双免耕覆盖对旱地土壤微生物、酶活性和冬小麦干物质转运的影响
2020-01-05邵运辉李俊红岳俊芹丁志强张德奇方保停李向东
邵运辉,李俊红,岳俊芹,丁志强,张德奇,方保停,杨 程,李向东
(1.河南省农业科学院小麦研究所,河南郑州 450002; 2.洛阳农林科学院,河南洛阳 471023; 3.中国农业科学院洛阳旱农试验基地,河南洛阳 471023)
在豫西旱区,干旱少雨且水土流失严重,自然降水是作物生产的主要水资源[1]。保护性耕作主要指秸秆还田、免耕播种和残茬覆盖等耕作手段,可有效增加土壤有机质含量,调节土壤的C/N、温度和水分变化,对作物有一定的增产效果[2]。免耕等保护性耕作基本上不扰动土层,秸秆和根茬等有机物腐解后的残茬和根系分泌物主要积累在表层土壤中,为土壤微生物的生命活动提供了足够的能源[3]。土壤微生物通过分解动植物残体参与生态系统的能量流动和物质循环,其数量是评价土壤肥力的重要指标之一。土壤酶则主要来源于活体植物、死亡植株残体,直接参与植物与土壤之间的物质循环和土壤中生物的生化反应,可提高土壤养分的可用性,其活性大小直接影响土壤肥力[4]。保护性耕作可增加和维护土壤微生物种类和数量,提高土壤酶的活性,改善作物根区环境,提高作物抗逆性,促进作物高产、稳产,并提高土壤质量和改善生态环境[5-6]。土壤微生物和土壤酶共同影响并推动着土壤的代谢过程,二者的活性关系密切[7]。土壤酶活性、土壤微生物的代谢和种类与植物根系生长状况有一定关系。林雁冰等[6]认为,冬小麦根系生理代谢对根区土壤微生物数量和种类有显著影响,覆盖可以提高土壤细菌、真菌和放线菌的数量。土壤酶活性高,则小麦根系衰老较慢,冬小麦根系的生长状况直接影响地上部生长状况和抗旱能力,最终影响其产量[8-11]。冬小麦产量的物质来源主要有两个方面,一是花后光合同化物的直接输入,另一个是花前贮藏在营养器官的光合同化物的再运转。王征宏等[9]研究发现,干旱对冬小麦干物质转运有较大的影响,灌浆前期干旱会导致花前贮藏物质向籽粒的转运量降低。
目前,国内外对保护性耕作下土壤微生物、酶活性及干物质积累等方面研究有较多的报道,但有关旱作区双免耕覆盖对冬小麦土壤微生物和酶活等的影响尚未见报道。鉴于此,本研究拟对双免耕覆盖下旱地冬小麦田土壤微生物、酶活及冬小麦花前、花后干物质积累和转运进行深入研究,旨在为旱作区冬小麦增产提供理论依据,为选择合理有效的保护性耕作模式提供参考。
1 材料和方法
1.1 试验地概况
试验设在洛阳农科院旱农基地防渗精确水分池,土壤为潮褐土,质地中壤,耕层容重1.53 g·cm-3,田间相对持水量23.48%,饱和含水量 33.36%。该区域年均辐射量491.5 kJ·cm-2,年均气温14 ℃,日平均温度超过10 ℃的天数约210 d,大于10 ℃的积温可达4 000 ℃,年蒸发量 1 841.7 mm。土壤耕层有机质为15.8 g·kg-1,碱解氮(N)为62.7 mg·kg-1,速效磷(P2O5)为10.39 mg·kg-1,速效钾(K2O)为166.0 mg·kg-1。种植模式为冬小麦、玉米一年两 熟制。
1.2 试验设计
本研究于2013年10月—2014年6月进行。设2个处理:(1)双免耕覆盖(NT),冬小麦收获时留茬35~40 cm,秸秆脱粒后还田并覆盖于地表,不翻耕土壤,铁茬播种玉米;玉米收获后留根茬,其秸秆覆盖于冬小麦行间,冬小麦采用人工开沟种植,尽量少扰动土壤,等行播种;(2)传统耕作(CK),即深翻耕25~30 cm(冬小麦、玉米秸秆均不还田)。小区面积为4 m×4 m。供试冬小麦品种为洛旱7号,种植密度13.5×104株·hm-2。玉米品种为洛玉7号,种植密度为4.5×104株·hm-2。
1.3 测定项目与方法
1.3.1 土样采集
在冬小麦苗期(seedling stage,SS)、越冬期(over winter stage,OS)、拔节期(jointing stage,JS)、灌浆期(filling stage,FS)、成熟期(maturation stage,MS)的晴天上午10:00左右,每个小区按五点法取土样,每点取土深度为0~60 cm,每10 cm取一次,剔除石块、植物残根、蚯蚓等杂物后装袋,样品风干后经研磨,过1 mm筛,采用四分法选取待测样品。
1.3.2 土壤相关酶活性测定[7]
土壤脲酶活性采用苯酚钠比色法测定;土壤蛋白酶活性采用茚三酮比色法测定。
1.3.3 土壤微生物数量测定[4]
细菌采用牛肉膏蛋白胨培养基平板计数法测定;真菌采用马丁氏培养基平板计数法测定;放线菌采用淀粉铵盐培养基平板计数法测定。
1.3.4 冬小麦干物质积累相关指标测定
在冬小麦开花期和成熟期,每个小区随机取10株具代表性植株地上部,开花期取样后将植株分为叶片、茎杆+叶鞘和穗三部分,成熟期取样后将植株分为叶片、茎秆+叶鞘、颖壳+穗轴、籽粒四部分。将样品于105 ℃杀青15 min,65 ℃烘干至恒重,测定干物质积累量。相关指标计算公式如下[12]:
花前同化物转运量=开花期植株干重-成熟期营养器官干重
(1)
花前同化物转运效率=花前同化物转运量/开花期植株干重×100%
(2)
花前同化物对籽粒产量贡献率=花前同化物转运量/成熟期籽粒干重×100%
(3)
花后同化物积累量=成熟期籽粒干重-花前同化物转运量
(4)
花后同化物对籽粒产量贡献率=花后同化物积累量/成熟期籽粒干重×100%
(5)
1.3.5 产量及其构成因素测定
在冬小麦收获期,按一般方法调查穗数、穗粒数,测定千粒重,整区收获计产。
1.4 数据处理
采用Excel 2010 和DPS 7.05进行数据处理与统计分析,用LSD法进行差异显著性检验。
2 结果与分析
2.1 双免耕覆盖对麦田土壤中微生物数量的 影响
2.1.1 对麦田土壤中细菌数量的影响
从图1可知,随着冬小麦生育进程的推进,不同土层的细菌数量呈现先上升后下降的趋势;以越冬期最少,拔节期最多,拔节期以后逐渐减少。双免耕覆盖下0~10 cm土层细菌数量在冬小麦不同生育时期均显著高于传统耕作(图1A),尤其是成熟前,二者差异达极显著水平,双免耕覆盖下细菌数量平均较传统耕作高94.7%。双免耕覆盖下10~20 cm土层冬小麦不同生育时期细菌数量在苗期和越冬期显著高于传统耕作外,之后二者差异达极显著水平(图1B),双免耕下细菌数量平均较传统耕作高53.3%。这种现象可能与秸秆覆盖能保存较充足的土壤水分和维持适宜的土壤温度有较大关系。
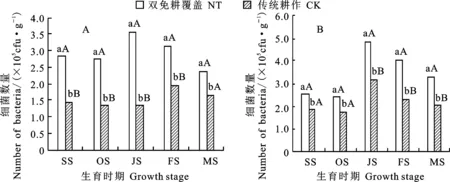
相同时期图柱上不同小写字母和大写字母分别表示处理间差异达5%和1%显著水平。下同。
2.1.2 对麦田土壤中真菌数量的影响
土壤中的真菌参与土壤中有机质的分解,在土壤腐殖质的合成、氨化作用以及团聚体的形成等过程中发挥着重要的作用,可直接影响土壤肥力[6]。 由图2可知,0~20 cm土层土壤真菌数量在越冬期最低,拔节期和灌浆期较高;双免耕覆盖下麦田土壤真菌数量均不同程度高于传统耕作。0~10 cm土层,双免耕处理土壤中真菌数量均显著或极显著高于传统耕作(图2A),拔节期和灌浆期分别较传统耕作高23.4%和34.1%。除拔节期外双免耕覆盖处理下10~20 cm土层土壤 中真菌数量均显著或极显著高于传统耕作 (图2B)。

图2 两种耕作模式对麦田不同生育时期土壤真菌数量的影响
2.1.3 对麦田土壤中放线菌数量的影响
放线菌能分解土壤腐殖质中最稳定的有机化合物,提高土壤肥力。从图3可以看出,0~20 cm土层土壤中的放线菌数量以苗期最高,0~10 cm(图3A)和10~20 cm土层(图3B),双免耕覆盖较传统耕作分别高28.5%和74.7%,差异均达极显著水平;越冬期土壤中放线菌的数量最少。从越冬期至成熟期,双免耕覆盖0~20 cm土层中放线菌数量有不同程度的增加;不同时期双免耕覆盖土壤中放线菌数量均显著高于传统耕作。
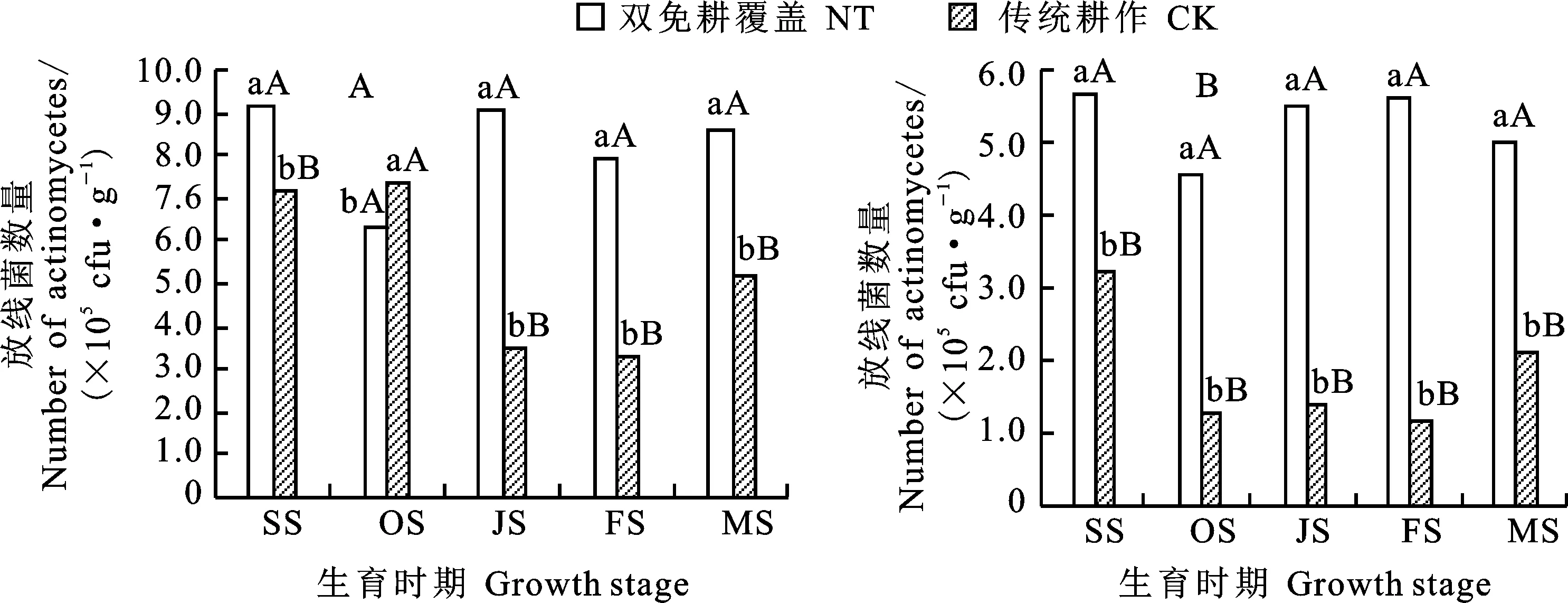
图3 两种耕作模式对麦田不同生育时期土壤中放线菌数量的影响
2.1.4 双免耕覆盖下麦田土壤中微生物总量的垂直分布
由图4可知,双免耕覆盖和传统耕作下,冬小麦土壤中微生物总量(细菌、真菌和放线菌数量之和)随土层深度的增加均呈现逐渐下降趋势。双免耕覆盖下冬小麦土壤微生物总量在不同土层均极显著高于传统耕作,0~60 cm土层较传统耕作平均高 68.2%。双免耕覆盖下冬小麦土壤0~40 cm土层中微生物总量迅速递减,而传统耕作则在0~20 cm土层呈现迅速递减趋势。
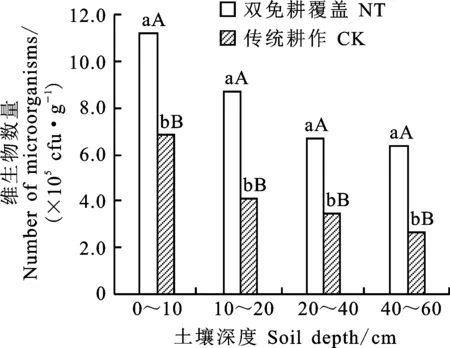
图4 两种耕作模式对麦田土壤微生物总量垂直分布的影响
2.2 双免耕覆盖对麦田土壤酶活性的影响
2.2.1 对冬小麦不同生育期土壤酶活性的影响
从图5可知,冬小麦不同生育时期0~20 cm土层土壤中蛋白酶活性的变化幅度较为缓和(图5A),脲酶活性在越冬期最低(图5B),其余生育时期的变化与蛋白酶的活性变化表现一致,均随生育进程的推进逐渐减小。双免耕覆盖下,冬小麦不同生育期土壤中的两种酶活性均高于传统耕作。在冬小麦全生育期内,双免耕覆盖下0~20 cm土层土壤中蛋白酶活性较传统耕作平均增加了33.6%,差异达极显著水平(P<0.01)。土壤中蛋白酶活性以苗期最高,较传统耕作增加了15.0%,拔节期次之,成熟期最低,成熟期双免耕覆盖仍较传统耕作高27.6%(图5A)。
双免耕覆盖下土壤中脲酶活性在冬小麦全生育期较传统耕作平均高28.6%,除成熟期外,其他生育期二者差异均达极显著水平(图5B)。土壤脲酶活性在冬小麦苗期最大,双免耕覆盖较传统耕作高28.8%。越冬期脲酶活性急剧下降,在冬小麦整个生育期中为最低,此期双免耕覆盖较传统耕作高55.3%。这可能因为双免耕覆盖有利于提高地温,从而有利于提高土壤酶活性。拔节期土壤脲酶活性逐渐上升,双免耕覆盖较传统耕作高28.6%,灌浆期又迅速下降,成熟期二者差异不显著。

图5 两种耕作模式对冬小麦不同生育时期土壤酶活性的影响(0~20 cm)
2.2.2 双免耕覆盖下麦田土壤中酶活性的垂直分布
从表1可以看出,在0~60 cm土层,双免耕覆盖下麦田土壤中蛋白酶和脲酶的活性均随着土层深度的增加逐渐降低,蛋白酶和脲酶活性分别较传统耕作平均增加了27.3%和32.7%。双免耕覆盖下蛋白酶活性在0~10 cm和40~60 cm分别较传统耕作高31.5%和37.7%,脲酶活性在10~20 cm和40~60 cm土层均较传统耕作高35.9%和36.4%,差异均达极显著水平。这表明在干旱条件下,双免耕覆盖有利于提高耕层土壤的酶活性。
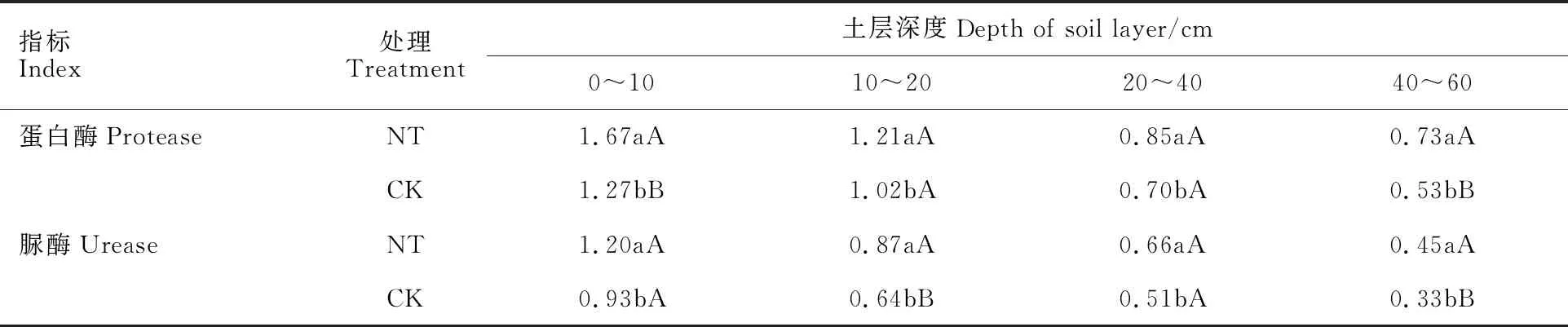
表1 两种耕作模式下麦田土壤酶活性分布
2.3 双免耕覆盖对冬小麦开花前后干物质积累和转运的影响
灌浆期是冬小麦籽粒干物质积累的主要时期,大约有70%~90%的籽粒干物质来自于灌浆期的光合同化物[13]。从表2可以看出,双免耕覆盖下,冬小麦较传统耕作增产18.4%,差异极显著;花后干物质积累量及其对籽粒产量的贡献率均极显著高于传统耕作;花后干物质积累量对籽粒产量的贡献率为44.2%,较传统耕作高 61.9%。传统耕作冬小麦花前干物质积累量对籽粒产量的贡献率为72.7%,显著高于双免耕覆盖,但干物质转运效率显著低于双免耕覆盖。整体而言,冬小麦籽粒干物质主要源自开花前干物质的转运,双免耕覆盖较传统耕作提高了旱地冬小麦开花后干物质积累量及其对籽粒产量的贡献率,为双免耕覆盖下冬小麦获得高产奠定了基础。

表2 旱作区两种耕作模式对冬小麦开花前后干物质积累和转运的影响
3 讨论与结论
3.1 双免耕覆盖对冬小麦土壤微生物的影响
本研究发现,双免耕覆盖下冬小麦土壤中细菌、真菌和放线菌的数量均显著高于传统耕作,这与赵亚丽等[2]、张星杰等[3]和王 静等[5]的研究结果基本一致,但土壤中微生物数量的多少和种类是否受秸秆还田量及还田年限的影响还需进一步研究。免耕覆盖可提高微生物增长的空间,可明显提高微生物种群数量,尤其是耕层微生物更为活跃[13-16]。随土壤深度的增加,微生物数量逐渐减少,这可能与土壤含水量、温度等条件的变化有关。双免耕覆盖下,微生物数量的变化在冬小麦各生育期不同深度土层的表现不完全一致,细菌数量以拔节期最多,真菌数量以拔节期和灌浆期较多,放线菌数量以苗期最多,这可能与冬小麦根系生长状况、土壤含水量、温度及微生物生长所需环境等有关。
3.2 双免耕覆盖对冬小麦土壤酶活性的影响
双免耕覆盖下冬小麦土壤中蛋白酶和脲酶的活性均随着土层深度的增加逐渐降低,双免耕覆盖下冬小麦土壤中酶活性均极显著高于传统耕作,这是因为在豫西旱作区,干旱是影响作物生长的主要因素,与传统耕作相比,双免耕覆盖有利于提高土壤水分含量,有利于作物根系的生长,提高土壤酶活性。这与王 静等[5]、林雁冰等[6]和邱现奎等[14]的研究结论一致。双免耕覆盖有利于冬小麦耕层土壤酶活性的增加,这与秸秆覆盖后保持土壤水分有较大关系[17],但秸秆类型、还田量及腐解速率对土壤酶活性的影响还需继续研究。
3.3 双免耕覆盖对冬小麦花前花后干物质积累及转运的影响
双免耕覆盖下冬小麦花后干物质积累量和对籽粒的贡献率均高于传统耕作,双免耕覆盖下冬小麦产量极显著高于传统耕作,可能因为双免耕覆盖可延缓植株衰老[18],提高冬小麦花后干物质积累,提高花前干物质积累和花后同化物向籽粒转运的能力,有利于冬小麦产量的提高,这与雷钧杰[8]和王健波[10]研究结论较为一致。但不同降水年型下双免耕覆盖对冬小麦花前、花后干物质积累及转运的影响还有待做深入研究。
