稻瘟病菌分泌蛋白MoCDIE2 诱导植物细胞死亡的功能分析
2020-01-14吴航李智强刘文德
吴航 李智强 刘文德
(中国农业科学院植物保护研究所,北京 100193)
稻瘟病是由稻瘟病菌引起的一种真菌病害,可导致水稻的大面积减产,产量损失在10%-30%,严重发生时甚至绝收。有研究表明,全球每年因稻瘟病而损失的粮食可以养活大约6000 万人口[1]。近年来,稻瘟病菌的致病机理已成为研究热点。对稻瘟病菌侵染结构的研究表明:隔膜蛋白Sep3 的GTPases 通过改变细胞骨架结构形成侵染钉,促进5.9 μm 环状结构的形成,环状结构和F-actin 共同定位在附着胞孔周围,导致细胞质膜的弯曲和侵染钉的产生,促进稻瘟病菌的侵染[2]。稻瘟病菌侵染水稻过程中,多种信号传递和转导途径参与了稻瘟病菌和水稻相互作用,如丝裂原活化蛋白激酶(Mitogenactivated protein kinase,MAPK)途径、环腺苷酸(cyclic adenosine monophosphate,cAMP)途径和钙离子信号途径。稻瘟病菌中的MAPK 途径主要有3 种:Pmk1,Mps1 和Osm1[3-5]。另有研究表明:钙离子信号途径参与稻瘟病菌生长、侵染和致病力的形成。例如,受钙调磷酸酶调控的转录因子CRZ1 缺失突变体会导致分生孢子形成率降低和致病力的减弱[6]。cAMP 途径也参与稻瘟病菌附着胞的形成,腺苷酸环化酶Mac1基因缺失突变体不能形成附着胞[7]。
在病原物与植物相互作用的分子机制中,由病原物引起的植物防卫免疫反应可以分为两个层次[8-9]。包括模式识别受体(Pattern recognition receptor,PRRs)识别而引起的免疫防卫反应(PAMP-Triggered Immunity,PTI)[10]与 病 原 物 效应蛋白激发引起的免疫防卫反应(Effector triggered immunity,ETI)[11]。在PTI 中,植物的模式识别受体PRRs(如EFR 和 FLS2)识别病原菌的病原相关分子模式PAMPs(如elf18/elf26 和flg22)引起植物一系列生理生化反应,进而增强植物自身抗性[12]。ETI 产生比PTI 更快速和强烈的免疫反应,并引起植物的细胞死亡。例如,水稻的Pi-ta 蛋白C 端的富含亮氨酸区域(Leucine-rich domain,LRD)和稻瘟病菌的AvrPita 专化性直接结合,引起Pi-ta 介导的防卫反应[13]。稻瘟病菌的效应蛋白AvrPiz-t 抑制RING-finger 类型泛素连接酶APIP6 的泛素化活性,从而抑制PTI 反应[14]。
植物病原物分泌的效应蛋白可以促进病原物的侵染,探究病原菌效应蛋白的结构和进化,明确其在致病过程中的作用,具有重要科学意义。研究表明:细菌通过Ⅲ型分泌系统(Type Ⅲ secretion system,T3SS)分泌多种效应蛋白进入植物细胞内引起非寄主植物的过敏性反应或寄主植物的抗性反应[15];真菌的吸器不仅是吸取和交换营养的结构,也是分泌效应蛋白的首要位点[16];卵菌的吸器不仅是专化性的营养结构,也是效应蛋白的分泌位点[17]。
第一个被克隆的稻瘟病菌效应蛋白Pwl2 是在研究稻瘟病菌侵染弯叶画眉草(Eragrostis curvula)的过程中被发现的[18]。Pwl2 同源蛋白Pwl1 同样可以阻止稻瘟病菌对弯叶画眉草的侵染[19]。在水稻原生质体中瞬时表达Avr1-CO39 引起Pi-CO39专化性过敏性反应,表明Pi-CO39 特异识别Avr1-CO39[20-21]。稻瘟病菌MC69基因编码一个假定的分泌蛋白,MC69缺失的突变体的致病力下降,但对附着胞形成没有影响。在附着胞形成以后,MC69的缺失突变体不能形成侵染菌丝,因此,效应蛋白MC69在稻瘟病菌致病过程中有重要作用[22]。
对稻瘟病菌分泌效应蛋白的结构、进化和其在致病过程中的作用的研究,可为植物与病原菌协同进化和植物病害的防治提供理论依据。本研究通过农杆菌介导的瞬时表达系统,筛选到能诱导非寄主植物细胞死亡的稻瘟病菌候选效应蛋白MoCDIE2,对其进行系统生物信息学分析。然后构建MoCDIE2的敲除突变体,并对其生长表型与致病性进行分析,为进一步研究MoCDIE2 的功能奠定基础。
1 材料与方法
1.1 材料
本研究所用烟草材料为本生烟草,水稻材料为感病品种CO39,稻瘟病菌Guy11 菌株,农杆菌Agrobacterium tumefaciens菌株EHA105 均由中国农业科学院植物保护研究所植物抗病功能基因研究组保存。
1.2 方法
1.2.1 水稻培养 将水稻种子浸泡于盛有水的培养皿中,37℃恒温培养24 h 后沥干,继续培养48 h 催芽至种子破胸露白。将出芽的种子播种到盛有营养土的小方盒里,每盒6-9 粒种子。
1.2.2 烟草注射与瞬时表达 将目的基因的瞬时表达载体电击转化到农杆菌EHA105。转化后的EHA105 菌株接种于LB 液体培养基,28℃ 210 rpm条件下培养48 h。将培养后的菌液离心,用侵染液[10 mmol/L MgCl2,0.5 mmol/L MES(pH=5.7),200 μmol/L 乙酰丁香酮]重新悬浮菌块并调整菌液浓度至OD600=1.5。将目的基因的农杆菌菌液与P19菌液(OD600=1.0)等体积混合。室温条件下黑暗静置重悬的菌液3 h。使用1 mL 注射器将混合后的菌液注射到生长期为4-5 周的烟草叶片上,注射液浸润叶片面积为80%,并用记号笔标下注射范围。
1.2.3 RNA 提取和qRT-PCR 用TRIzol 法提取稻瘟病菌Guy11 的菌丝总RNA,使用RNA 反转录试剂盒(Promega 生物公司)获得cDNA,稀释cDNA 作为qRT-PCR 模板。qRT-PCR 实验中使用的PCR 引物(表1)由Primer Premier 5.0 软件设计,并由北京擎科生物技术有限公司合成。
本实验使用全式金公司的TranStart Top Green qPCR SuperMix(+Dye Ⅰ)试剂盒进行qRT-PCR反应。反应体系为20 μL,以稻瘟病菌Actin 基因(MGG_03982)为内参基因,并使用ABI Prism7500仪器完成,按照2-ΔΔct计算表达量。
1.2.4 Split PCR 方法构建MoCDIE2基因敲除突变体 根据数据库(https://www.ncbi.nlm.nih.gov/gene/)提供的基因组序列,以稻瘟病菌菌株Guy11的基因组DNA 为模板,MoCDIE2-LBCK/MoCDIE2-LB-R 和MoCDIE2-RBCK/ MoCDIE2-RB-F 为 引 物 进行第一轮PCR 扩增MoCDIE2上下游2 kb 序列LB和RB。以第一轮的PCR 产物的100 倍稀释液为模板,LB-F/HYG-R1 和RB-R/HYG-F1 为引物扩增LB+HY和YG+RB 片段。将LB+HY 和YG+RB 片段的PCR产物混合后转化到稻瘟病菌原生质体中。经潮霉素平板抗性压力筛选得到转化子,并提取转化子的DNA 和RNA,进行转化子的鉴定。
1.2.5MoCDIE2突变体的表型分析 从野生型Guy11 和突变体菌落边缘打孔取同等大小的圆形菌块接菌到CM(Complete medium)固体培养基上,28℃条件下黑暗培养7 d 后观察菌落生长形态并测量菌落直径。
1.2.6MoCDIE2突变体致病力检测 培养稻瘟病菌菌株Guy11 和突变体菌株,25℃条件下持续光照至产生足够的孢子数。用0.05%的Tween-20 水溶液洗脱孢子,调整孢子悬浮液浓度至5×104个/mL。将制备的孢子悬浮液均匀喷施在“三叶一心”期的水稻叶片上。喷施结束,用透明薄膜密封黑暗处理24 h,再光照5-7 d 后,统计病害发生情况。
1.2.7 生物信息学分析 在NCBI 数据库中进行BLASTP 检索,获得19 个与MoCDIE2 高度同源的氨基酸序列,用CLUSTAL W 比对分析,进而用MEGA 7.0 软件构建系统进化树。
2 结果
2.1 MoCDIE2基因的生物信息学分析
在稻瘟病菌中,预测大约有1306 个基因编码分泌蛋白[23-24]。实验室前期通过基因组表达谱,MPSS[25-26],RL-SAGE[27]和SBS[28]3 种 技 术鉴定出851 个稻瘟病菌侵染水稻过程中表达的分泌蛋白[29]。选取其中一个分泌蛋白(MGG_03356,MoCDIE2)作为候选效应蛋白,搜索比对稻瘟病菌全基因组数据库,发现MoCDIE2的cDNA 全长为1068 bp,编码一个含有355 个氨基酸的蓖麻毒素B凝集素蛋白(Ricin B lectin protein)。利用MoCDIE2的氨基酸序列在NCBI 数据库中进行比对搜索,发现其在19 个真菌物种中均有同源性较高的蛋白,再利用Clustal W 比对分析MoCDIE2 及其同源性最高的其它19 个蛋白氨基酸序列,最后用MEGA 7.0 构建系统发育进化树分析MoCDIE2 与其它19 个蛋白氨基酸序列的亲缘关系的远近。同源蛋白对比分析表明:MoCDIE2 与子囊菌门的根腐病菌(Monosporascus ibericus)、炭疽病菌(Colletotrichum gloeosporioides)、痂囊腔菌(Elsinoe australis)的同源性高达82%、80%和75%。在MoCDIE2 的19 个同源性最高的真菌蛋白中,大多来源于子囊真菌,个别属于担子菌,如:灰盖鬼伞菌(Coprinopsis cinerea)和羽瑚菌(Pterula gracilis)(图1)。这说明MoCDIE2 同源蛋白保守存在于真菌物种中。利用DNAMAN 进行MoCDIE2 和另外4 个丝状真菌中的同源蛋白氨基酸序列比对分析发现:MoCDIE2 在丝状真菌中具有高度的保守性,并且蛋白氨基酸数目相差不大(图2)。
2.2 MoCDIE2在烟草中的瞬时表达
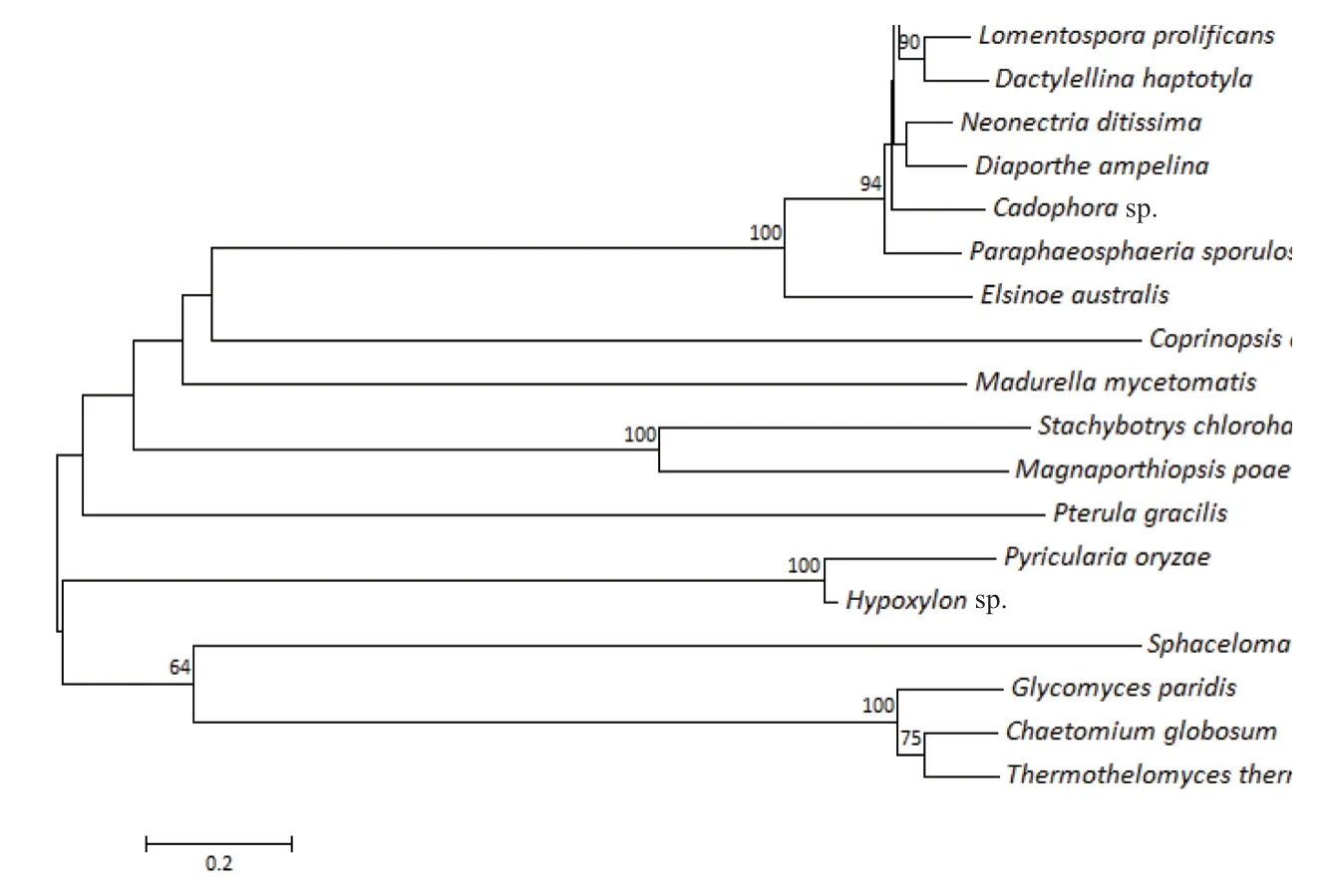
图1 MoCDIE2 和其它物种中同源蛋白的进化树分析

图2 MoCDIE2 和其它物种中同源氨基酸序列比对分析
为了筛选能诱导植物细胞死亡的效应蛋白,将MoCDIE2构建到植物瞬时表达载体pGDR 质粒,利用农杆菌介导的转化系统将MoCDIE2表达载体注射到烟草叶片上进行瞬时表达,同时设置BAX 和空载pGDR 注射的烟草叶片分别作为阳性对照和阴性对照,注射后观察烟草叶片表型。结果表明:72 h 后,在对照组中,阳性对照BAX 表达载体注射的烟草叶片有严重细胞死亡现象,阴性对照pGDR 和P19 表达载体注射的烟草叶片未出现细胞死亡。在实验组,MoCDIE2 的瞬时表达载体注射72 h 后,烟草叶片发生明显皱缩的细胞死亡现象(图3)。结果表明:MoCDIE2 的瞬时表达能引起非寄主植物烟草叶片的细胞死亡,而细胞死亡是植物限制植物病原物生长的一种防御机制,推测MoCDIE2 可能在植物-植物病原物相互作用过程中起重要作用。
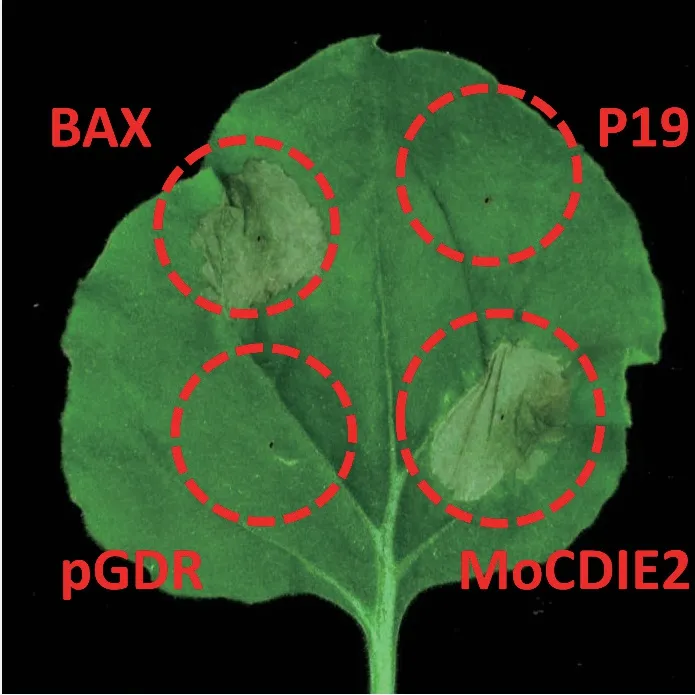
图3 MoCDIE2 在烟草叶片细胞中的瞬时表达
2.3 MoCDIE2基因敲除突变体的获得
为构建MoCDIE2敲除突变体,进一步分析其功能,我们利用Split-PCR 方法(图4-A)扩增MoCDIE2上下游片段和HYG 片段,再利用PEG 介导的原生质体转化法将LB+HY 和RB+YG 片段混合转化到稻瘟病菌Guy11 原生质体中,RT-PCR 验证MoCDIE2敲除转化子。PCR 结果表明:由LB-F/H YG-R1 扩增得到2488 bp 的LB+HY 的条带,由RBR/HYG-F1 扩增得到2250 bp 的RB+YG 的条带(图4-B);进一步RT-PCR 验证,使用引物MoCDIE2-F/MoCDIE2-R 在转化子2、3、5 和Guy11 中能扩增出条带,而在阳性转化子1、4 和6 中不能扩增出条带(图4-C),说明成功获得3 个具有潮霉素抗性的MoCDIE2基因敲除突变体,在后续实验中分别命名为MoCDIE2-KO1、MoCDIE2-KO2 和MoCDIE2-KO3。
2.4 MoCDIE2表型鉴定
2.4.1 营养生长 在CM 固体培养基上培养稻瘟病菌的野生型菌株Guy11 和3 个MoCDIE2基因的敲除菌株MoCDIE2-KO1、MoCDIE2-KO2 和MoCDIE2-KO3,一周后观察菌落形态发现:敲除突变体和野生型菌株在CM 平板上的生长速率未见差异,突变体和野生型的菌落形态无明显变化(图5-A)。通过测量野生型Guy11 和MoCDIE2基因的敲除突变体的菌落并进行统计分析的结果表明:突变体和野生型的菌落直径没有显著差异(图5-B)。因此,MoCDIE2基因可能不直接参与调控稻瘟病菌的营养生长。
2.4.2 致病性测定 将野生型菌株Guy11 和MoCDIE2基因的敲除菌株分别接种到燕麦固体培养基上,光照培养诱导产孢后,将野生型菌株Guy11 和MoCDIE2基因的敲除菌株的孢子悬浮液调整浓度至5×104个/mL,喷雾接种到水稻叶片上,7 d 后观察水稻叶片的发病情况、病斑大小和病斑数量等。结果(图5-C)表明:MoCDIE2的敲除菌株和野生型菌株均能导致接种后的水稻叶片产生典型稻瘟病病斑,且病斑大小和数量没有明显的差异。因此,MoCDIE2基因的缺失对稻瘟病菌的致病性没有影响。
3 讨论
在植物和病原物的相互作用中,效应蛋白在促进病原物成功侵染和抑制植物防卫反应方面都起重要调控作用[30]。因此采取合适策略分离和克隆效应蛋白对植物病原物的效应蛋白功能研究以及明确病原物与植物之间的相互作用机制有重要意义。
在已被鉴定到的稻瘟病菌效应蛋白中,Pwl1[19]、Pwl2[18]和AvrPiz-t[31]等都是通过图位克隆策略分离克隆得到。图位克隆是根据与目的效应蛋白基因紧密连锁的分子标记在染色体上的位置,通过构建遗传连锁群体以及筛选基因组文库等步骤逐步确定和分离目的基因的方法。图位克隆策略的优点是未知基因的序列信息和基因表达产物的序列、结构等相关信息的条件下,可以获得该基因。因此,图位克隆作为一个重要分离克隆未知基因的技术手段,为探索未知效应蛋白的功能提供技术支持。在这一策略中,最重要的因素是开发与目的效应蛋白基因紧密连锁的分子标记以及细菌人工染色体(Bacterium artificial chromosome,BAC)或酵母人工染色体(Yeast artificial chromosome,YAC)等基因组文库的构建。但是传统的图位克隆策略实验周期长,不够高效,操作步骤繁琐。在完成了稻瘟病菌全基因组测序的基础上,借助植物异源表达系统瞬时表达候选效应蛋白为更加快速地克隆目标效应蛋白提供了技术支持。
本研究应用高效的由农杆菌介导的异源表达策略筛选稻瘟病菌效应蛋白。将候选效应蛋白构建到pGDR 表达载体,再利用农杆菌介导的瞬时表达策略快速、有效地初步鉴定效应蛋白的功能。农杆菌介导的转化系统具有操作简单,周期短的优点。利用生物信息学分析技术结合农杆菌转化系统,我们成功筛选到一个能诱导植物细胞死亡的效应蛋白,相比较于图位克隆策略中的周期长、工作量大等缺点,异源表达策略能快速的筛选到诱导非寄主植物细胞死亡的效应蛋白。高效便捷地筛选到诱导植物细胞死亡的效应蛋白,可以为后续效应蛋白的功能研究奠定基础,同时提高研究的效率。但如何保证效应蛋白表达载体的顺利转化与稳定表达是异源表达策略需要面对的问题。首先,通过制备活性较高的农杆菌感受态细胞是提高效应蛋白表达载体转化效率的关键之一。其次,烟草植株的选择也是提高效应蛋白表达载体表达效率的重要因素。在进行瞬时表达的过程中,需要注意植物的生长状态,选取健康烟草植株,因为较大或老的烟草叶片不利于效应蛋白表达载体的注射和渗透,从而会导致表达不稳定,影响细胞死亡表型的观察。
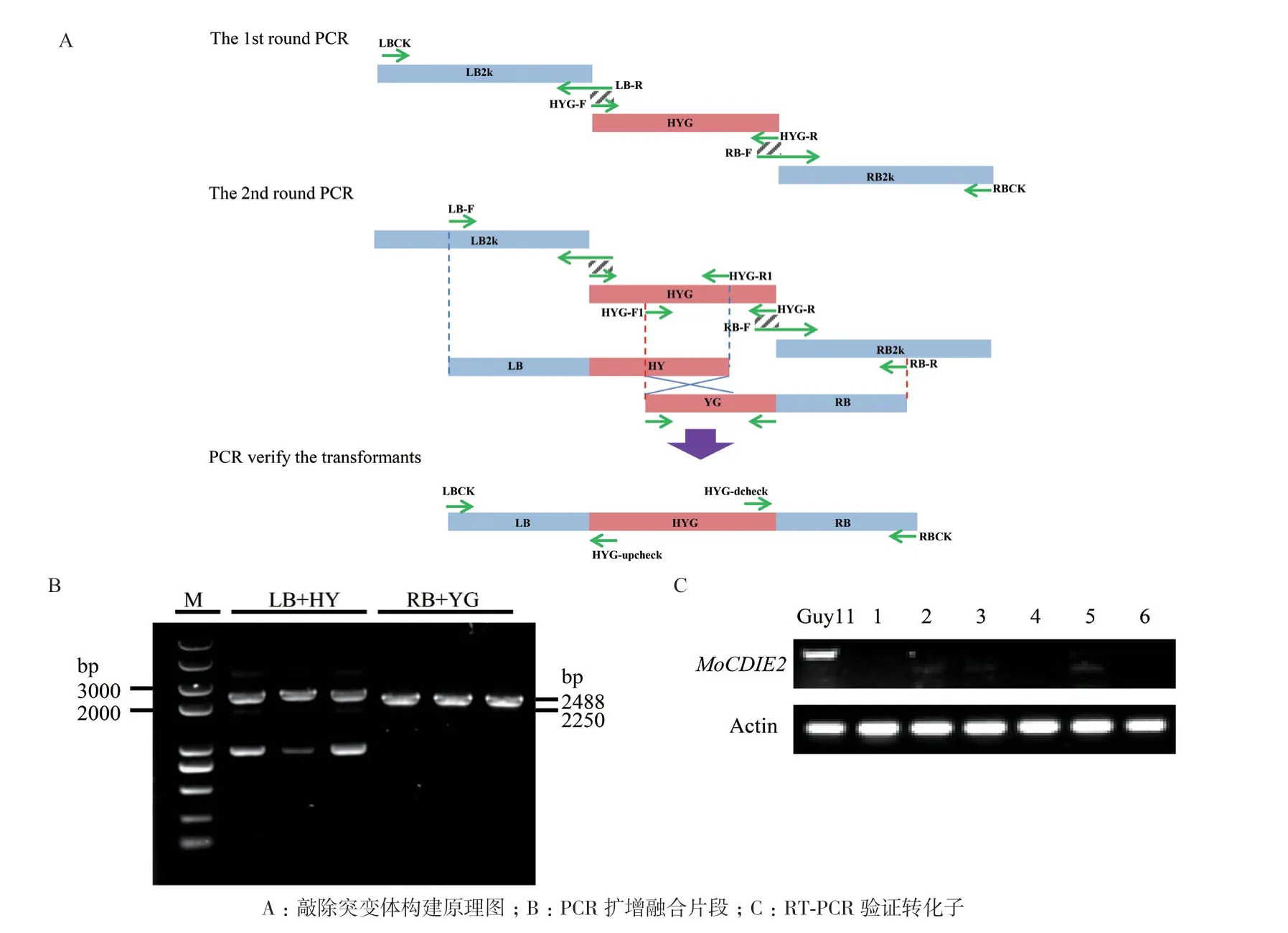
图4 MoCDIE2 敲除突变体的构建与验证
在本实验中,能诱导植物细胞死亡的MoCDIE2基因编码一个蓖麻毒素B 凝集素蛋白。蓖麻毒素蛋白是由A、B 两条肽链通过二硫键连接的一种剧毒植物糖蛋白。蓖麻毒素蛋白的A 链在B 链的协助下,穿透细胞膜并破坏细胞核蛋白体60S 亚蛋白,从而抑制蛋白质的合成,导致细胞死亡。在蓖麻毒素的B 链上有3 个重要的半乳糖结合位点,因此通过特异性抑制B 链的结合位点,可能阻断细胞死亡的发生。有研究表明:蓖麻毒素能够诱导小鼠淋巴细胞发生凋亡而引起细胞死亡,并且检测到被蓖麻毒素处理后的细胞的线粒体膜电位发生了显著变化[32]。因此,MoCDIE2 引起的细胞死亡可能和植物细胞线粒体膜电位的变化有关,在后续的研究中,可以检测细胞的线粒体膜电位,进一步明确MoCDIE2 引起的细胞死亡的作用机制。
在本实验中,与野生型相比,MoCDIE2敲除突变体对稻瘟病菌的营养菌丝生长和对水稻的致病性方面没有显著差异,可能是效应蛋白的功能冗余性造成的。其次,MoCDIE2 可能不直接参与调控稻瘟病菌的生长,也不直接参与水稻的致病过程,但MoCDIE2 或许在侵染过程中与其他未知的效应蛋白发生作用,进而导致某些生理生化过程的发生。稻瘟病菌中存在着1000 多个分泌蛋白,生物信息学数据表明,大多数分泌蛋白都没有功能注释,敲除或过表达某一单个分泌蛋白基因,并不足以影响稻瘟病菌的致病性。这是因为在稻瘟病菌中可能存在其他功能相似的一个或一类基因继续行使其功能,从而继续保持稻瘟病菌的生长和致病特性,多数功能冗余的效应蛋白存在于稻瘟病菌中,为效应蛋白的功能研究带来了困难和挑战。
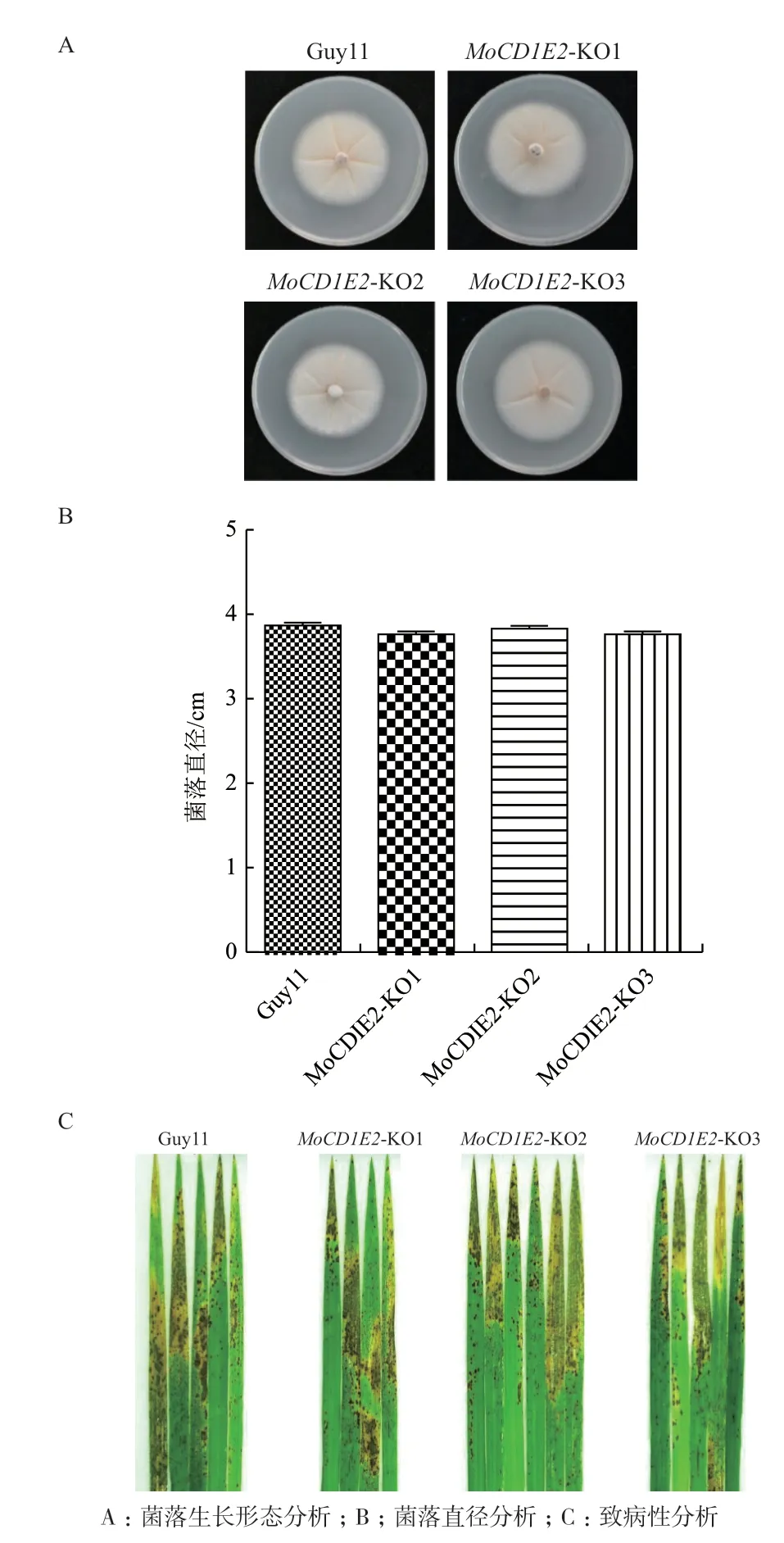
图5 稻瘟病菌MoCDIE2 的功能分析
4 结论
本研究通过农杆菌介导的转化系统筛选到稻瘟病菌的分泌蛋白MoCDIE2(MGG_03356)能诱导非寄主植物烟草细胞死亡。通过生物信息学分析发现:MoCDIE2 编码一个蓖麻毒素B 凝集素蛋白,在子囊菌门的根腐病菌(M. ibericus)、炭疽病菌(C.gloeosporioides)、痂囊腔菌(E. australis)均存在高度同源蛋白,其同源蛋白也存在于其他16 种丝状真菌中,并具有高度的保守性。通过Split-PCR 和PEG介导的原生质体转化系统构建MoCDIE2敲除突变体,获得了3 个MoCDIE2的敲除突变体。进一步对MoCDIE2功能分析表明:MoCDIE2基因的缺失不影响稻瘟病菌的生长和对水稻的致病性。
