安达曼海浮游有孔虫群落对全新世海洋环境变化的响应
2019-12-03张玲芝向荣唐灵刚杨艺萍钟福昌
张玲芝, 向荣, 唐灵刚, 杨艺萍, 钟福昌
海洋生物学
安达曼海浮游有孔虫群落对全新世海洋环境变化的响应
张玲芝1,2, 向荣1, 唐灵刚1,2, 杨艺萍1, 钟福昌1,2
1. 中国科学院边缘海与大洋地质重点实验室(南海海洋研究所), 广东 广州 510301;2. 中国科学院大学, 北京 100039
通过对安达曼海重力柱ADM-C1的浮游有孔虫群落分析, 探讨了该区全新世以来的海洋环境演化。研究发现, 该区浮游有孔虫群落总体以热带暖水种和等为主。其中的相对丰度从早全新世至今呈逐渐降低的趋势,则呈大致相反的变化趋势。的相对丰度从11—7.9ka BP较高, 到7.9—3.8ka BP偏低, 3.8ka BP以来又逐渐升高。而的变化趋势则与大致相反。研究认为, 安达曼海不存在与冲绳海槽类似的晚全新世普林虫低值事件。浮游有孔虫群落Q型因子显示全新世海洋环境呈现三个明显的阶段变化: 早全新世 11—7.9ka BP期间, 浮游有孔虫群落以与为主, 反映了该阶段较强的夏季风降水会导致表层水体盐度较低, 同时冬季风的影响相对较强; 7.9—3.8ka BP期间, 以和为主, 指示水体盐度仍然较低, 同时次表层属种显著增多, 对应了该阶段强盛的印度夏季风, 上层海水混合强烈; 3.8—0ka BP期间, 以和为主要特征属种, 而相对丰度明显下降, 表明该时期表层海水的盐度有显著上升, 对应了印度夏季风降水的明显减弱。安达曼海浮游有孔虫群落所呈现的全新世海洋环境阶段性变化和陆地夏季风记录有很好的一致性, 也与该孔敏感粒级组分所反映的印度季风的强度变化一致, 表明热带边缘海区的有孔虫群落组合可以很好地响应区域海洋环境变化。
全新世; 安达曼海; 浮游有孔虫; 印度季风; 海洋环境
印度季风又称西南季风, 是由于印度洋北接世界最大的高原, 南临广阔的南大洋海域, 造成巨大的海陆热力差异所形成的季风系统。印度季风是影响地球上人口最稠密地区的亚洲季风系统的一个重要组成部分(Staubwasser et al, 2003)。印度夏季风受印度低压和马斯克林高压的气压梯度驱动, 对印度洋10°S以北的大部分海域以及周边陆地地区有着重要影响(An et al, 2011)。印度季风带来的丰沛降雨也为南亚地区提供了重要的水资源, 对该地区的社会发展具有重大的作用(Achyuthan et al, 2014), 对南亚文明也有着重大影响(Staubwasser et al, 2003)。安达曼海是位于东北印度洋东侧的边缘海, 地处强烈的亚洲季风活动带, 海洋表层环流的变化与季风演变具有明显的同步性 (Brown, 2007; 曹鹏, 2015)。此外, 印度夏季风带来的降水以及伊洛瓦底江的径流输入对安达曼海的水体盐度有着重要的影响, 是区域海洋环境变化的重要控制因素。由于安达曼海紧邻亚洲大陆, 在西边又存在岛弧链,使其与开阔大洋之间有一定的屏蔽, 因此能较好地保存古海洋环境演化信息, 是研究印度季风演化的重要区域。
关于印度夏季风的演化, 陆地方面的研究非常多。大量来自石笋的记录显示, 印度夏季风全新世有明显的阶段变化: 全新世早期最强盛, 随后从中全新世开始, 印度季风大体呈逐渐减弱的趋势, 晚全新世的季风强度相对呈较弱状态(Neff et al, 2001; Fleitmann et al, 2003; Yuan et al, 2004; Dykoski et al, 2005; Wang et al, 2005)。来自湖泊孢粉的记录同样发现全新世的印度季风降水和温度呈现明显的阶段性变化, 全新世早期夏季风强度最强, 随后逐渐减弱(Chen et al, 2014)。此外, 孢粉记录还揭示了在中全新世存在一个温暖而湿润的全新世气候适宜期(Holocene climate optimum), 大约从8.4ka BP开始, 结束时间在不同记录中有差异, 最晚可到4.2ka BP (Shen et al, 2006; Xiao et al, 2005; Yang et al, 2016)。相较陆地而言, 海洋中关于全新世印度夏季风的研究明显较少。而且与阿拉伯海相比, 东北印度洋的古海洋环境研究明显更少。方念乔等(2001)、丁旋等(2006)曾通过东北印度洋浮游有孔虫的发育情况来探讨印度夏季季风的演化, 认为印度夏季季风显著偏弱的时期基本集中在冰期或冰消期。在安达曼海, Rashid等(2007)通过分析浮游有孔虫的壳体氧同位素和Mg/Ca来估算古温度, 重建了安达曼海25ka BP以来的表层水体温度和盐度变化, 进而探讨了印度季风的强弱变化, 他们认为, 印度夏季风在博令-阿罗德暖期(Bølling-Allerød)和早全新世期间强度比现今明显要强, 而在新仙女木事件(Younger Dryas)和晚全新世期间则相对较弱。然而该孔全新世方面的记录分辨率却较低。黄云等(2017)分析了安达曼海ADM-C1孔全新世高分辨率沉积物粒度资料, 将全新世以来夏季风演化分为三个阶段, 认为中全新世为季风强度最强盛时期。
浮游有孔虫作为生活在上层海洋的一类重要微体生物, 在古海洋环境研究中有着非常重要的作用。浮游有孔虫对海洋环境变化非常敏感, 其群落有明显的纬度分带性(Bé, 1977), 其属种组合和丰度与上层海水的温度、盐度、含氧量等物理化学性质密切相关, 因此, 以浮游有孔虫群落为基础的转换函数和古温度重建曾经广泛应用于全球古海洋环境研究(Thompson, 1981; Kucera et al, 2005; 向荣等, 2011)。在冲绳海槽和南海, 浮游有孔虫群落对末次冰期以来千年尺度的环境变化事件均有很好的响应(Li et al, 1997; Xiang et al, 2007, 2009)。然而在安达曼海, 这方面的研究却很少开展。本研究选取位于安达曼海的ADM-C1孔(图1), 该孔位于研究印度夏季风较理想的区域。通过对该区域浮游有孔虫的群落分析, 来探讨区域海洋环境对全新世呈阶段性变化的印度夏季风的响应。
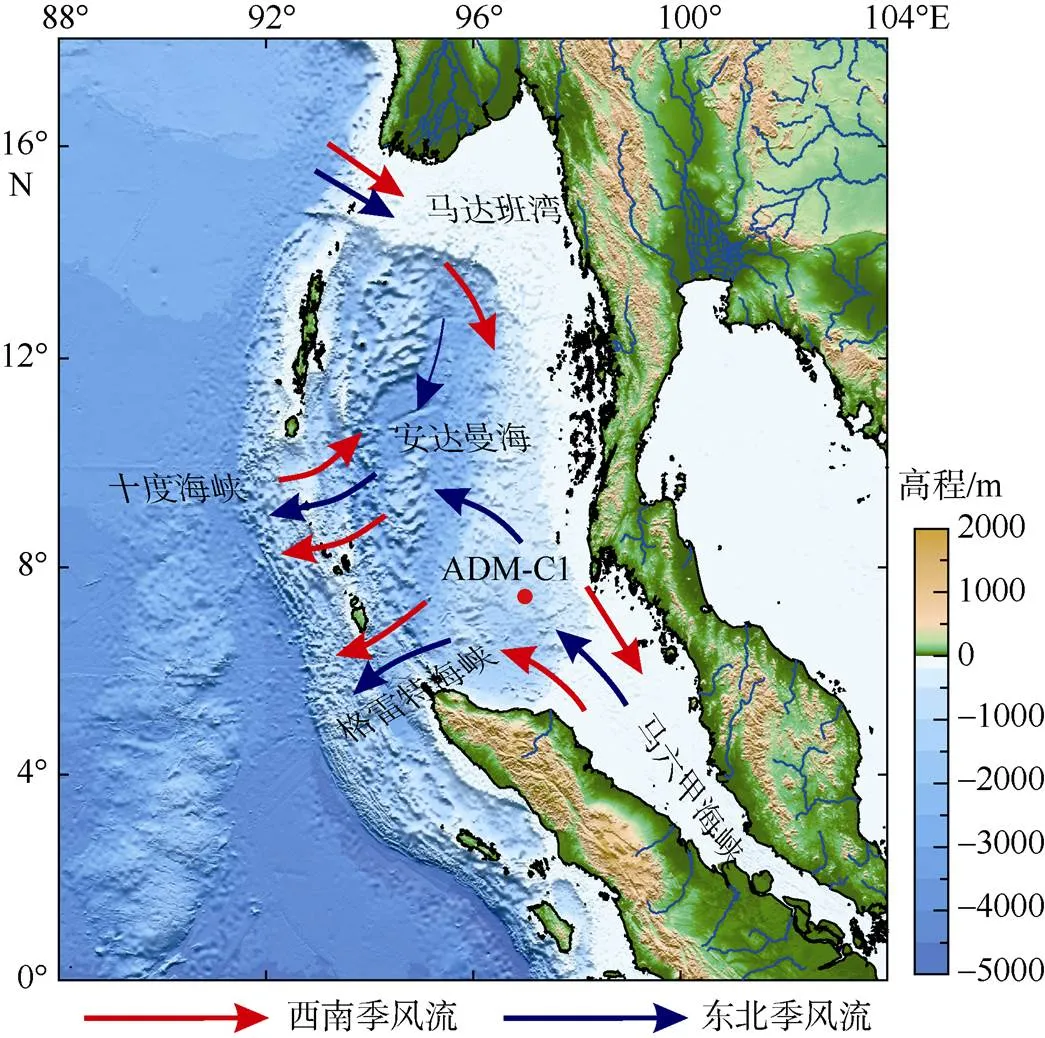
图1 研究区海流模式与取样站位(改自Cao et al, 2015)
1 区域背景
安达曼海位于东北印度洋, 以安达曼群岛和尼科巴列岛为界区分于孟加拉湾, 右侧是马来半岛, 西边通过格雷特海峡、十度海峡和普雷帕丽斯海峡与孟加拉湾进行水体交换, 在东南方向通过马六甲海峡与南海进行海水交换(Varkey et al, 1996)。整个安达曼海面积约8×105km2, 最深处水深可达4400m。受印度季风降水和河流径流的影响, 安达曼海的水体盐度明显比开放大洋低, 并且具有明显的季节变化。安达曼海的径流补给主要来自于陆上的伊诺瓦底江和萨尔温江, 伊诺瓦底江和萨尔温江入海径流和陆输运源物质在安达曼海北部马达班湾经入海口汇入安达曼海。伊洛瓦底江每年输入安达曼海的淡水大约有428km3(Milliman et al, 1983), 主要径流出现在夏季到晚秋, 峰值出现在10月, 入海径流比降水高峰8月份大约有2个月的滞后(Rashid et al, 2007)。安达曼海表层环流体系主要受印度季风控制并随季节变化。夏季风盛行时期, 由季风引起的季风流自安达曼海西北方向进入安达曼海, 与穿越马六甲海峡的西北向海流共同形成一个顺时针涡流, 最终在安达曼海西南部流出; 冬季风盛行时期, 北赤道暖流由马六甲海峡穿过安达曼海到达斯里兰卡南部, 北向和东北向的沿岸流经马来半岛流入孟加拉湾, 形成东北季风流(Brown, 2007; 曹鹏, 2015; 黄云, 2017; Olive, 1994)。
2 材料及方法
研究材料来源于安达曼海ADM-C1重力岩芯(7°26′24″N, 97°E, 水深850m)(图1), 岩芯全长157cm, 沉积物总体上较均一, 以青灰色粉砂质粘土为主。按1cm的间距进行分割取样, 本研究共对44个样品进行了浮游有孔虫群落定量分析。浮游有孔虫分析时, 取大约5g沉积物干样进行处理。首先将样品放在60℃恒温烘箱中烘干, 然后称重, 再将样品放入清水浸泡大约24h, 使样品充分分散。之后将样品用63μm的铜筛冲洗, 将>63μm的部分收集、烘干, 再次称重并计算>63μm的粗组分含量。由于有孔虫氧同位素与Mg/Ca分析过程中, 对浮游有孔虫粒级有一定的要求, 选取的多为250~350μm的有孔虫进行分析测试, 因此为了评价不同粒级有孔虫属种的代表性, 将浮游有孔虫分为>250μm和150~250μm两个粒级进行分析。通过分样器筛分后, 保证每个粒级统计个体在300个以上, 最后将两个粒级组分的样品进行合并, 得到>150μm的有孔虫群落数据。每个样品统计个数均在600个以上。浮游有孔虫鉴定参考了Bé(1977)的分类鉴定标准。
通过对 ADM-C1孔5个不同层位混合浮游有孔虫进行 AMS14C年代测试结果(黄云等, 2017), 结合线性内插法获得其他层位的年龄数据。年龄数据均通过Marine13放射性碳数据库进行了日历年代校正, 5个年代控制点(31、61、93、125、157cm)的日历年龄分别为 3220、5485、6735、8620和11245a。从沉积速率可以看出, 8.6—5.4ka期间沉积速率在全孔最高, 为20.5cm·ka–1; 8.6ka BP之前的早全新世为12.2cm·ka–1; 5.4ka BP之后的晚全新世为11.1cm·ka–1, 较中全新世均明显偏低。
3 结果
3.1 浮游有孔虫的群落结构特征
ADM-C1孔沉积物中含有丰富的浮游有孔虫, 在大于150μm粒级中, 平均1g干样中含835个浮游有孔虫。该孔有孔虫绝对丰度在4.6ka BP出现高值, 浮游有孔虫粗组分与浮游有孔虫丰度变化趋势(图2)较为一致, 在5—6ka BP之前相对较低, 随后相对较高。浮游有孔虫丰度与粗组分的这种变化可能受到了陆源沉积物质稀释作用的影响。从该孔沉积速率的变化可以还看出(图2), 5ka BP之前的沉积速率相对较高, 而其后沉积速率明显降低, 表明了有孔虫丰度变化主要受沉积速率的影响。在该孔共鉴定出22种浮游有孔虫, 为典型的亚热带群落。包括以及。在粒径>150μm的群落组合中, 浮游有孔虫以(44.3%)、(16.3%)、(11.9%)、(8.1%)、(4.03%)、(3.43%)和(3.29%)为主, 以上7种约占浮游有孔虫群落总量的91.4%(图2)。生活在浅层喜光的浮游有孔虫和的相对百分含量最大。广温广盐种相对丰度从早全新世最高到晚全新世以来逐渐降低, 而同为暖水浅层种的则与之呈大致反向的变化趋势(图2)。ADM-C1柱样中相对丰度从11—7.9ka BP较高, 到7.9—3.8ka BP相对丰度有所降低, 再到3.8ka BP以来逐渐升高, 而的变化趋势则与相反, 早全新世相对丰度偏低, 中全新世为最高时期, 晚全新世有所降低。浮游有孔虫和的相对丰度变化趋势与大体相似, 在11—7.9ka BP期间相对丰度相对较低, 随后逐渐增大, 7.9—3.8ka BP达到高值并较为平稳, 3.8ka BP以后明显降低(图2)。过渡带属种的含量变化在11ka BP前最高, 其后至3.8ka BP一直处于含量较低阶段, 大约3.8ka BP以来, 其含量呈逐渐增加的趋势。浮游有孔虫粗组分与浮游有孔虫丰度变换趋势(图2)较为一致, 在11—7.9ka BP期间为低值, 随后逐渐增大, 7.9—3.8ka BP期间为高值, 3.8ka BP以后又明显降低。
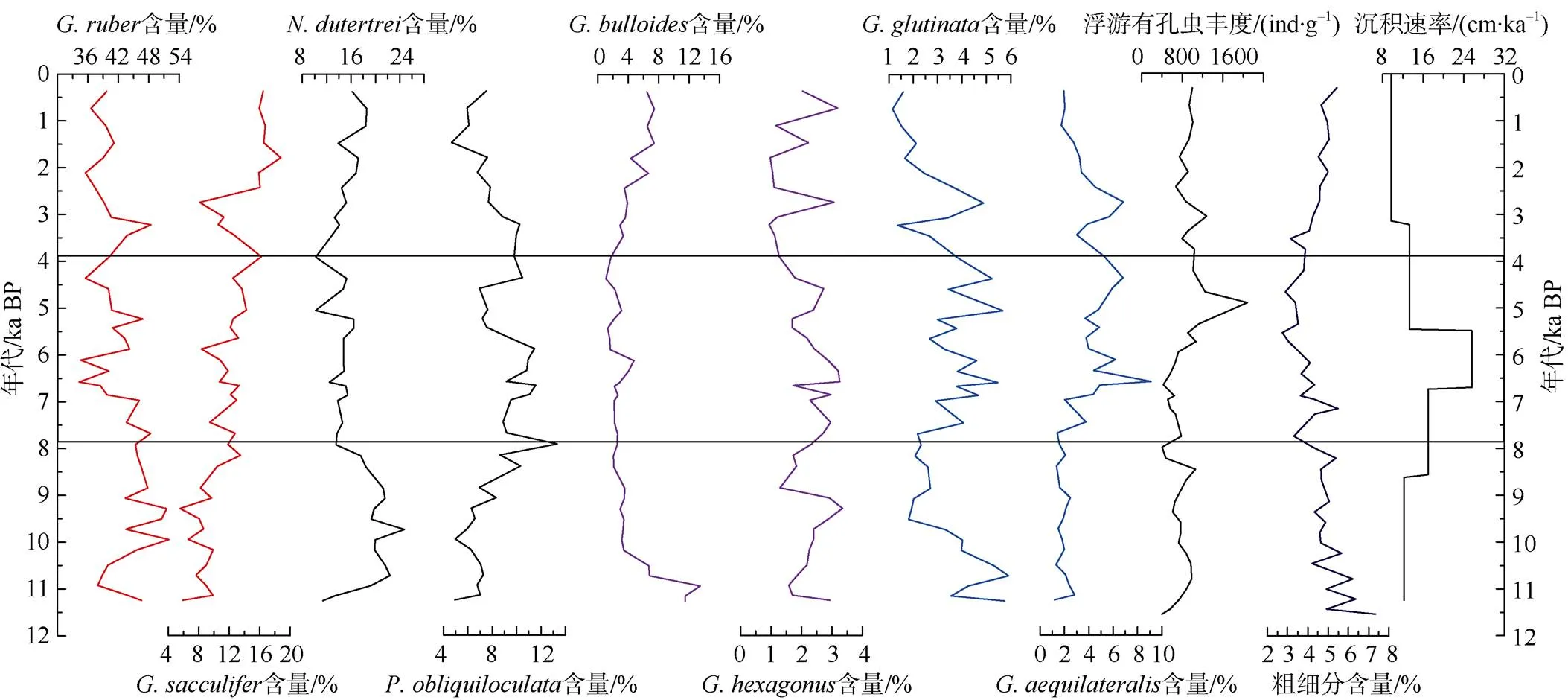
图2 ADM-C1柱样中粒径>150μm的浮游有孔虫主要属种相对丰度和>63μm的粗组分百分含量及浮游有孔虫总丰度与沉积速率变化
>250μm的粗粒级中浮游有孔虫群落同样呈现出明显的变化(图3), 浮游有孔虫主要以(22.7%)、(18.77%)、(18.76%)、(18.53%)、(4.36%)、(4.05%)、(3.86%)、(3.50%)和(3.1%)为主, 这9个属种约占浮游有孔虫群落的97%。在>250μm的粒级中,的相对含量从11ka BP至今呈逐渐增加的趋势,的含量在8ka BP前相对较高, 之后呈逐渐下降的趋势。大个体浮游有孔虫和在大约7ka BP后呈现逐渐增加的趋势, 在3.8ka BP后含量最高。大于250μm的浮游有孔虫占所有>150μm粒级有孔虫的19.97%。具体属种占比情况如下:(27.8%)、(8.31%)、(44.8%)、(30%)、(21%)、(79.9%)、(97.5%)。大于250μm的只占>150μm粒级的的不到10%, 因此选取250~350μm的进行氧同位素与Mg/Ca测试可能缺乏代表性。
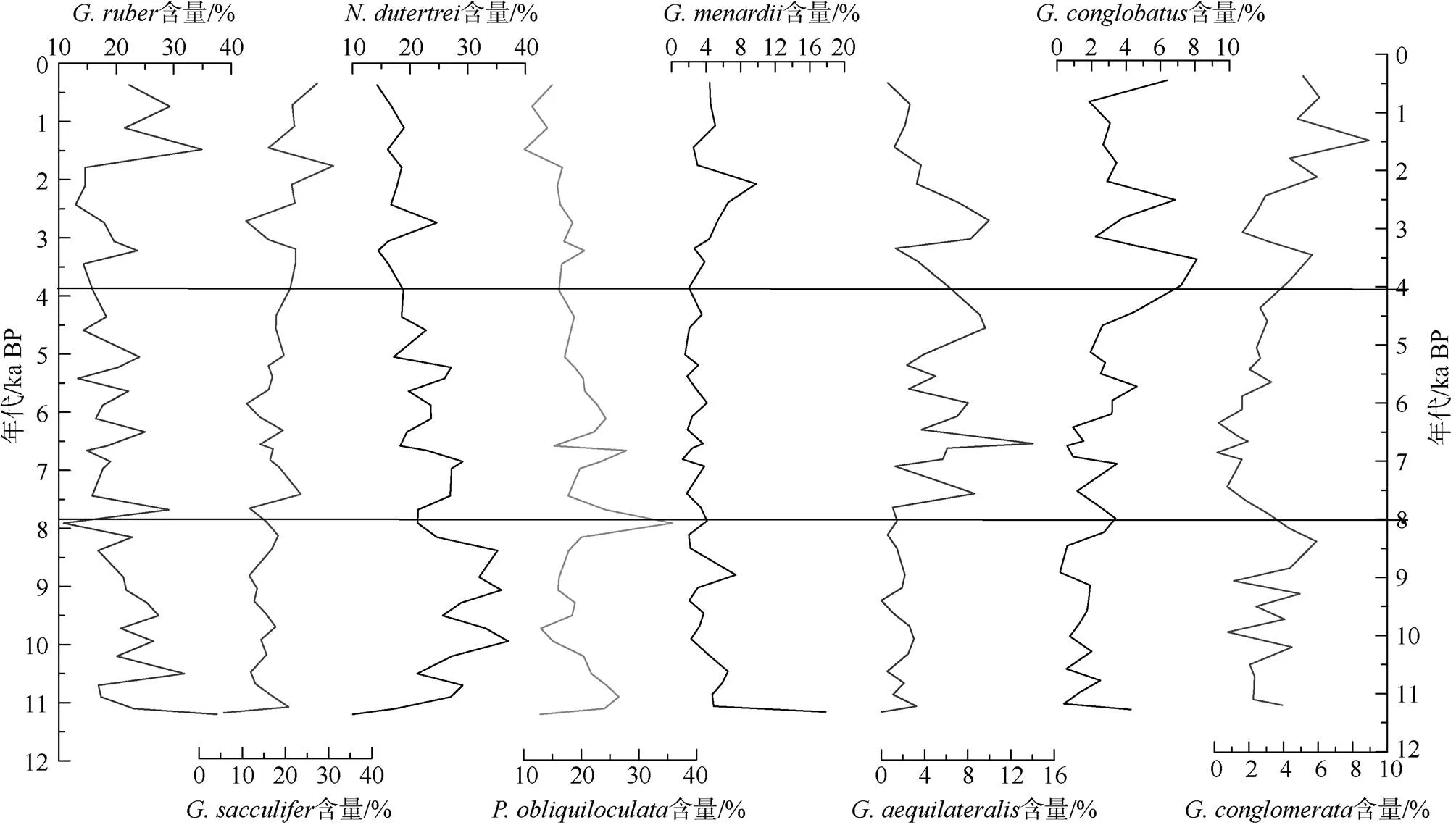
图3 ADM-C1柱样中>250μm浮游有孔虫主要属种的相对百分含量变化
3.2 浮游有孔虫的组合特征
为了更加客观地描述研究区有孔虫属种分布变化, 进一步了解研究区浮游有孔虫与古海洋环境的关系, 对各属种>150μm的百分含量进行了Q型因子分析(Klovan et al, 1971)。分析结果显示, ADM-C1柱样全新世以来的浮游有孔虫群落变化可以分为三个主因子(图4), 这三个因子共解释了总方差的99.2%。主因子1解释了总方差的35.8%, 主要以的因子得分最高(表1), 因子2解释了总方差的28.5%, 该因子和的因子得分最高(表1); 主因子3解释了总方差的34.87%, 该因子载荷主要为负值, 其中.和在该因子上负得分最高, 为该因子的主要贡献属种, 而偏冷环境的和则表现最高正得分(表1)。根据三个主因子的变化特征可以把研究区全新世以来的海洋环境变化划分为3个明显的阶段, 分别为 11—7.9ka BP、7.9—3.8ka BP以及3.8—0ka BP。其中11—7.9ka BP期间主因子1载荷最高; 7.9—3.8ka BP期间主因子3载荷最高, 而3.8ka BP以来则以高的主因子2载荷为特征(图4)。
4 讨论
浮游有孔虫的分布与海洋环境变化有密切的关系, 本文根据对安达曼海ADM-C1孔浮游有孔虫群落及其生态特征探讨浮游有孔虫与古海洋环境演化的联系, 发现热带海区安达曼海浮游有孔虫对古海洋环境有很好的响应。本文群落分析主要讨论>150μm的浮游有孔虫, 经过浮游有孔虫群落统计分析(图2、3)发现, 研究区浮游有孔虫群落以、、、、为主要属种(占84.63%), 为典型热带暖水群落, 同时发现选取250~350μm的进行氧同位素与Mg/Ca测试可能缺乏代表性。总体上, 浮游有孔虫群落变化与全新世环境演化记录有很好的相似性。下面将就安达曼海浮游有孔虫群落全新世阶段性环境变化以及晚全新世普林虫低值事件的响应展开具体讨论。
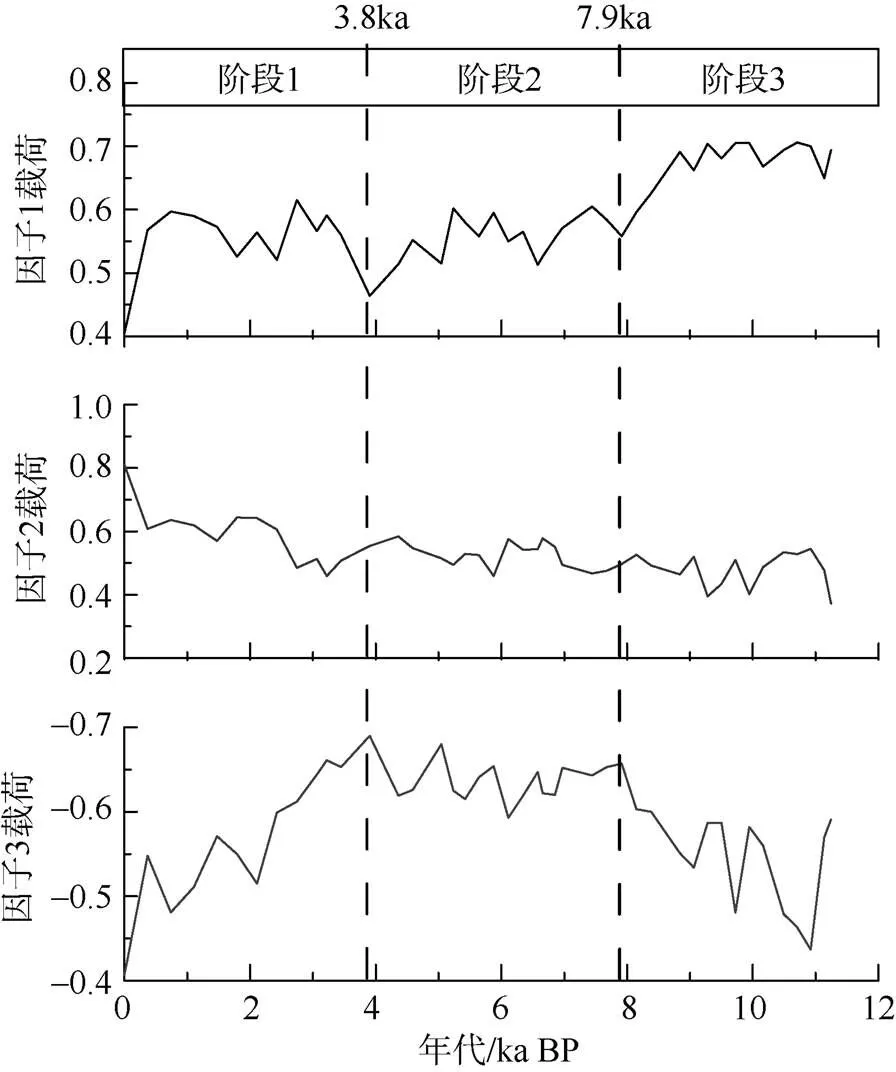
图4 ADM-C1孔中浮游有孔虫3个主因子方差最大化因子载荷随年代变化曲线
4.1 浮游有孔虫群落对全新世环境变化的响应
安达曼海的浮游有孔虫群落在全新世呈现明显的阶段性变化, 结合Q型因子分析结果(图4), 安达曼海全新世古海洋环境可划分为3个阶段: 分别为11—7.9ka BP、7.9—3.8ka BP以及3.8—0ka BP。早全新世11—7.9ka BP期间, 浮游有孔虫以与组合为主。和是该孔最主要两个属种, 虽都属于暖水浅水种, 但生活的温盐环境略有不同。倾向于相对高温高盐环境, 此阶段广温广盐的属种的含量占比最大, 也是整个全新世最高阶段(图3), 可能指示了此阶段表层海水盐度在全新世是相对较低的。这与安达曼海RC12-344孔(Rashid et al, 2007)有孔虫氧同位素记录的表层水体盐度变化一致。此外, 来自陆地石笋的大量记录也指示了早全新世印度夏季风降水明显较强(Fleitmann et al, 2003; Wang et al, 2005; Yuan et al, 2004)(图5)。该时期较强的夏季风降水和入海径流共同造成了早全新世安达曼海表层水体的低盐环境。该阶段另两个主要属种与分别为温带和过渡带特征分子(Bé, 1977), 都偏向生存于温度较低的环境, 且都更加倾向于生活在冬季(Xiang et al, 2015), 其中海水混合强烈的环境更有利于于生存(向荣等, 2011)。同时与这两个属种都与生产力密切相关,在南海被认为与表层生产力密切相关, 是东亚冬季风的良好替代指标(Jian et al, 2003), 而则在阿拉伯海是上升流高生产力的典型指示属种(Anderson et al, 1993)。因此, 这两个属种在11—7.9ka BP的相对高含量可能指示了全新世早期相对较低的表层海水温度或较高的表层生产力水平。
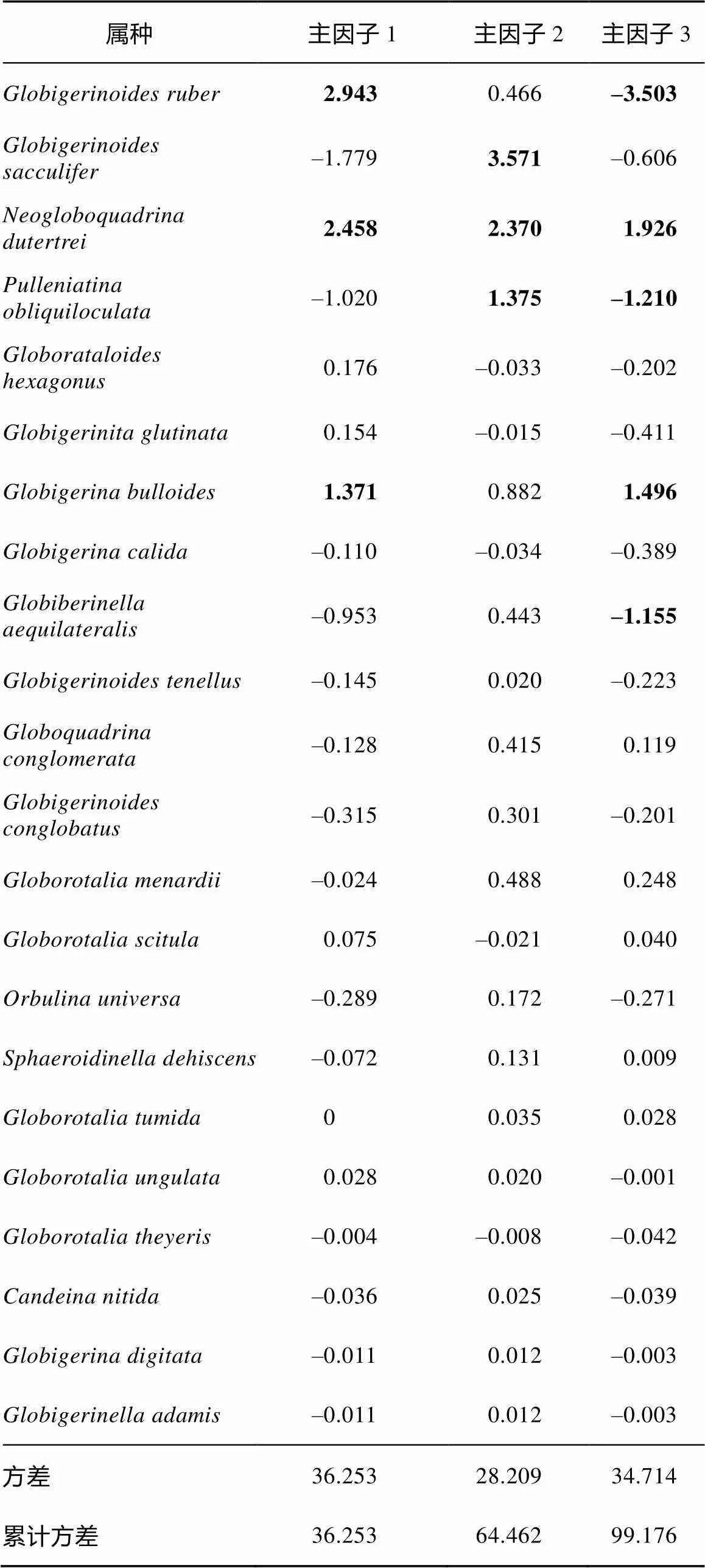
表1 ADM-C1孔中浮游有孔虫方差最大因子得分
注:加粗数字为得分较高属种
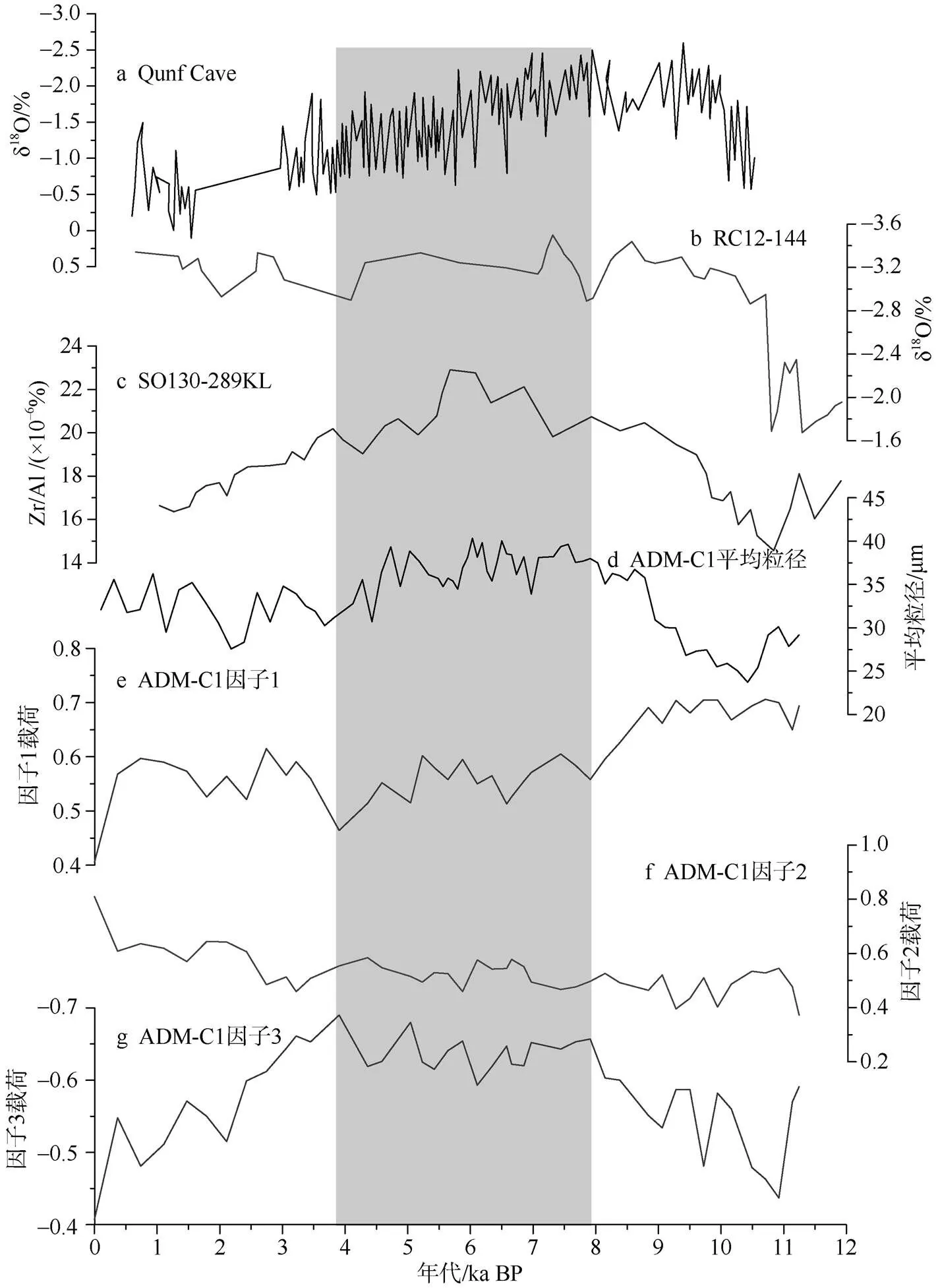
图5 ADM-C1孔中浮游有孔虫群落全新世阶段变化与其他古环境记录对比
a. Qunf cave石笋δ18O值变化(Fleitmann et al, 2003); b. 安达曼海RC12-144孔.δ18O值变化(Rashid et al, 2007); c. SO130-289KL的Zr/Al(Deplazes et al, 2014); d. 主要粒组1.5~11.9μm与11.9~74μm的平均粒径 (黄云等, 2017) ; e—g. ADM-C1孔中浮游有孔虫因子载荷变化。阴影部分为阶段2
Fig. 5 Changes of planktonic foraminiferal in Core ADM-C1 compared with other paleoenvironmental records. a. The records of speleothem δ18O from the Qunf Cave (Fleitmann et al, 2003). b. δ18Oswof Core RC12-344 in the Andaman Sea (Rashid et al., 2007); c. Zr/Al of SO130-289 KL (Deplazes et al, 2014); d. the average grain size variation of grain groups 1.5~11.9 m and 11.9~74 m in ADM-C1 (Huang et al, 2017); e-g. the variation of main planktonic foraminiferal factors in Core ADM-C1. The dark shade indicates phrase 2
中全新世7.9—3.8ka BP期间, 浮游有孔虫以和为特征组合。广温广盐分子相对丰度仍占据主导, 然而其含量相较早全新世阶段有一定下降, 同时含量有所增高(图3), 表明这一阶段表层海水盐度仍处于低值, 但是表层海水盐度与上一阶段相比有所上升, 上升的盐度导致了热带暖水中含量的增加。从石笋反映的夏季风降水(Fleitmann et al, 2003; Yuan et al, 2004; Wang et al, 2005)和安达曼海重建的盐度变化(Rashid et al, 2007)可以看出这一阶段的季风降水仍然较强盛(图6), 与浮游有孔虫记录的海洋环境基本一致。该阶段另两个主要属种分别为和, 其中是世界大洋中重要的喜暖种, 分布于热带—亚热带, 最适宜温度在 22~24℃之间, 系“中层水种”。在冲绳海槽其视为典型的黑潮标志种 ( Jian et al, 2000)。根据现代拖网和捕获器的研究,的通量和相对百分含量在冬季出现高值(陈荣华等, 2006; 向荣等, 2010; Wan et al, 2010)。含量在热带和亚热带上升流或强流区域出现高值(Bé, 1977)。这两个属种都是次表层水属种, 其在中全新世的高含量可能指示该期间较强的上层水体混合(图3)。此外,在此阶段同样存在最高值,的高含量常与上升流带来的高营养相伴(Schiebel et al, 2001)。这也进一步支持了该阶段上层海水混合强烈。这种强烈混合可能和强盛的夏季风风强度对应。来自阿拉伯海的夏季风上升流强度与安达曼海ADM-C1孔的粒度指标指示夏季风风强度均在8.8—4.7ka阶段最强盛(Schulz et al, 1998; 黄云等, 2017), 与浮游有孔虫群落反映的海洋环境有很好的一致性。此外, 来自孟加拉湾的SO130-289KL孔的Zr/Al比值也指示了该时期印度夏季风达到全新世以来最强时期(Deplazes et al, 2014)。同时此阶段粗组分与浮游有孔虫丰度也是全新世最高, 指示生产力最强。
早中全新世安达曼海记录的印度夏季风均显示有较强的降水。然而, 这两个阶段的浮游有孔虫群落却有明显差别, 一个可能的解释是, 反映印度夏季风强弱的另一个指标,即风强度变化在这两个阶段有显著差异。阿拉伯海有机碳含量与印度夏季风引起的上升流密切相关, 较强的夏季风风强导致强盛的上升流发育, 进而导致埋藏在海底的有机碳含量增加。来自阿拉伯海的夏季风上升流强度在早全新世有明显的减弱(Schulz et al, 1998), 这与安达曼海ADM-C1孔的粒度指标指示的这一阶段印度夏季风较弱相一致(黄云等, 2017)。表明早全新世期间印度夏季风的风强度较弱, 而中全新世则显著增强。这种风强度变化导致的海洋环境差异可能是浮游有孔虫群落变化的主要原因之一。
晚全新世3.8—0ka BP期间, 浮游有孔虫以和为主要特征属种。在晚全新世, 浮游有孔虫的减少和含量增加是整个群落变化的重要特征。在这一阶段, 广温广盐分子的相对丰度降至全孔最低, 而热带暖水分子的相对丰度全孔最高, 造成这一变化的主要原因可能与晚全新世明显减弱的夏季风降水密切相关。石笋记录的夏季风降水(Fleitmann et al, 2003; Yuan et al, 2004; Wang et al, 2005)和安达曼海重建的盐度变化(Rashid et al, 2007)均表明这一时期区域降水和入海径流明显减弱, 导致安达曼海表层盐度相较前两个阶段明显升高, 从而有利于较高盐度属种的繁盛。
4.2 全新世普林虫低值事件
普林虫()低值事件是指发生在2.6—4.7ka BP左右的一次普林虫含量明显降低的事件, 该事件在冲绳海槽和南海北部有广泛的记录(Li et al,1997; Ujiié et al, 2003; Lin et al, 2006; Xiang et al, 2007), 被认为是一个可靠的区域年代地层对比依据(李保华等, 2009)。该事件曾被认为是由于黑潮的减弱和东亚冬季风的增强所导致(Jian et al, 2000), 然而事件期间并没有发现明显的古海洋指标异常(Lin et al, 2006)。
在安达曼海, Sijinkumar等(2011)曾报道也有类似普林虫低值事件的存在。他们根据安达曼海的两个沉积物岩芯(SK168和AAS11)中的浮游有孔虫群落分析, 认为在4.5—3ka BP存在一个普林虫含量的低值事件, 可能对应了南海和冲绳海槽全新世普林虫低值事件。此外在YD和20—18ka期间, 他们还认为存在两次普林虫低值事件。
从ADM-C1柱样的有孔虫含量变化(图3)可以看出, 浮游有孔虫的含量变化在2—0.4ka BP和9—11ka BP左右存在有两次相对较低含量阶段, 另外在5ka BP左右也有一次显著的含量降低(图6), 然而这些含量降低事件所发生的时间与Sijinkumar等(2011)曾报道的低值事件无明显的一致性, 尤其在晚全新世阶段。因此, 根据已有的资料, 我们认为安达曼海晚全新世不存在与冲绳海槽和南海一致的普林虫低值事件。
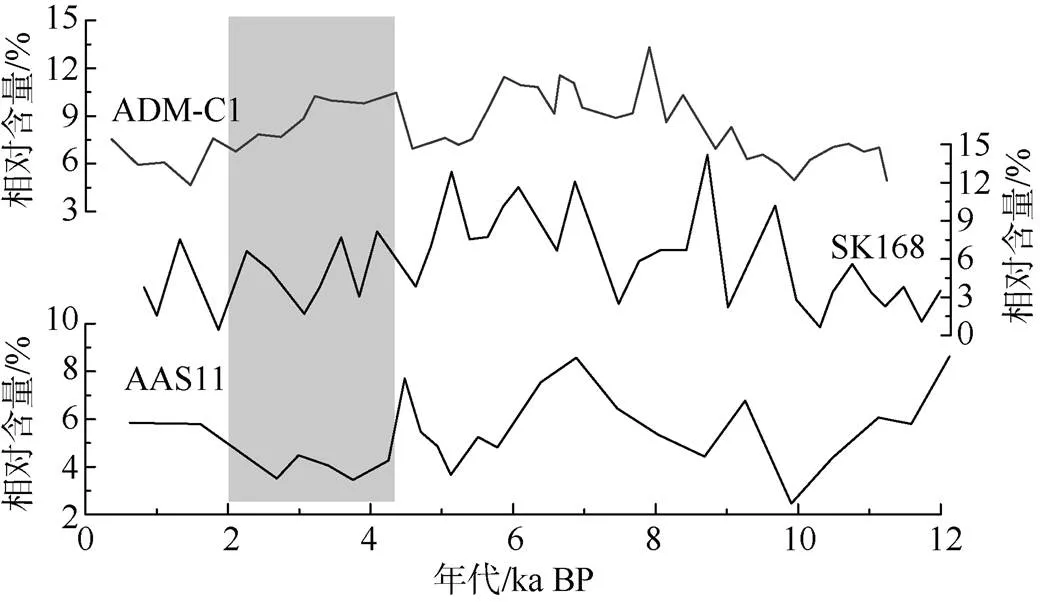
图6 安达曼海三个岩芯ADM-C1、SK168和AAS11 (Sijinkumar et al, 2011)的P. obliquiloculata相对含量对比
阴影为普林虫低值事件
Fig. 6 Abundance ofin Core ADM-C1, SK168 and AAS11 (Sijinkumar et al, 2011)
5 结论
通过对安达曼海的浮游有孔虫群落定量分析, 对比了该区全新世以来的印度夏季风演化记录, 我们认为热带海区的有孔虫群落组合可以很好地响应区域古环境变化。 具体获得以下认识。
1) 浮游有孔虫群落反映了区域海洋环境有明显的阶段变化, 在早全新世11—7.9ka BP期间, 浮游有孔虫群落以与为主, 反映了该阶段低温低盐的水体环境, 可能与较强的夏季风降水和较强的冬季风影响相关; 7.9—3.8ka BP期间,和为主的群落, 指示水体盐度仍然较低, 同时次表层属种显著增多, 可能与该阶段强盛的印度夏季风导致的上层海水强烈混合相关; 3.8—0ka BP期间, 以的含量显著增加, 伴随相对丰度明显下降, 指示区域表层海水盐度显著上升, 对应印度夏季风降水的明显减弱。
2) 通过区域普林虫含量变化的综合对比, 我们认为在安达曼海不存在与冲绳海槽和南海北部类似的晚全新世普林虫低值事件。
曹鹏, 2015. 末次盛冰期以来安达曼海东南部沉积特征及其对印度季风的响应[D]. 青岛: 中国海洋大学. CAO PENG, 2015. The characteristics of the sediment in the southeastern Andaman Sea and its response to the India Monsoon since the Last Glacial Maximum[D]. Qingdao: Ocean University of China (in Chinese with English abstract).
陈荣华, 郑玉龙, WIESNER M G, 等, 2006. 1993~1996年南海中部海洋沉降颗粒通量的季节和年际变化[J]. 海洋学报, 28(3): 72–80. CHEN RONGHUA, ZHENG YULONG, WIESNER M G, et al, 2006. Seasonal and annual variations of marine sinking particulate flux during 1993~1996 in the central South China Sea[J]. Acta Oceanologica Sinica, 28(3): 72–80 (in Chinese with English abstract).
丁旋, 方念乔, 2006. 东北印度洋区BAR9427岩心末次冰期以来的古季风活动记录[J]. 地球科学—中国地质大学学报, 31(6): 765–772. DING XUAN, FANG NIANQIAO, 2006. Records of Paleo-Monsoon of core BAR9427 in northeastern Indian Ocean during Last Glaciation[J]. Earth Science— Journal of China University of Geosciences, 31(6): 765–772 (in Chinese with English abstract).
方念乔, 陈学方, 丁旋, 等, 2001. 孟加拉湾和东径90°海岭260ka以来的古海洋学记录与印度季风的影响[J]. 中国科学 D辑: 地球科学, 31(S1): 280–286. FANG NIANQIAO, CHEN XUEFANG, DING XUAN, et al, 2001. Paleoceanographical records under impact of the Indian monsoon from the Bengal Deep Sea Fan and Ninetyeast ridge during the last 260 ka[J]. Science in China Series D: Earth Sciences, 44(S1): 351–359 (in Chinese).
黄云, 向荣, 刘升发, 等, 2017. 安达曼海沉积物粒度记录的全新世印度洋夏季风演化[J]. 热带海洋学报, 36(6): 19–26. HUANG YU, XIANG RONG, LIU SHENGFA, et al, 2017. Indian Summer Monsoon evolution in the Andaman Sea during the Holocene: evidence from grain size records[J]. Journal of Tropical Oceanography, 36(6): 19–26 (in Chinese with English abstract).
黄云, 2017. 安达曼海全新世以来高分辨率印度洋夏季风演化[D]. 北京: 中国科学院大学. HUANG YUN, 2017. High resolution archive of India summer monsoon variability in the Andaman Sea during the Holocene[D]. Beijing: The University of Chinese Academy of Sciences (in Chinese with English abstract).
李保华, 王晓燕, 2009. 末次冰期以来中国海区浮游有孔虫含量变化的地层学意义[J]. 微体古生物学报, 26(4): 313–322. LI BAOHUA, WANG XIAOYAN, 2009. Variations ofabundance in the china sea since the last glacial period and their stratigraphic significance[J]. Acta Micropalaeontologica Sinica, 26(4): 313–322 (in Chinese with English abstract).
向荣, 陈木宏, 张兰兰, 等, 2010. 南海北部秋季活体浮游有孔虫的组成与分布[J]. 地球科学—中国地质大学学报, 35(1): 1–10. XIANG RONG, CHEN MUHONG, ZHANG LANLAN, et al, 2010. Compositions and distribution of living planktonic foraminifera in autumn waters of the Northern South China Sea[J]. Earth Science—Journal of China University of Geosciences, 35(1): 1–10 (in Chinese with English abstract).
向荣, 刘芳, 孙有斌, 等, 2011. 冲绳海槽末次冰消期以来的浮游有孔虫群落变化及5种古温度转换函数对比[J]. 地球科学—中国地质大学学报, 36(4): 599–609. XIANG RONG, LIU FANG, SUN YOUBIN, et al, 2011. Changes of planktonic foraminiferal assemblages in the Okinawa Trough since the Last Deglaciation and comparisons of sea-surface temperature estimated from five foraminiferal transfer functions[J]. Earth Science—Journal of China University of Geosciences, 36(4): 599–609 (in Chinese with English abstract).
ACHYUTHAN H, NAGASUNDARAM M, GOURLAN A T, et al, 2014. Mid-Holocene Indian summer monsoon variability off the Andaman Islands, Bay of Bengal[J]. Quaternary International, 349: 232–244.
AN ZHISHENG, CLEMENS S C, SHEN JI, et al, 2011. Glacial-interglacial Indian summer monsoon dynamics[J]. Science, 333(6043): 719–723.
ANDERSON D M, PRELL W L, 1993. A 300 KYR record of upwelling off Oman during the Late Quaternary: evidence of the Asian Southwest Monsoon[J]. Paleoceanography, 8(2): 193–208.
BÉ A W H, 1977. An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera[M]//RAMSAY A T S. Oceanic micropaleontology. London: Academic Press: 1–100.
BROWN B E, 2007. Coral reefs of the Andaman Sea-an integrated perspective[M]//GIBSON R N, ATKINSON R J A, GORDON J D M. Oceanography and marine biology an annual review. London: Taylor & Francis 45. 173–194.
CAO PENG, SHI XUEFA, LI WEIRAN, et al, 2015. Sedimentary responses to the Indian Summer Monsoon variations recorded in the southeastern Andaman Sea slope since 26 ka[J]. Journal of Asian Earth Sciences, 114: 512–525.
CHEN FAHU, CHEN XUEMEI, CHEN JIANHUI, et al, 2014. Holocene vegetation history, precipitation changes and Indian Summer Monsoon evolution documented from sediments of Xingyun Lake, South-West China[J]. Journal of Quaternary Science, 29(7): 661–674.
DEPLAZES G, LÜCKGE A, STUUT J B W, et al, 2014. Weakening and strengthening of the Indian monsoon during Heinrich events and Dansgaard-Oeschger oscillations[J]. Paleoceanography, 29(2): 99–114.
DYKOSKI C A, EDWARDS R L, CHENG HAI, et al, 2005. A high-resolution, absolute-dated Holocene and deglacial Asian monsoon record from Dongge Cave, China[J]. Earth and Planetary Science Letters, 233(1–2): 71–86.
FLEITMANN D, BURNS S J, MUDELSEE M, et al, 2003. Holocene forcing of the Indian monsoon recorded in a stalagmite from Southern Oman[J]. Science, 300(5626): 1737–1739.
JIAN ZHIMIN, WANG PINXIAN, SAITO Y, et al, 2000. Holocene variability of the Kuroshio Current in the Okinawa Trough, northwestern Pacific Ocean[J]. Earth and Planetary Science Letters, 184(1): 305–319.
JIAN ZHIMIN, ZHAO QUANHONG, CHENG XINRONG, et al, 2003. Pliocene–Pleistocene stable isotope and paleoceanographic changes in the northern South China Sea[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 193(3–4): 425–442.
KLOVAN J E, IMBRIE J, 1971. An algorithm and FORTRAN-IV program for large-scale-mode factor analysis and calculation of factor scores[J]. Journal of the International Association for Mathematical Geology, 3(1): 61–77.
KUCERA M, WEINELT M, KIEFER T, et al, 2005. Reconstruction of sea-surface temperatures from assemblages of planktonic foraminifera: multi-technique approach based on geographically constrained calibration data sets and its application to glacial Atlantic and Pacific Oceans[J]. Quaternary Science Reviews, 24(7–9): 951–998.
LI BAOHUA, JIAN ZHIMIN, WANG PINXIAN, 1997.as a paleoceanographic indicator in the southern Okinawa Trough during the last 20,000 years[J]. Marine Micropaleontology, 32(1–2): 59–69.
LIN Y S, WEI K Y, LIN I T, et al, 2006. The HoloceneMinimumEvent revisited: geochemical and faunal evidence from the Okinawa Trough and upper reaches of the Kuroshio current[J]. Marine Micropaleontology, 59(3–4): 153–170.
MILLIMAN J D, MEADE R H, 1983. World-wide delivery of river sediment to the oceans[J]. Journal of Geology, 91(1): 1–21.
NEFF U, BURNS S J, MANGINI A, et al, 2001. Strong coherence between solar variability and the monsoon in Oman between 9 and 6 kyr ago[J]. Nature, 411(6835): 290–293.
OLIVE P, 1994. Oceanography and marine biology: an annual review[J]. Marine Pollution Bulletin, 28(1): 56–57.
RASHID H, FLOWER B P, POORE R Z, et al, 2007. A ~25 ka Indian Ocean monsoon variability record from the Andaman Sea[J]. Quaternary Science Reviews, 26(19–21): 2586–2597.
SCHIEBEL R, WANIEK J, BORK M, et al, 2001. Planktic foraminiferal production stimulated by chlorophyll redistribution and entrainment of nutrients[J]. Deep Sea Research Part I: Oceanographic Research Papers, 48(3): 721–740.
SCHULZ H, VON RAD U, ERLENKEUSER H, et al, 1998. Correlation between Arabian Sea and Greenland climate oscillations of the past 110,000 years[J]. Nature, 393(6680): 54–57.
SHEN JI, JONES R T, YANG XIANGDONG, et al, 2006. The Holocene vegetation history of Lake Erhai, Yunnan province southwestern China: the role of climate and human forcings[J]. The Holocene, 16(2): 265–276.
SIJINKUMAR A V, NATH B N, POSSNERT G, et al, 2011.Minimum Events in the Andaman Sea (NE Indian Ocean): implications for winter monsoon and thermocline changes[J]. Marine Micropaleontology, 81(3–4): 88–94.
STAUBWASSER M, SIROCKO F, GROOTES P M, et al, 2003. Climate change at the 4.2 ka BP termination of the Indus valley civilization and Holocene south Asian monsoon variability[J]. Geophysical Research Letters, 30(8): 1425.
THOMPSON P R, 1981. Planktonic foraminifera in the Western North Pacific during the past 150 000 years: comparison of modern and fossil assemblages[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 35: 241–279.
UJIIÉ Y, UJIIÉ H, TAIRA A, et al, 2003. Spatial and temporal variability of surface water in the Kuroshio source region, Pacific Ocean, over the past 21,000 years: evidence from planktonic foraminifera[J]. Marine Micropaleontology, 49(4): 335–364.
VARKEY M J, MURTY V S N, SURYANARAYANA A, 1996. Physical oceanography of the bay of Bengal and Andaman Sea[J]. Oceanography and Marine Biology: An Annual Review, 34: 1–70.
WAN SUI, JIAN ZHIMIN, CHENG XINRONG, et al, 2010. Seasonal variations in planktonic foraminiferal flux and the chemical properties of their shells in the southern South China Sea[J]. Science China Earth Sciences, 53(8): 1176–1187.
WANG YONGJIN, CHENG HAI, EDWARDS R L, et al, 2005. The Holocene Asian monsoon: links to solar changes and North Atlantic climate[J]. Science, 308(5723): 854–857.
XIANG RONG, SUN YOUBIN, LI TIEGANG, et al, 2007. Paleoenvironmental change in the middle Okinawa Trough since the last deglaciation: evidence from the sedimentation rate and planktonic foraminiferal record[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 243(3–4): 378–393.
XIANG RONG, CHEN MUHONG, LI QIANYU, et al, 2009. Planktonic foraminiferal records of East Asia monsoon changes in the southern South China Sea during the last 40,000 years[J]. Marine Micropaleontology, 73(1–2): 1–13.
XIANG RONG, LIU JIANGUO, WANG DONGXIAO, et al, 2015. Seasonal flux variability of planktonic foraminifera during 2009–2011 in a sediment trap from Xisha Trough, South China Sea[J]. Aquatic Ecosystem Health & Management, 18(4): 403–413.
XIAO XIAYUN, SHEN J I, HABERLE S G, et al, 2015. Vegetation, fire, and climate history during the last 18 500 cal a BP in south-western Yunnan Province, China[J]. Journal of Quaternary Science, 30(8): 859–869.
YANG YIPING, ZHANG HUCAI, CHANG FENGQIN, et al, 2016. Vegetation and climate history inferred from a Qinghai Crater Lake pollen record from Tengchong, southwestern China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 461: 1–11.
YUAN DAOXIAN, CHENG HAI, EDWARDS R L, et al, 2004. Timing, duration, and transitions of the last interglacial Asian monsoon[J]. Science, 304(5670): 575–578.
Response of planktonic foraminifera to Holocene marine environmental changes in the Andaman Sea
ZHANG Lingzhi1, 2, XIANG Rong1, TANG Linggang1, 2, YANG Yiping1, ZHONG Fuchang1, 2
1. CAS Key Laboratory of Ocean and Marginal Sea Geology, South China Sea Institute of Oceanology, Guangzhou 510301, China;2. University of Chinese Academy of Sciences, Beijing 100049, China
Through analysis of planktonic foraminifera in a gravity core ADM-C1 from the Andaman Sea, the response of planktonic foraminifera to Holocene marine environmental changes in the sea was discussed. The result shows that the planktonic foraminiferal communities in this region are dominated by tropical warm-water species,,, and. The relative abundance ofshows a gradual decreased trend from the early Holocene to present, which is roughly opposite to that of. The relative abundance ofis higher during 11 to 7.9 ka BP (before present), generally lower during 7.9 to 3.8 ka BP, and then gradually increases after 3.8 ka BP.shows an almost opposite trend to that of. Our result indicates no obviousEventduring the late Holocene, occurs in the Andaman Sea. Q-mode factor analyses of planktonic foraminifera in the core ADM-C1 identify three stages in marine environmental changes during the Holocene. During the early Holocene (11-7.9 ka BP), planktonic foraminifera assemblage was dominated byandwhich reflected that strong summer monsoon and precipitation led to lower sea surface salinity, while the influence of winter monsoon was also strong during this stage. During 7.9-3.8 ka BP,,anddominated, indicating lower sea surface salinity during this period. Meanwhile, there was obviously increase in abundance of subsurface water species, reflecting strong mixing in the upper-ocean waters, possibly due to the strong Indian Ocean summer monsoon at this stage. After 3.8 ka BP,andbecame the dominant species. The abundance ofdeclined obviously, indicating that the salinity of surface sea water significantly increased during this period, which was consistent with the reduced Indian Ocean summer monsoon precipitation. The phase change of the Holocene marine environment reflected by the planktonic foraminiferal assemblages in the Andaman Sea is well consistent with the terrestrial records on the Indian Ocean summer monsoon, and it also shows good consistent with the wind intensity variation in Indian Ocean summer monsoon reflected by sensitive grain size proxy of the same core, suggesting rapid response of planktonic foraminifera to marine environmental changes in the tropical marginal seas.
Holocene; Andaman Sea; Planktonic foraminifera; Indian Ocean monsoon; Marine environment
10.11978/2019010
http://www.jto.ac.cn
P532; Q958.8
A
1009-5470(2019)06-0051-11
2019-01-15;
2019-04-01。
林强编辑
国家自然科学基金项目(41476040, 91228207) ; 中国科学院战略专项项目(XDA11030104)
张玲芝(1993—), 女, 江苏省连云港市人, 硕士研究生。E-mail: zhanglingzhi16@mails.ucas.ac.cn
向荣(1972—), 男, 湖南省洞口县人, 研究员, 主要从事微体古生物与海洋沉积古环境研究。E-mail: rxiang@scsio.ac.cn
2019-01-15;
2019-04-01.
Editor: LIN Qiang
National Natural Science Foundation of China (91228207, 41476040); Strategic Leading Science & Technology Programme, Chinese Academy of Sciences (XDA11030104);
XIANG Rong. E-mail: rxiang@scsio.ac.cn
