培养基中的pH为6.6~6.8时,最适合乳酸克鲁维酵母产乳糖酶[21];SONG C等人对psychrotolerantyeastGuehomycespullulans17-1培养条件和酶学特性进行研究,培养基中添加乳糖可以促进乳糖酶的产生,不同比例的氮源亦可以促进菌株生长和乳糖酶的产生[5]。张明丽等人的研究表明Zn2+、Cu2+、Pb2+等离子对节杆菌代谢产生的乳糖酶具有显著的抑制作用,其中Cu2+对乳糖酶的抑制作用最强[22];BIAKOWSKA A M等人的研究表明,Ca2+、Mn2+普遍被认为是酶抑制剂,其研究把Mg2+作为乳糖酶激活剂使用,同时亦指出不同浓度的Mg2+所起到的作用亦不同,过高浓度的Mg2+可以起到抑制乳糖酶的作用[11]。本研究SporidioboluspararoseusXWSP1最适培养温度为28 ℃,其所产胞外乳糖酶最适pH为7.0,并在pH为5.5~8.0范围内较稳定,适宜于工业化应用,后续将进一步研究乳糖酶代谢调控机理。
