我国棉花形态标记性状应用研究进展
2019-11-28陈莹张法铭姜辉柴启超王秀丽高明伟王家宝张超王永翠郑锦秀赵军胜
陈莹 张法铭 姜辉 柴启超 王秀丽 高明伟 王家宝 张超 王永翠 郑锦秀 赵军胜
摘要:本文综述棉花主要形态标记性状的相关应用研究进展,旨在为形态标记性状在棉花遗传育种和分子育种中的应用提供参考。通过对我国目前具备形态标记性状的审定棉花品种进行分类和统计发现,芽黄标记主要被用于与不育基因连锁实现不育系在苗期的早期鉴定选择,有助于提高棉花杂交制种产量和效益,育成品种1个。鸡脚叶标记棉具有良好的冠层结构及光合生理特性,因此具有一定的早熟性、耐旱性和较好的抗病虫性,在三系杂交棉生产上也有较好的应用前景,育成品种3个。红花标记作为非常直观的形态性状标记,可用于简便高效地鉴别真假杂种,其中山东棉花研究中心培育审定了4个红花标记新品种。腺体标记除了是直观的形态性状标记以外,还具有低酚少腺体的特点,山东棉花研究中心培育审定2个低酚标记的抗虫杂交棉新品种,邯郸市农业科学院与河北农业大学审定了3个抗虫低酚棉品种。其他形态标记如窄卷苞叶性状、红叶性状等在杂种优势利用方面也有较好的生产应用潜力。以上各类形态性状标记在棉花生产上均具有广泛的应用前景,需进一步提高标记利用率,加强其应用基础研究。
关键词:棉花;形态标记;芽黄;鸡脚叶;红花标记;腺体标记
中图分类号: S562.03文献标志码: A
文章编号:1002-1302(2019)18-0046-04
收稿日期:2018-10-16
基金项目:国家自然科学基金(编号:31601348);国家现代农业产业技术体系——棉花产业技术体系(编号:CARS-15-05);山东省重点研发计划(编号:2017GNC10114)。
作者简介:陈 莹(1979—),女,山东济宁人,硕士,副研究员,主要从事棉花育种及相关生物学研究及科研管理。E-mail:chenyingkb@126.com。
通信作者:赵军胜,博士,研究员,主要从事棉花育种研究。E-mail:zhaojunshengsd@163.com。
在各种作物遗传育种过程中,标记性状被广泛用于品种纯度鉴定和种质资源的遗传多样性分析。目前,标记主要包括形态标记、细胞学标记、生化标记和DNA分子标记,其中形态标记和DNA分子标记应用较多。DNA分子标记虽不受环境和组织特异性的影响,可客观准确地反映遗传个体间的差异,但是需要借助于实验仪器,限制了其在大田生产中的应用。虽然部分形态标记易受环境影响,但是大部分较为稳定,易于观察,有助于田间的快速有效选择。棉花杂种优势较强,高纯度的杂交种是实现棉花杂种优势利用的重要前提和保证。在大田制种过程中,借助于形态标记可有效去除杂株,保证制种纯度。由此可见,形态标记研究对棉花生产具有重要意义。因此,本文就棉花主要形态标记性状的相关应用研究进展进行综述,旨在为其在遗传育种理论研究和生产中应用提供一定依据。
1 我国棉花形态标记应用研究现状
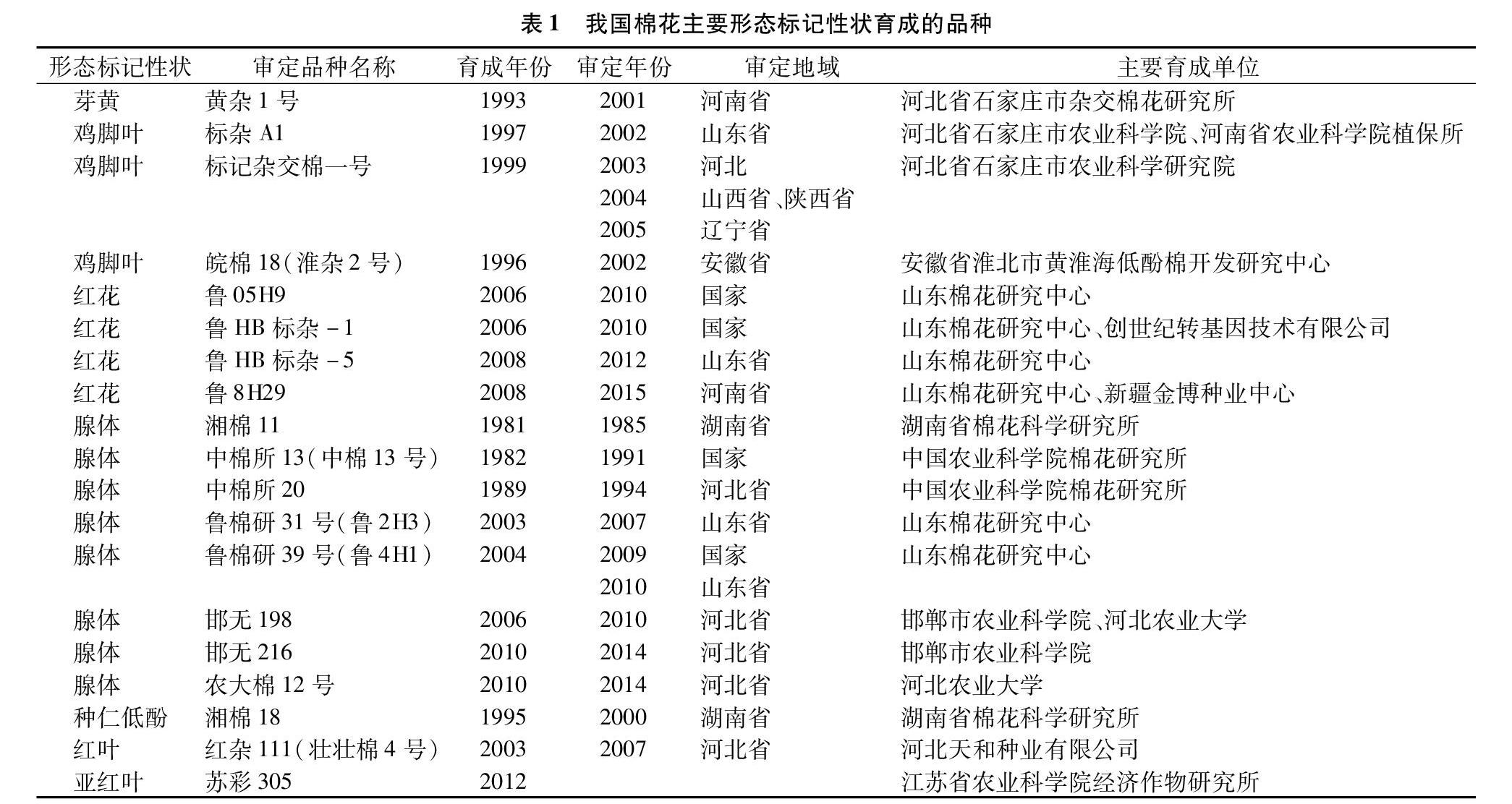
棉花形态标记性状大部分属于质量性状[1],涉及18个性状,其中研究较多的有芽黄、鸡脚叶、红花、腺体标记性状,研究较少的包括茸毛、红株、红叶、窄卷苞叶、黄花药、光籽等标记。在应用层面上可分为2类:第1类,本身仅可作为形态标记,如芽黄、红花、植株茸毛、黄花药等标记;第2类,除了具有标记作用外,还有其他功能性作用,如鸡脚叶、腺体标记等。经不完全统计,目前我国具备主要形态标记性状的育成、审定棉花品种见表1。
2 棉花形态标记性状类型
2.1 芽黄标记
芽黄(virescent)是叶绿素含量较正常稀少的隐性突变,类型较多,主要特点是苗期真叶表现明显的黄色,随着植株生长,黄化叶片随着叶绿素含量的不断增加而逐渐变绿,达到正常水平。芽黄性状表现明显、稳定且容易辨别,遗传方式简单,大多受1对隐性基因控制,是理想的标记性状,在棉花杂种优势利用领域已有很多研究。肖松华等研究发现,同一芽黄基因导入不同遗传背景陆地棉后,对农艺性状影响的范围和程度差异很大[2]。陆地棉芽黄品系与常规品系杂交,F1具有明显的杂种优势,包括产量构成因素、早熟性和纤维品质性状优势,尤其是产量性状优势明显[2-4]。王学德等经亲本配合力分析认为,彭泽芽黄和湖北芽黄是较好的芽黄亲本[5]。潘家驹等鉴定了6个芽黄基因对其转育系经济性状的影响并研究证实了这6个芽黄转育系与陆地栽培品种的杂交组合的产量优势,证明了v10、v15、v20 3个芽黄基因作为标记基因用于杂交棉制种的可能性[6]。
通过芽黄标记与不育基因连锁来实现不育系在苗期的早期鉴定选择,可有效提高棉花杂交制种的产量和效益。其中,81A是携带芽黄标记的陆地棉核雄性不育系,其在子叶期表现为浅黄色,1~3张真叶期表现为正常绿色,4叶期开始表现整株的淡黄色,7叶期后黄色渐褪,而到开花期表现为芽黄类型,即上部几张叶片表现为淡黃色,下部叶片表现为正常绿色,遗传研究证实,其芽黄基因和雄性不育基因紧密连锁,可以用于杂交制种母本筛选[7]。宇文璞等引入81A后,对芽黄A的温敏感特性进行了系统研究,发现并提出了芽黄A的温控繁育法,选育出在子叶至3张真叶期就具有准确芽黄标记性状、杂种优势显著的温敏性“芽黄A”雄性不育系;以此转育成的芽黄8748A不育系配制的杂交种大部分组合优势明显,产量高,抗逆性强,其中用2145作父本配制的黄杂1号,表现出早发早熟、有效铃多、铃大、衣分高、增产明显、稳产性强、纤维品质好等优良性状,生产示范应用综合表现突出[8-9]。1998年湖南农业大学发现了高温敏感型棉花雄性不育材料,培育出稳定的棉花温敏雄性不育系特棉S-1,其育性在日平均温度<26.5 ℃时表现为可育,>27.0 ℃时表现为不育[10-11]。
棉花光温敏雄性不育系是从海陆杂交种后代中发现的;具有芽黄标记性状的新型核雄性不育系,是对芽黄A经过30年40多代选育出的棉花雄性不育系新类型[12-13]。芽黄396A是携带连锁芽黄基因的棉花单隐性光温敏核雄性不育系,品质和农艺性状优良,在高温长日照或低温短日照条件下表现为完全雄性不育,而在高温短日照条件下表现为可育,可自交成铃,不再需要专门的保持系,该不育系携带的芽黄标记,在正常条件下,于出苗5~7 d的子叶期即可明显区别可育株与不育株,在非正常条件下,最晚到3~5张真叶期即可区别[14]。牛金生等在新疆和三亚对396A的育性变化及其杂交种的产量性状、抗病性、纤维品质等开展试验研究,发现其杂交种F1代育性恢复好,花粉量大,全部能够自然成铃且杂种优势明显[15]。张涛等对影响棉花光温敏不育A系杂交成铃的4个方面因素进行分析研究,确定了杂交授粉的最佳授粉日期、时间段和日平均气温,发现以不同不育系为母本时的杂交成铃率之间存在明显差异[16]。
2.2 鸡脚叶标记
鸡脚叶是陆地棉正常阔叶的显性突变类型之一,具有裂刻极深、叶裂片窄的特点。鸡脚叶标记棉单株和群体通风透光性好,因此具有良好的冠层结构及光合生理特性。朱伟研究发现,鸡脚叶叶面积比正常叶缩小12.94%~25.59%,大大改善了棉田光照条件[17]。同鸭掌叶比较,鸡脚叶叶绿素含量增加12.77%~39.36%,光合作用强度高17.5%,光照度增加12.77%~39.36%,透光度增加12.90%~39.45%,且中下层增加趋势更明显[17-18]。杜明伟等研究发现,鸡脚叶棉上部冠层光吸收率为38%~47%,中下部冠层为22%~31%,整体冠层的光分布均匀;而正常阔叶棉上部冠层光吸收率为65%~70%,极显著高于中下部;鸡脚叶棉冠层群体光合速率分布比正常阔叶棉更均衡,特别是在盛铃后期,鸡脚叶棉上部、中部、下部冠层的群体光合速率分别占总体光合速率的40.9%~46.7%、26.8%~29.6%、18.5%~21.5%,而正常阔叶棉则分别占61.1%~72.3%、21.7%~25.9%、102%~15.3%[19-20]。
鸡脚叶棉还具有一定的早熟性、耐旱性。鸡脚叶棉的果节出现速率比普通叶棉高50%~100%,开花前期每天开花3.9朵,而正常阔叶棉每天开花1.8朵;鸡脚叶棉的霜前花率为81.8%,比普通叶棉高12.3%[21-22]。鸡脚叶的蒸腾系数小于正常阔叶,可使叶片在干旱状态下处于有利保持水分的状态[23-24]。用鸡脚叶(或超鸡脚叶)作亲本配制的杂交棉也显示了早熟性,如标杂A1在河南省区试中,霜前皮棉产量比对照极显著增加36.6%[17]。
由于综合了形态抗性、生态抗性和生理生化抗性,鸡脚叶棉花一般抗棉铃虫、红铃虫、棉大卷叶螟、斜纹夜盗蛾、抗叶螨等[17,25]。鸡脚叶品种棉田透光性好、田间湿度低,所以喜湿或喜郁蔽条件的黏虫、玉米螟等害虫的生长发育也受到抑制[26];鸡脚叶棉对角斑病的18个生理小种均具有抗性[25],极大地限制了棉铃疫病的发生、发展[17]。另外,鸡脚叶有利于农药穿透,能大大提高化学防治病虫害效果;鸡脚叶棉花一般较矮,株型紧凑,能抗风暴[25]。
美国在1976、1981年先后发放了Gumbo等鸡脚叶陆地棉品种[27]。澳大利亚从1984年开始发放育成的鸡脚叶品种,至20世纪90年代,鸡脚叶品种种植面积已经达到澳大利亚植棉面积的45%以上,主要种植品种有Siokral-4、Siokra L23、Siokra S324、SiokraV1等[25]。我国曾育成超鸡脚叶川简3号、鸡脚紫絮棉等品种,鸡脚德字棉在四川省推广种植10多万hm2。目前,通过省级审定的品种中包括3个杂交种——通过河南省、山东省审定的标杂A1[28],通过河北省、山西省、陕西省、辽宁省4个省审定的标记杂交棉一号[29],通过安徽省审定的皖棉18(淮杂2号)[30]等。朱伟以超鸡脚叶、鸡脚叶和正常叶棉花不育系及其恢复系为杂交亲本配制组合,验证发现,杂种F1代由于单铃质量和衣分增加出现皮棉增产342%(增产极显著)的情况,表明鸡脚叶标记在三系杂交棉生产上具有广泛的应用前景;另外发现,鸡脚叶杂交种双亲自交系可增强优势,通过自交系配组可使其功能充分展现[17]。
2.3 红花标记
20世纪80年代,由原中国科学院遗传研究所牵头的棉花远缘杂交协作组,用陆地棉与比克氏棉进行远缘杂交,选育出了具有粉红色花瓣和红色大基斑的红花纯合材料,定名为“HB(hirsutum-bickii)红花系”[31]。赵军胜等利用4个陆地棉HB红花近等基因系作父本,分别与轮回亲本杂交组合,研究红花标记性状棉花在产量杂种优势中的表现,同时将HB红花近等基因系及轮回亲本作父本分别与共同母本杂交,比较各组F1有关性状及产量等方面的超亲、平均及竞争优势。结果表明,HB红花标记杂交组合籽棉与皮棉产量具有明显的杂种优势,野生二倍体比克氏棉的渐渗片段对陆地棉杂种优势有重要的正向效应[32]。山东棉花研究中心通过组配强优势杂交组合,育成并审定了鲁05H9(国审棉2010003)、鲁HB标杂-1(国审棉2010004)、鲁HB标杂-5(鲁农审2012020号)、鲁8H29(豫审棉2015010)等多个红花标记转基因抗虫杂交棉[33-34]。
2.4 腺体标记
色素腺体为棉族植物特有,是棉属植物分类的依据之一。棉花的色素腺体不仅是棉酚贮藏场所,还与棉酚的合成有着密切关系,但在不同类型的棉花之间,色素腺体与棉酚存在不同相关性[35]。棉酚对棉花抵抗某些病虫害有重要作用,但是不利于种子营养成分的充分利用,因此,选育植株有腺体而种子无腺体或者有腺体无酚的品种是解决这一问题的有效方法。
腺体标记可用于真假杂种的简便辨认。袁有禄等利用显性无腺体陆地棉新品系N1为父本,以有腺体中棉所19和有腺体MS5MS6核不育系为母本,进行不去雄人工辅助授粉,在后代中利用腺体标记对真假杂种进行分辨发现,获得真杂种率分别为52.4%和99.3%[36]。
棉酚对单胃动物具有毒害,选育腺体标记的低酚棉品种是提高棉籽利用率的关键。我国的低酚棉育種研究开始于1972年,20世纪90年代达到高峰,当时全国棉区育成不同类型低酚棉品种近60个,推广面积也快速增长;至1987年,我国低酚棉种植面积已经超过美国和科特迪瓦共和国,居世界第1位,至1991年种植面积一直稳定在10万hm2以上,且以河北省、河南省、山东省棉区种植面积最大[37]。喻树迅等成功选育了具有纯合双隐性基因gl2gl2gl3gl3的低酚棉品种中棉所20,该品种总棉酚含量仅为0.002 5%,远低于世界粮农组织和卫生组织规定的0.04%的标准,而且还具有柱头短、花丝长、散粉早的特点,可大大降低异交率,在不隔离种植的条件下,纯度可达98%以上[38]。祝水金等培育了具有子叶腺体延缓形成特性的陆地棉种质系ABH-0318,该材料色素腺体性状稳定,休眠种子仅在子叶边缘有少量腺体,棉酚含量仅为0.017 5%;种子萌发后,子叶和植株腺体数量与一般有酚品种相似,为培育植株有腺体而种子无腺体的棉花品种奠定了基础[39]。山东棉花研究中心育成了具有低酚标记性状的转基因抗虫杂交棉品种鲁棉研31号(山东省审定)和鲁棉研39号(国家审定、山东省审定),并建立了鲁棉研39号的指纹图谱[40]。2010年河北省也首次审定了抗虫低酚棉品种邯无198,2014年育成审定了邯无216、农大棉12号2个抗虫低酚棉[37,41]。刘芳等在新疆巴州对47个低酚棉品种资源的主要性状进行了调查分析,优选了11个适宜的资源材料[42]。李函利等对179份不同来源的低酚棉种质资源的耐盐性进行了评价,筛选出8份达到了耐盐和抗盐水平的突出材料,为低酚棉育种筛选到了优异种质资源[43]。刘慧等研究了不同基因型棉花不同器官的色素腺体与棉酚旋光体含量间的相关性,结果发现,低酚棉虽全株无色素腺体,但器官中也能检测到低于0.02%的少量游离棉酚存在,有酚棉棉仁中色素腺体在各器官中最密,花瓣和棉仁色素腺体直径最小,且棉仁和花瓣的总棉酚含量存在高度相关性,因此可以用花瓣的棉酚含量来判断棉仁的棉酚含量[44]。
2.5 其他标记
窄卷苞叶(Frego bract)是由美国阿肯色州棉农Frego发现的,具有苞叶狭长、革质化、向外扭翻生长,使花冠和棉铃充分暴露的特性,受1对隐性基因(fg)控制。张金发等利用鄂沙78和7010 2个陆地棉品种作为母本,分别与窄卷苞叶棉Frego杂交组成1对近等基因系,与正常苞叶近等基因系试验比较,窄卷苞叶棉叶片光合速率下降、生长旺盛、不易早衰,对棉花產量、衣分、单株果枝数、脱落率和纤维品质无明显不良效应,在现蕾之前延迟棉花生育期,影响早熟,导致现蕾开花推迟,且对棉铃象鼻虫、棉铃虫等有一定的抗性[45-47]。
李俊兰等在研究红叶观赏棉花的同时,提出利用红叶作为标记性状应用于陆地棉杂种优势研究[48]。2006年狄佳春等报道了陆地棉亚红株突变体,并对其光合特性进行了初步研究[49]。王忠义等于2007年育成红叶棉花品种红杂111,霜前皮棉比DP99B增产11.3%[50]。万艳霞等利用3个红叶材料作父本,与15个绿叶棉花品种组配19个红叶杂交组合,对具有红叶标记性状棉花的产量杂种优势进行研究发现,19个红叶杂交组合籽棉产量均具有明显的杂种优势,皮棉产量具有正向中亲优势、正向超亲优势和负向竞争优势,衣分和铃质量具有正向的中亲优势,单株铃数表现正向中亲、超亲和竞争优势,并筛选出了4个产量竞争优势在5%以上、综合性状较好的优势组合[50]。陈旭升等选育出了具有亚红叶标记的优质高产彩色杂交棉组合苏彩305,在生产上具有推广潜力[51]。
为了克服胞质雄性不育恢复系恢复基因鉴定难、保存难的问题,刘英新等选育了丛生铃、鸡脚叶、海岛棉和无腺体4个标记性状恢复系,并进行了产量、品质和光合生理特性比较,发现它们在三系杂交棉育种、制种和繁种中都具有良好的利用价值[52]。
3 问题与展望
棉花形态标记性状在研究与应用方面取得了一定成果,特别是在育种应用研究方面成效显著,已经培育、选育出了一些具有标记性状的优良种质材料或品种,推动了棉花杂种优势在生产上的应用。但目前形态标记性状的应用还存在以下2个问题:
(1)形态标记性状利用率偏低。棉花具有丰富的形态标记性状,目前应用比较成功的只有芽黄标记、红花标记及腺体标记,而大部分处于未开发状态。因此,需要对形态标记进行进一步的深入发掘,同时修改现行部分试验方法,促进形态标记的利用。(2)针对标记性状的基础研究相对薄弱。相对于应用研究,针对棉花形态标记性状遗传特性解析及相关基因功能分析方面的研究较少,而且不够深入,这也是阻碍形态标记性状在棉花育种中应用的因素。随着棉花基因组序列的公布,这一问题已逐步得到解决,受到广泛关注的如色素腺体性状标记的遗传基础研究已经有了一定突破,但其他性状标记的分子机制和遗传研究都还比较滞后,有待进一步深入挖掘。
基因工程和生物技术的不断发展,势必会推进棉花形态性状标记的基础和应用研究更加深入,促进棉花产业与其他产业的结合,不断简化生产成本,提高植棉收益,带动植棉积极性,进而有利于我国棉花产业的可持续发展。
参考文献:
[1]承泓良. 棉花质量性状遗传研究进展[J]. 江苏农业科学,1984(7):46-49.
[2]肖松华,张天真,潘家驹. 陆地棉芽黄近等基因系研究[J]. 棉花学报,1996,8(5):235-240.
[3]肖松华,黄骏麒,潘家驹,等. 陆地棉芽黄品系和常规品种间杂种优势利用研究[J]. 棉花学报,1996,8(2):71-76.
[4]阂留芳,何金龙,肖松华,等. 陆地棉芽黄品系在棉花杂种优势上的利用研究[J]. 棉花学报,1996,8(3):113-119.
[5]王学德,潘家驹. 陆地棉芽黄指示性状的杂种优势利用研究[J]. 南京农业大学学报,1989,12(1):1-8.
[6]潘家驹,闵留芳,刘 康,等. 陆地棉芽黄基因应用于杂种棉的研究[J]. 南京农业大学学报,1998,21(3):7-14.
[7]张天真,潘家驹,冯福祯. 一个有芽黄标记性状的棉花雄性不育系的遗传鉴定[J]. 中国农业科学,1989,22(4):17-21.
[8]宇文璞,宇文纲,王淑霞,等. 温敏型隐性核不育棉花应用研究初报[J]. 河北农业科学,1994(4):38,24.
[9]宇文璞,宇文纲,王淑霞,等. 棉花温敏型核不育芽黄A生产利用[J]. 中国棉花,1996,23(9):28.
[10]余筱南,陈金湘,李瑞莲,等. 棉花温敏雄性不育系的选育与应用研究简报[J]. 棉花学报,2003,15(6):380-381.
[11]唐海明,余筱南,陈金湘,等. 棉花温敏雄性不育系特棉S-1的利用[J]. 湖南农业大学学报(自然科学版),2005,31(5):30-33.
[12]王凯辉,郭宝生,刘素恩,等. 棉花光温敏核雄性不育系育性及其杂种优势研究[J]. 河北农业科学,2013,17(3):55-59.
[13]宇文小岗,王淑霞,宇文璞,等. 棉花雄性核不育系光温A系简介[J]. 中国棉花,2006,33(11):18.
[14]张建丰,张战奇,宇文璞. 棉花单隐性光温敏细胞核雄性不育系芽黄396A[J]. 农业科技通讯,2006(4):33.
[15]牛金生,韩光明,蓝家样,等. 棉花光温敏核不育系396A育性特征及杂种优势分析[J]. 棉花科学,2016,38(6):7-12.
[16]张 涛,孟庆忠,王玉美,等. 棉花光温敏雄性核不育系杂交成铃率的影响因素研究[J]. 农村经济与科技,2017,28(增刊1):23-24.
[17]朱 伟. 鸡脚叶标记的三系杂交棉杂种优势机理的研究[D]. 杭州:浙江大学,2006.
[18]吴巧娟,肖松华,刘剑光. 鸡脚叶对陆地棉光合特性的影响[J]. 江西棉花,2007,29(5):11-13.
[19]杜明伟,冯国艺,姚炎帝,等. 杂交棉标杂A1和石杂2号超高产冠层特性及其与群体光合生产的关系[J]. 作物学报,2009,35(6):1068-1077.
[20]冯国艺,罗宏海,姚炎帝,等. 新疆超高产棉花叶、铃空间分布及与群体光合生产的关系[J]. 中国农业科学,2012,45(13):2607-2617.
[21]Andries J A,Jones J E,Sloane L W,et al. Effects of okra leaf shape on boll rot,yield,and other important characters of upland cotton,Gossypium hirsutum L.[J]. Crop Science,1969,9(6):705-710.
[22]Karami E,Weaver J B J. Dry-matter production,yield,photosynthesis,chlorophyll content and specific leaf weight of cotton in relation to leaf shape and colour[J]. Journal of Agricultural Science,1980,94(2):281-286.
[23]Baker D N,Myhre D L. Effects of leaf shape and boundary layer thickness on photosynthesis in cotton(Gossypium hirsutum)[J]. Physiologia Plantarum,1969,22(5):1043-1049.
[24]Karami E,Krieg D R,Quisenberry J E. Water relations andcarbon-14 assimilation of cotton with different leaf morphology[J]. Crop Science,1980,20(4):421-426.
[25]王淑民. 澳大利亚棉业[M]. 北京:中国农业出版社,1995.
[26]王忠义,芦立婷,樊建英. 超鸡脚叶棉花资源育种潜力研究[J]. 中国棉花,2009,36(9):25-26.
[27]张雪林,韩世杰,彭凡嘉,等. 鸡脚叶陆地棉育种研究进展[J]. 中国棉花,2016,43(10):17-20.
[28]王忠义,赵敬霞,王兴三,等. 超鸡脚叶类型棉花的利用价值[J]. 中国棉花,2007,34(11):30-31.
[29]王忠义. 标杂A1和标记杂交棉一号的选育体会[J]. 中国棉花,2005,32(6):38-39.
[30]张家宪,蒋国柱,刘 伟. 高产优质短季低酚杂交棉——淮杂2号[J]. 中国棉花,2002,31(4):19-20.
[31]何鉴星,姜茹琴,张欣雪,等. 比克氏棉渐渗到陆地棉的花色基因的遗传分析[J]. 遗传学报,2000,27(4):344-350.
[32]赵军胜,高明伟,张兴居,等. HB红花性状对陆地棉杂种优势的影响[J]. 山东农业科学,2011(3):11-14.
[33]赵军胜,王秀丽,高明伟,等. 红花性状标记杂交棉新品种鲁05H9 SSR指纹图谱构建及应用[J]. 山东大学学报(理学版),2014,49(1):44-49.
[34]王家宝,赵军胜,高明伟,等. 红花标记杂交棉鲁8H29的选育及配套栽培技术[J]. 中国棉花,2015,42(12):33-34.
[35]钱玉源,张海娜,崔淑芳,等. 棉花色素腺体的研究进展[J]. 作物研究,2017,31(2):205-210.
[36]袁有禄,靖深蓉,张天真,等. 陆地棉显性无腺体指示性状的杂种优势利用研究[J]. 种子,1997(6):15-17.
[37]权月伟,李继军,唐光雷,等. 河北省低酚棉育种研究进展及应用前景[C]//2015全国棉花青年学术研讨会论文集. 新疆石河子:中国棉花杂志社,2015.
[38]喻树迅,原日红,余学科,等. 低酚棉中棉所20遗传特异性与丰产性机理的研究[J]. 中国农业科学,1999,32(5):16-22.
[39]祝水金,蒋玉蓉,Reddy N,等. 陆地棉子叶色素腺体延缓形成种质系的育成及其遗传研究[J]. 科学通报,2004,49(19):1987-1992.
[40]陈 莹,赵军胜,高明伟,等. 腺体标记棉花杂交种鲁棉研39号指纹图谱构建[C]//中国棉花学会2014年年会论文集. 呼和浩特:中国棉花杂志社,2014.
[41]翟雷霞,米换房,李继军,等. 抗虫低酚棉品种邯无216的选育及栽培技术要点[J]. 中国棉花,2016,43(7):33-34.
[42]刘 芳,谭忠宁,比买热木·阿不都艾海提. 47个低酚棉品种资源性状观察与分析[J]. 中国棉花,2015,42(7):15-17.
[43]李函利,王省芬,王国宁,等. 低酚棉种质资源耐盐性鉴定[J]. 植物遗传资源学报,2016,17(6):1099-1105.
[44]刘 慧,季灵艳,赵天伦,等. 陆地棉色素腺体与不同棉酚旋光体含量之间的相关性研究[J]. 棉花学报,2017,29(5):437-446.
[45]张金发,孙济中,刘金兰. 棉花窄卷苞叶基因对产量品质的影响[J]. 作物学报,1993,19(4):315-320.
[46]张金发,孙济中,刘金兰. 窄卷苞叶基因对棉花光合性能的影响[J]. 华中农业大学学报,1993,12(4):328-332.
[47]张金发,孙济中,刘金兰. 窄卷苞叶基因对棉花早熟性的影响[J]. 华中农业大学学报,1991,10(4):321-326.
[48]李俊兰,崔淑芳,韩择林. 棉花新类型——观赏棉花[J]. 中国棉花,2003,30(9):29-51.
[49]狄佳春,殷剑美,陈旭升,等. 陆地棉亚红株突变体光合特性初步研究[J]. 江蘇农业科学,2006(2):33-34.
[50]万艳霞,马峙英,王国印,等. 红叶棉花的产量杂种优势研究初探[J]. 棉花学报,2010,22(2):186-188.
[51]陈旭升,狄佳春,赵 亮. 具有亚红叶标记的抗虫彩色杂交棉选育[J]. 江西农业学报,2015,27(4):1-3.
[52]刘英新,王学德,倪 密,等. 四个带有标记性状的棉花CMS恢复系的主要农艺与光合生理特性[J]. 棉花学报,2010,22(5):448-453.
